3个绒山羊品种DRB1基因遗传变异和生物信息学分析
2022-12-15吕雪峰许艳丽郑文新
吕雪峰, 许艳丽, 郑文新, 胡 昕, 王 乐
(新疆畜牧科学院 畜牧业质量标准研究所, 新疆 乌鲁木齐 830000)
0 引言
【研究意义】绒山羊是我国重要的牲畜品种之一,为人类提供羊绒、羊皮和羊肉等重要畜产品,在畜牧业发展中占有重要地位,特别在西北荒漠地区,绒山羊一直是牧民经济收入的重要来源。我国绒山羊经过长期的自然选择和人工选育,已经拥有了许多产绒量高、绒品质优的品种,如辽宁绒山羊、内蒙古绒山羊、新疆山羊、西藏山羊和陕北白绒山羊等,据2021年《中国统计年鉴》统计,截止2020年底,我国山羊存栏13 345万只,山羊绒产量15 244 t,是绒山羊养殖和产绒大国,在世界上占有举足轻重的地位。近年来,随着生态环境保护意识增强,绒山羊养殖已经逐步转向半舍饲或全舍饲,为追求更高的经济效益,高繁殖力和抗病性成为选育关注的焦点。主要组织相容性复合体(Major histocompability complex, MHC)是由紧密连锁、高度多态的基因座组成的复合体,具有高度的多态性,控制着机体对抗原产生免疫应答的能力[1]。由I类、II类和III类基因组成,其中II类基因的多态性在动物免疫中发挥最主要的作用,II类基因包含DQ、DR、DO等亚区,其产物参与抗原递呈和T细胞激活,在机体免疫中有重要作用,是开展抗病育种的首选标记基因,同时适用群体适应性遗传变异研究[2-3]。因此,研究绒山羊DRB1基因有助于绒山羊抗病育种,还可为种质资源的深度挖掘、分子评价、优良绒山羊品种的保护和利用提供理论依据。【前人研究进展】MHC基因(Goat lymphocyte antigen,GOLA)遗传多态性的研究主要集中在DR亚区的DRB基因[4-6]和DQ亚区的DQA1基因[7-9],已经发现,DRB基因外显子多态性与绵羊布鲁氏菌病[10-11]、线虫[12]、包虫病[13]、葡萄球菌病[14]等多种疾病存在一定的关联性。在陕北绒山羊中发现DRB1*16 等位基因频率与球虫感染强度显著相关[15],河西绒山羊中发现DRB1*8可能是流产发病的遗传易感基因[16];黄兰等[17]在黔北麻羊DRB1基因Exon3 中检测7个SNPs,均引起RNA二级结构的改变,但并未引起抗原表位改变。【研究切入点】大量研究证实,DRB1基因作为抗病分子遗传标记,其多态位点主要位于第2外显子序列,新疆山羊、内蒙古绒山羊和西藏山羊生活的自然环境差异很大,在适应性方面有各自的特点,如西藏山羊受到高海拔适应性的选择[18],内蒙古绒山羊在人为选育方面强度更大。