涝渍胁迫对入侵植物白花鬼针草及其本地近缘种金盏银盘生长和生理特性的影响
2022-12-14岳茂峰尹爱国辛国荣
宋 玉,岳茂峰,尹爱国,辛国荣
1广东石油化工学院生物与食品工程学院/岭南现代农业科学与技术广东省实验室茂名分中心,广东 茂名 525000;2中山大学生命科学学院,广东 广州 510275
生物入侵、动植物栖息地丧失和全球气候变化并称为21世纪全球三大环境问题,三者常交织在一起对生态系统造成重大影响(Salaetal.,2000)。在全球气候变化下,极端气候将导致动植物栖息地环境因子发生变化,改变生态系统中各生物的生长发育和竞争效应,进而影响生物入侵的进程(Azzurroetal.,2019;Liuetal.,2021)。因此,极端气候造成的动植物栖息地生境改变如何影响生物入侵是当前生态学需要回答的科学问题之一。
极端降水是全球气候变化造成的极端气候之一。当前,极端降水事件在全球范围内的频率和强度均显著增加,并且这种状况可能继续恶化(Donatetal.,2016)。极端降水所造成的土壤水分变化影响稳定生态系统中不同植物的竞争关系,重塑新的竞争格局,进而影响外来植物的入侵能力(Drakeetal.,2016)。涝渍是极端降水引起的常见气象灾害,对陆生植被生长、发育造成严重危害(Le Provostetal.,2016;Niinemets &Valladares,2006)。虽然已有学者研究了涝渍如何影响旱地作物的生长(Mendiondoetal.,2016;Setter &Waters,2003),但关于涝渍对入侵植物影响的研究主要集中在湿地和河岸生态系统,且不同入侵植物对涝渍的响应并不一致(Allred &Cary,2019;Chenetal.,2013;Yueetal.,2019a)。涝渍对外来植物入侵能力的影响仍不明确,我国华南地区降水丰富,雨季降水常持续数日,使得旱地土壤水分频繁处于饱和状态,当前雨季持续性降水所造成的土壤涝渍在华南地区外来植物入侵过程中所扮演的角色仍不清楚(王家宜等,2017;Sunetal.,2020)。因此,明确入侵植物与本地植物对涝渍胁迫响应的差异,对揭示华南地区外来植物的入侵机制具有重要意义。
白花鬼针草Bidensalba(L.) DC,有别于传统“白花鬼针草”Bidenspilosavar.radiataSch.-Bip.,属于菊科鬼针草属,原产于热带美洲,是一种近年来在我国华南地区新暴发的一年生或多年生入侵植物(田兴山等,2010),在我国广东、广西、福建等地区作物田、果园、疏林和荒地等生境造成严重危害,已成为我国华南地区危害最严重的入侵植物之一(岳茂峰等,2016)。虽然部分学者从白花鬼针草的种子萌发、生长和繁殖特性等研究了白花鬼针草入侵机制(罗娅婷等,2019;韦春强等,2013;闫小红等,2019),但上述研究仍无法充分解释其在我国华南地区暴发成灾的机制。研究表明,白花鬼针草在我国的高度适生区主要分布在北回归线两侧附近(岳茂峰等,2016),华南地区位于北回归线附近,该地区极端降水引起的涝渍是否促进了白花鬼针草的入侵还不明确。
本研究以入侵植物白花鬼针草为例,其本地近缘种金盏银盘Bidensbiternata(Lour.) Merr.et Sherff为对照(金盏银盘与白花鬼针草在华南地区生境相似,但其分布范围更广)(Yueetal.,2017),通过研究二者在涝渍胁迫下的生长和生理特性变化,分析其生长和生理特性对涝渍胁迫响应的差异,明确涝渍对白花鬼针草入侵能力的影响,从而揭示白花鬼针草在华南地区的入侵机制,为白花鬼针草的防控和管理提供科学依据。
1 材料与方法
1.1 研究材料
入侵植物白花鬼针草及其本地近缘种金盏银盘的种子均于2015年秋季采自广东省广州市番禺区。
1.2 试验方法
试验于2016年3—4月在广东省农业科学院植物保护研究所进行。地理位置为N23°8′46″;E113°20′52.4″,海拔高度14 m,属于南亚热带气候,年平均温度21.4~21.9 ℃,年平均降雨量1623.6~1899.8 mm。先将白花鬼针草和金盏银盘种子放入盛有滤纸的培养皿在人工气候箱(30/25 ℃,12 h光照)中育苗。当幼苗生长到2叶期,将幼苗移栽到装有砂土(pH6.0、有机碳9.50 g·kg-1、有效氮59.63 mg·kg-1、有效磷40.5 mg·kg-1、有效钾44.5 mg·kg-1)的塑料盆(直径14 cm,高13 cm)中培养,每盆种植1株植物。当幼苗生长到6叶期(高度13 cm左右)开始进行涝渍胁迫试验。试验设置涝渍和对照2种处理。其中,涝渍处理是将种有植株的花盆浸在水里,土壤表面与水平面齐平;对照处理的土壤含水量维持在土壤最大持水量的70%~80%。每种处理40个重复,共计160盆。分别于处理第0、5、10、15、20天取出8株植株测定相关参数。涝渍胁迫前选取与试验处理幼苗一致的2种植物的幼苗各8株进行烘干处理,测定其初始生物量。试验于处理20 d后结束。
1.3 生长及生理指标观测
1.3.1 生长指标 试验处理期间,每5 d测定一次株高(cm),根、茎、叶生物量(g),叶面积(cm2),不定根数目,不定根长度(cm),计算总生物量、根冠比、比叶面积(specific leaf area,SLA,cm2·g-1)及相对生长速率(relative growth rate,RGR,mg·d-1·g-1)。