通过DRB1基因第2外显子研究我国3个主要绒山羊品种的群体结构、遗传多样性和适应性选择等方面还未见报道。【拟解决的关键问题】以新疆山羊、内蒙古绒山羊和西藏山羊为研究对象,通过分析3个绒山羊品种DRB1基因第2外显子序列的遗传变异,以阐明该基因在不同绒山羊品种中的遗传多样性和遗传变异,并利用生物信息学软件对编码序列进行分析和功能预测,筛选出绒山羊疾病抗性和易感病候选基因,以期为开展抗病育种提供科学依据。
1 材料与方法
1.1 材料
新疆山羊、内蒙古绒山羊和西藏山羊样品采集地点分别位于新疆和丰县、内蒙古鄂尔多斯市和西藏日土县,采集数量分别为33只、17只和22只。颈静脉采血,EDTA抗凝,冷藏保存。
1.2 方法
1.2.1 血液基因组DNA提取 采用生工生物工程(上海)股份有限公司的EZUP柱式动物基因组DNA抽提试剂盒提取。
1.2.2 引物设计与PCR扩增 采用在线primer设计DRB1基因第2外显子引物:5′-TATCCCGTCTCTGCAGCACATTTC-3′和5′-TCGCCGCTGCACACTGAAACTCTC-3′,预计扩增片段长度258 bp,引物由工生物工程(上海)股份有限公司合成。PCR扩增采用德国biometra PCR仪,反应体系(25 μL):基因组DNA 1 μL,10 μmol/L上、下游引物各1 μL,PCR Mix 12.5 μL,ddH2O 9.5 μL。PCR扩增程序:94℃变性3 min;94℃变性30 s,60℃退火30 s,72℃延伸30 s,30个循环;72℃延伸5 min,4℃保存。
1.2.3 测序及数据分析 PCR产物经1%琼脂糖凝胶电泳检测后,送工生物工程(上海)股份有限公司纯化和测序,测序结果在NCBI网站中比对,确保所得序列是目标序列,用Clustal X 1.81软件进行序列比对,采用DnaSP V5(version 5.10.01)计算各碱基组成比例、单倍型多样度(Hd)、核苷酸多样性(Pi)、平均核苷酸差异数(k)、品种间核苷酸平均差异数(Kxy)、基因分化系数(Gst)、核苷酸歧义度(Dxy)、遗传分化指数(Fst)和基因交流值(Nm),并进行中性检验;运用MEGA 7.0计算品种之间的遗传距离(采用Kimura双参数模型),计算序列同义突变率(dS)和非同义突变率(dN),分析DRB1基因受到的选择压力;采用NJ法(Bootstrap抽样重复次数为1 000次)构建系统发生树;用Arlequin V 3.5.1.3进行分子变异分析。
1.2.4 生物信息学分析 测定核酸序列翻译成氨基酸后,采用ProtParam工具(http://web.expasy.org/protparam/)分析蛋白的理化性质;Protscale工具(https://web.expasy.org/protscale/)分析蛋白质疏水性;SignalP 5.0 Server 工具(http://www.cbs.dtu.dk/services/SignalP/)预测信号肽;利用NetPhos 3.1 Server(http://www.cbs.dtu.dk/services/NetPhos/)分析磷酸化位点;利用SOPMA工具(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)分析二级结构;利用Phyre2(Esypred 3D Web Server 4.0)对DRB1蛋白三维结构进行预测(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)。
2 结果与分析
2.1 DRB1基因第2外显子序列特征
3个绒山羊品种的基因组 DNA 进行PCR扩增产物经电泳检测,得到与预期目标片段大小一致、整齐而清晰的条带,测序后与GeneBank(登录号:AB008346.1)中山羊CDS区比对,序列为DRB1基因第2外显子,经比对校正后得到240 bp的核酸序列。3个品种69条序列检出核苷酸多态位点32个,其中单碱基突变位点3个,简约信息位点29个,占全部核苷酸序列的13.33%。由表1可见,基因序列中T、C、A和G含量分别为16.9%、24.2%、22.2%和36.7%,其中A+T含量(39.1%)远低于C+G含量(60.9%)。
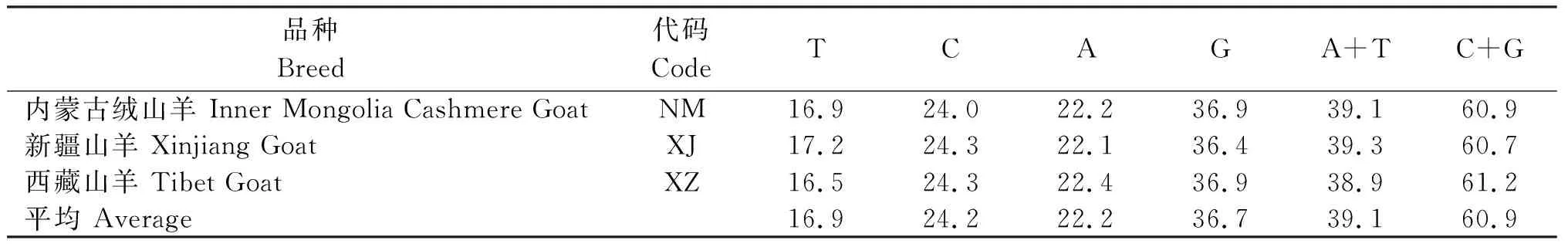
表1 3个绒山羊品种DRB1基因第2外显子序列碱基组成
2.2 3个绒山羊品种的遗传多样性
由表2可知,3个绒山羊品种群体的单倍型多样度(Hd)均大于0.9,其中,新疆山羊的单倍型多样度最高,为0.993;西藏山羊最低,为0.900。3个绒山羊品种的平均核苷酸多样度均大于0.02,其中,西藏山羊的核苷酸多样度最高,为0.060 25;内蒙古绒山羊最低,为0.022 00。3个绒山羊品种Tajima’s D中性检测无显著性差异。
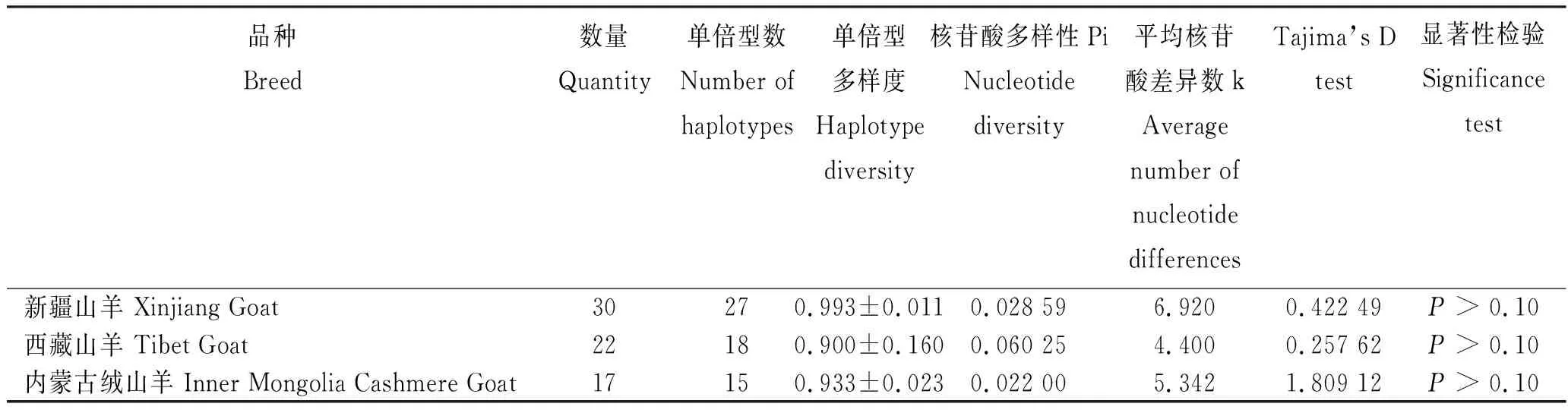