1.3.2 生理指标 (1)光合参数。在涝渍处理的第0、5、10、15、20天上午10:00左右用Li-6400便携式光合作用测定系统(LI-COR Inc.,Lincoln,NE,USA)测定叶片净光合速率(net photosynthetic rate,Pn,μmol·m-2·s-1)。所测叶片均使用饱和光强1200 μmol·m-2·s-1,CO2浓度保持在380 μmol·mol-1,叶面温度设定为25 ℃。所有处理过的样品在测量前暗适应20 min。光系统II(photosystemⅡ,PSⅡ)的能量分配利用叶绿素荧光仪(Dual-PAM-100,Heinz Walz,Effeltrich,Germany)通过在饱和脉冲下的慢动力诱导曲线测定(Klughammer &Schreiber,2008)。通过对慢动力曲线分析获得光系统Ⅱ最大光量子产量(the maximum quantum yield of PSⅡ,Fv/Fm)(Schreiberetal.,1986)、有效光量子产量[the effective quantum yield of PSⅡ,Y(Ⅱ)](Gentyetal.,1989)、非调节性能量耗散的量子产量[the quantum yield of non-regulated energy dissipation of PSⅡ,Y(NO)](Caillyetal.,1996)和调节性能量耗散的量子产量[the quantum yield of regulated energy dissipation of PSⅡ,Y(NPQ)](Krameretal.,2004)。
(2)根系脱氢酶活力。根系脱氢酶的活力(root dehydrogenase activity,RDA,mg·g-1·h-1)利用TTC(氯化三苯基四氮唑)法来测定(Liuetal.,2015)。涝渍处理前5 d时根系仍以旧根为主,所以原有根系被用于测定涝渍处理第0和5天后根系脱氢酶的活力。5 d后原有根系逐渐消失,因此,第10天开始新生不定根被用于测定根系脱氢酶的活力。
1.4 数据处理与分析
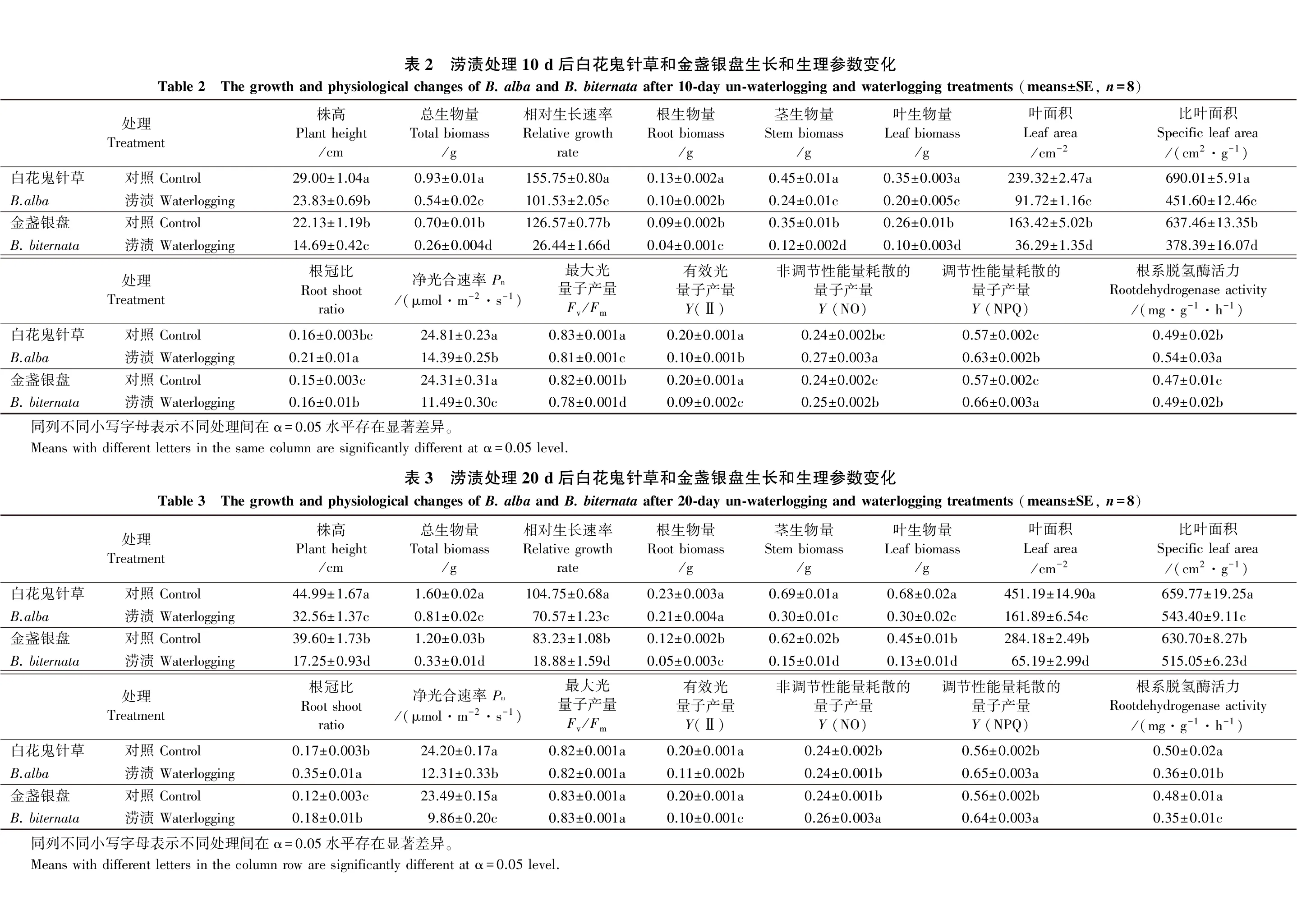
利用SPSS软件进行统计分析。采用二因素方差分析(two-way ANOVA)来评价涝渍处理10和20 d后物种、涝渍以及它们的交互作用对生长和生理的影响;各处理间生长和生理参数的差异使用单因素方差分析(one-way ANOVA:Tukey′s test)进行显著性检验。在涝渍胁迫下白花鬼针草和金盏银盘生长和生理参数差异利用独立样本T检验进行比较分析。采用SigmaPlot 14.0绘图。
2 结果
2.1 涝渍胁迫后白花鬼针草和金盏银盘生长参数的变化