表2 3个绒山羊品种的采集信息与遗传多样性
2.3 3个绒山羊品种的遗传分化
经计算3个绒山羊品种基因差异程度的遗传学参数显示,3个绒山羊品种总基因分化系数(Gst)为0.002 73,总遗传分化指数(Fst)为0.181 24,总基因流(Nm)为1.13。3个绒山羊品种间基因分化系数(Gst)为0.001 03~0.003 98,均值为0.002 42;遗传分化指(Fst)为0.116 14~0.212 63,均值为0.177 65;核苷酸差异数(Kxy)为12.106 95~15.243 94,均值为13.772 52;核苷酸歧义度(Dxy)为0.050 24~0.063 25,均值为0.057 15。
2.4 3个绒山羊品种的遗传距离
从NCBI中下载西班牙野山羊(登录号:AF461696)、野山羊(登录号:U00198)、苏格兰绒山羊(登录号:LS974820)、印度罗伊尔坎迪山羊(登录号:KT624235)、洛基山山羊(登录号:DQ648493)和萨能奶山羊(登录号:AB008362)的同源序列,以Kimura-2参数为模型,计算各品种之间的遗传距离(表3)。3个绒山羊品种之间的遗传距离比较接近,内蒙古绒山羊与新疆山羊之间的遗传距离为0.070,与西藏山羊之间的为0.084,新疆山羊与西藏山羊之间的为0.098。西藏山羊与西班牙野山羊之间的遗传距离较小,仅0.048,与野山羊之间的为0.075;新疆山羊和内蒙古绒山羊与西班牙野山羊间的遗传距离相同,均为0.084。3个绒山羊与其他山羊品种之间的遗传距离较远,为0.080~0.128。
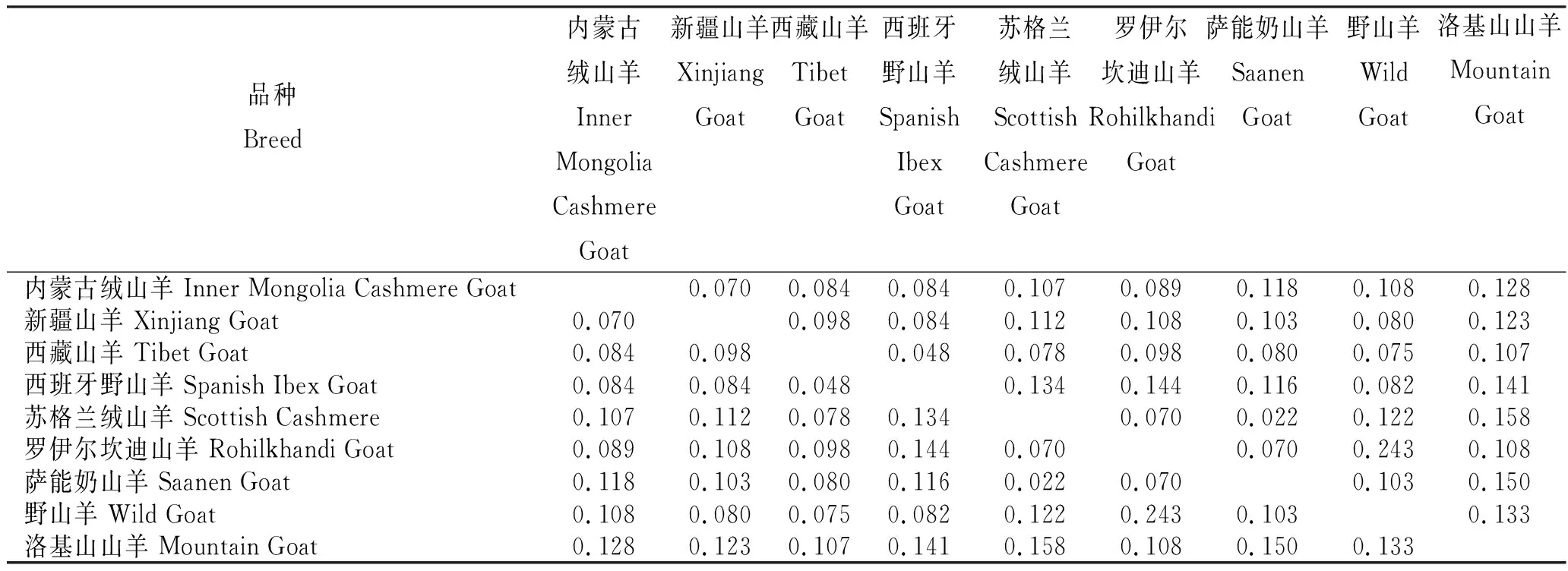
表3 9个山羊品种间的遗传距离(下三角)和标准误(上三角)
2.5 3个绒山羊品种的系统发育关系
以邻接法Neighbour-Joining构建3个绒山羊品种的系统发育树(图1a),新疆山羊先与内蒙古绒山羊聚在一起,然后再与西藏山羊聚在一起。以NCBI数据库中下载的6个山羊品种为外群,与3个绒山羊品种构建系统进化树(图1b)显示,西藏山羊先与西班牙野山羊聚在一起,再与野山羊聚在一起,最后与新疆山羊和内蒙古绒山羊聚为一个大的分枝,其余4个国外山羊品种(苏格兰绒山羊、印度罗伊尔坎迪山羊、洛基山山羊和萨能奶山羊)聚为另一分枝。
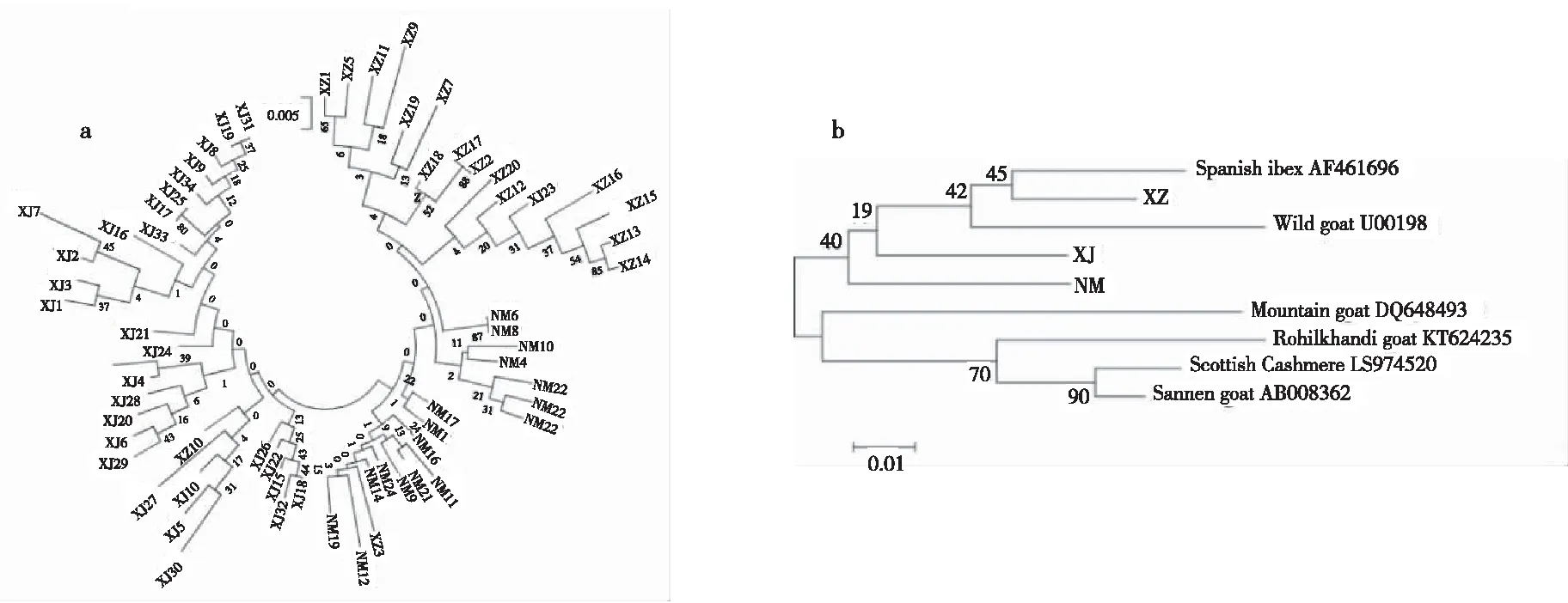
注:a,3绒山羊品种的系统发育树;b,9个山羊品种的系统发育树。
2.6 3个绒山羊品种的群体变异
经AMOVA 分子变异分析得出,3个绒山羊品种群体间的遗传变异为24.71%,远小于群体内的遗传变异(75.29%)。说明,遗传变异主要来自品种内,品种间的遗传变异较低。
2.7 3个绒山羊品种的选择压力
由表4可知,3个绒山羊品种DRB1基因受到明显的选择压力,序列中非同义替换率(dn)均显著大于同义替换率(ds)。其中,内蒙古绒山羊DRB1基因受到更强的选择压力,其次是新疆山羊,西藏山羊受到的选择压力最小。

表4 3个绒山羊品种DRB1基因同义替换与非同义替换率
2.8 DRB1序列生物信息学
2.8.1 蛋白理化性质 通过测序比对分析,绒山羊DRB基因第2外显子240个碱基共编码80个氨基酸残基,与GeneBank中AB008346.1中山羊完整CDS区比对,氨基酸差异达21个。选择每个品种中单倍型数量多的序列用于生物信息学分析,内蒙古绒山羊DRB1序列分子式为C416H620N128O134S2,预测分子量约为9 622.40 Da,理论等电点为5.15,由19种基本氨基酸组成,其中,精氨酸(Arg)含量最高,占氨基酸总数的15.0%,谷氨酸(Glu)其次,占12.5%(表5);含有带负电荷的残基(Asp+Glu)16个,带正电荷的残基(Arg+Lys)13个,280 nm 的摩尔消光系数为20 065 mol/cm,蛋白的平均亲水系数为-1.210,不稳定指数44.64,显示该蛋白不稳定。新疆山羊DRB1序列分子式为C426H626N124O129S2,预测分子量约为9 612.53 Da,理论等电点为6.13,由19种基本氨基酸组成,其中,精氨酸(Arg)含量最高,占氨基酸总数的12.5%,谷氨酸(Glu)其次,占10.0%;含有带负电荷的残基(Asp+Glu)13个,带正电荷的残基(Arg+Lys)12个,280 nm 的摩尔消光系数为21 555 mol/cm,蛋白的平均亲水系数为-0.945,不稳定指数50.74,显示该蛋白不稳定。西藏山羊DRB1序列分子式为C423H628N128O129S2,预测分子量约为9 634.54 Da,理论等电点为5.56,由19种基本氨基酸组成,其中,精氨酸(Arg)含量最高,占氨基酸总数的13.8%,谷氨酸(Glu)其次,占11.2%;含有带负电荷的残基(Asp+Glu)15个,带正电荷的残基(Arg+Lys)12个,280 nm 的摩尔消光系数为18 575 mol/cm,蛋白的平均亲水系数为-0.954,不稳定指数40.90,显示该蛋白不稳定。

表5 3个绒山羊品种DRB1基因序列的氨基酸组成
2.8.2 疏水性 利用ProtScale 对3个绒山羊品种DRB1氨基酸位点进行疏水性/亲水性预测(图2)。坐标轴X轴正值,说明此区域具有强疏水性;负值,说明此区域为亲水性。可见,3个绒山羊品种DRB1编码的蛋白总体表现出强的亲水性。内蒙古绒山羊DRB1序列第73位谷氨酸(Glu)疏水性最强(0.456),第25位精氨酸(Arg)亲水性最强(-2.211),整个蛋白序列中,亲水氨基酸占97.5%,疏水氨基酸占2.5%。新疆山羊DRB1序列第73位谷氨酸(Glu)疏水性最强(1.000),第48位天冬酰胺(Asn)亲水性最强(-2.167),整个蛋白序列中,亲水氨基酸占93.75%,疏水氨基酸占6.25%。西藏山羊DRB1序列第73位谷氨酸(Glu)疏水性最强(1.000),第47位色氨酸(Ser)和第48位天冬酰胺(Asn)亲水性最强(-2.333),整个蛋白序列中,亲水氨基酸占92.5%,疏水氨基酸占7.5%。
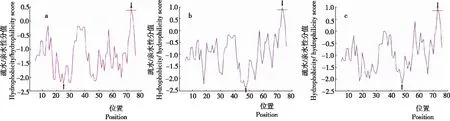
注:a为内蒙古绒山羊,b为新疆山羊,c为西藏山羊。箭头所指为疏水性/亲水性最强的氨基酸。
2.8.3 信号肽 3个绒山羊品种DRB1氨基酸序列进行信号肽预测显示,该蛋白不存在信号肽序列。
2.8.4 磷酸化位点 由图3可见,内蒙古绒山羊DRB1氨基酸序列共有4个酪氨酸磷酸化位点,3个苏氨酸磷酸化位点,4个丝氨酸磷酸化位点。新疆山羊DRB1氨基酸序列共有4个酪氨酸磷酸化位点,2个丝氨酸磷酸化位点,3个苏氨酸磷酸化位点。西藏山羊DRB1氨基酸序列共有3个酪氨酸磷酸化位点,3个苏氨酸磷酸化位点,2个丝氨酸磷酸化位点(得分均>0.5)。3个绒山羊品种DRB1氨基酸序列上磷酸化位点占比约10%,其中内蒙古绒山羊最多,占13.75%。
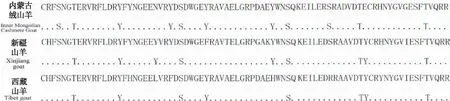
注:T为苏氨酸,S为丝氨酸,Y为酪氨酸。
2.8.5 二级结构和三级结构 3个绒山羊品种DRB1蛋白序列二级结构由α-螺旋、β-转角、延伸链和无规卷曲组成(表6),其中,主要以α-螺旋为主,其次是无规卷曲和延伸主链。利用Phyre2对3个绒山羊的DRB1蛋白进行三维结构预测,发现3个绒山羊品种DRB1蛋白三维结构基本相同(图4),结构中有较多的α-螺旋和无规卷曲,与二级结构预测相吻合。
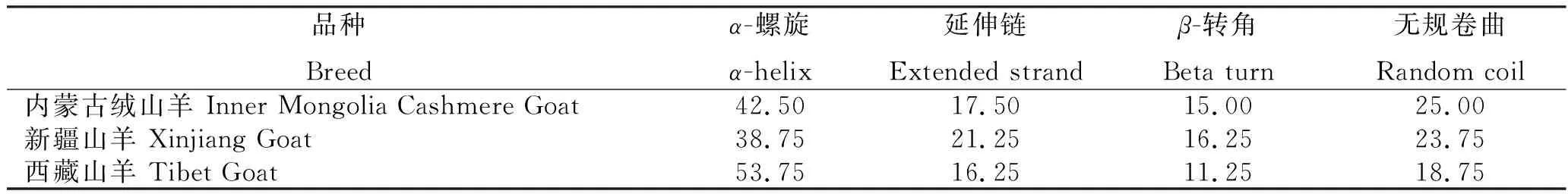
表6 3个绒山羊品种DRB1的二级结构
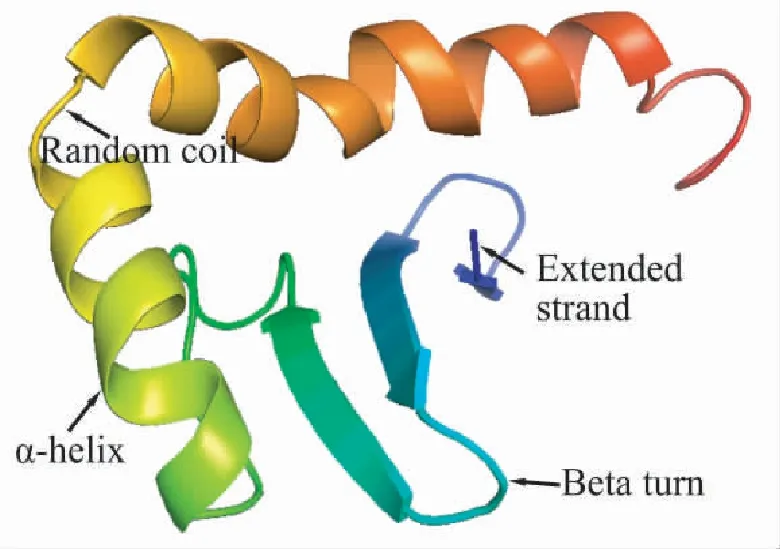
图4 绒山羊DRB1的三级结构
3 讨论
主要组织相容性复合体(major histocompatibility complex, MHC)是脊椎动物中与免疫相关的多基因家族,其蛋白产物负责识别和处理体内存在的外来致病因子[19]。MHC由Ⅰ、Ⅱ和Ⅲ类基因组成,山羊MHC II类抗原可与细胞外经加工后的抗原多肽结合,并将其提呈给表位特异性CD4+T淋巴细胞[20],从而引起机体的免疫调节反应[21]。MHC是脊椎动物免疫系统的关键组成部分,肽结合区决定了MHC蛋白可以结合的外源肽片段种类,这些区域的多样性增加MHC不同等位基因的基因产物和与不同抗原结合的可能性[22],从而赋予动物对某些抗原更大的易感性或抗性。已有研究发现,DRB基因在其他物种中均具有较高的多态性[23-24],本研究发现,新疆山羊、内蒙古绒山羊和西藏山羊的DRB1序列具有非常高的遗传多样性,单倍型多样度(Hd)均大于0.9,平均核苷酸多样性大于0.02,根据GRANT等[25]的分类,3个绒山羊品种均具有高的单倍型多样度(Hd>0.5)和核苷酸多样性(Pi>0.005);从核苷酸序列和氨基酸序列看,3个绒山羊品种检出核苷酸多态位点 32个,氨基酸差异达21个,分别占总数的13.3%和26.25%,与陕北白绒山羊的研究相似[26],说明这3个绒山羊品种在进化中产生了诸多突变,积累了丰富的单倍型和核苷酸变异,该物种对于变化的环境具有较强的生存能力。研究发现,3个绒山羊品种DRB1基因均受到较强的选择压力,其中,内蒙古绒山羊和新疆山羊受到的选择压力更强,可能是由于长期选育造成,2个品种是西北地区最主要的绒山羊品种,相对于西藏山羊来说,在生产性能的选育提高上力度更大,这种选择压力可能有助于适应当地寒冷、风沙等气候环境。
遗传分化指数Fst值为0~0.05,表明群体间的遗传分化程度很小;为0.05~0.15,表明群体间遗传分化程度中等;为0.15~0.25,表明群体间的遗传分化较大[27]。3个绒山羊品种总遗传分化指数Fst为 0.181 24,大于0.15,表明3个品种间的遗传分化较大,分子变异分析也表明,遗传变异主要来自于品种间而非品种内,与品种间遗传分化程度较高的结论一致。张乐超等[28]对我国7个地方山羊品种遗传结构研究也发现,我国的山羊群体遗传多样性丰富,群体间遗传分化程度大,基因交流少。
遗传距离和系统进化树分析显示,新疆山羊先与内蒙古绒山羊聚在一起,然后再与西藏山羊聚在一起,西藏山羊与其他2个品种遗传距离相对较远。狄冉等[29]通过微卫星将中国10个绒山羊品种进行了分类,其中一类由辽宁绒山羊、新疆山羊、柴达木山羊及陕北山羊组成,另一类由内蒙古绒山羊组成,与本研究基本一致。3个绒山羊品种与野山羊的亲缘关系更近,而与其他4种国外山羊品种的亲缘关系较远,表明我国的绒山羊可能是从野山羊驯化、选育而来。关于山羊起源研究的说法较多,LUIKART等[30]认为山羊主要存在3个起源。已有报道,西藏山羊部分支系起源于镰刀型角野山羊(Capraaegagrus)和捻角山羊(Caprafalconeri)[31],本研究也证实了这一点。我国山羊绒品质优良,市场需求大,我国也是山羊绒的出口国,羊绒及制品主要出口到欧洲、美国、日本及东南亚国家。因此,在绒山羊遗传资源上未与国外绒山羊进行交流,具有明显的地域性遗传特征,与国外山羊在遗传和起源上有明显的区别。据MANJUNATH等[32]对印度10个山羊品种mtDNA检测发现,印度家养山羊属于同一单系群,与所有野生山羊序列不同,因此,印度山羊可能来自一个未知的种群。
生物信息学分析显示,绒山羊DRB1基因第2外显子编码蛋白可判断为酸性蛋白,具有强的亲水性,无信号肽序列,有大量的磷酸化位点,可被不同的催化激酶引发磷酸化,这种磷酸化修饰可能使DRB1的构象更稳定,使其能很好地发挥生物学功能。二级结构预测结果显示,绒山羊DRB1基因第2外显子主要由α-螺旋和无规卷曲组成。在蛋白的二级结构中,起稳定蛋白作用的二级结构为α-螺旋和β-折叠,作为蛋白的骨架,不易变形,支撑着蛋白质的结构,抗体难以嵌合,而无规卷曲结构松散,则决定着蛋白质的功能,很可能成为抗原表位[33-34]。抗原通过与淋巴细胞的抗原受体结合,从而激活淋巴细胞,产生免疫应答。DRB1基因二级结构中无规卷曲比例也比较大,说明抗原结合位点多,对抗原捕获和呈递强,这些结合位点是由基因编码的且高度变化以调节结合位点的侧链[35],核苷酸序列的突变可能会改变编码氨基酸,最终导致结合位点构象变化,从而影响抗原的结合,引起动物对疾病的抵抗力或敏感性改变。
4 结论
新疆山羊、内蒙古绒山羊和西藏山羊3个绒山羊品种均具有丰富的遗传多样性,DRB1基因受到较强的选择压力,品种间遗传分化程度较高,3个绒山羊品种与野山羊的亲缘关系更近,表明3个绒山羊可能是从野山羊驯化、选育而来。绒山羊DRB1 基因第2外显子编码蛋白为酸性蛋白,具有强的亲水性,无信号肽序列,有大量的磷酸化位点,二级结构主要由α-螺旋和无规卷曲组成。