在涝渍处理5、10、15和20 d后,白花鬼针草的株高、新生不定根数量和长度显著高于金盏银盘(图1A、B、C)。试验处理10 d后,涝渍对白花鬼针草和金盏银盘的株高、总生物量、RGR、根生物量、茎生物量、叶生物量、叶面积、SLA和根冠比产生显著影响,且物种间各参数差异显著(表1)。与对照相比,白花鬼针草和金盏银盘的株高、总生物量、RGR、根生物量、茎生物量、叶生物量、叶面积和SLA均显著降低,且物种间存在显著差异(表2),其中白花鬼针草的各参数分别下降17.83%、41.94%、34.81%、23.08%、46.67%、42.86%、61.67%和34.55%,金盏银盘的各参数分别下降33.62%、62.86%、79.11%、55.56%、65.71%、61.54%、77.79%和40.64%。与白花鬼针草相比,金盏银盘各参数下降更加明显。与此同时,白花鬼针草和金盏银盘的根冠比均分别显著增加31.25%和6.67%。
试验处理20 d后,处理和物种对生长参数均有显著影响(表1)。与对照相比,白花鬼针草和金盏银盘的株高、总生物量、RGR、根生物量、茎生物量、叶生物量、叶面积和SLA均显著降低(表3),其中白花鬼针草的各参数分别下降27.63%、49.38%、32.63%、8.70%、56.52%、55.88%、64.12%和17.64%,金盏银盘的各参数分别下降56.44%、72.50%、77.32%、58.33%、75.81%、71.11%、77.06%和18.34%。与白花鬼针草相比,本地种金盏银盘各参数(SLA除外)下降更加明显。白花鬼针草和金盏银盘的根冠比则分别显著增加105.88%和50.00%。
2.2 涝渍胁迫后白花鬼针草和金盏银盘生理参数的变化
在涝渍处理前期(5和10 d后),白花鬼针草的Pn、Fv/Fm、Y(NPQ)和RDA均显著高于金盏银盘(P<0.01),后期(20 d)2物种的Fv/Fm、Y(NPQ)和RDA趋于一致(图1D、E、H和I)。
试验处理10 d后,涝渍对白花鬼针草和金盏银盘的Pn、Fv/Fm、Y(Ⅱ)、Y(NO)、Y(NPQ)和RDA均具有显著影响(表1)。与对照相比,涝渍胁迫下白花鬼针草和金盏银盘的Pn、Fv/Fm、Y(II)均显著下降,白花鬼针草的各参数下降比例小于金盏银盘(表2),其中:白花鬼针草的各参数分别下降42.00%、2.41%和50.00%;金盏银盘的各参数分别下降52.74%、4.88%和55.00%。白花鬼针草和金盏银盘的Y(NO)、Y(NPQ)和RDA均有所上升,其中,白花鬼针草的各参数分别上升12.50%、10.53%和10.20%,而金盏银盘的各参数分别上升4.17%、15.79%和4.26%。
试验处理20 d后,涝渍对2物种的Pn、Y(Ⅱ)、Y(NO)和RDA均有显著影响,而对Fv/Fm和Y(NPQ)的影响不显著(表1)。与对照相比,涝渍胁迫下白花鬼针草和金盏银盘的Pn、Y(II)和RDA显著下降(表3),其中:白花鬼针草的各参数分别下降49.13%、45.00%和28.00%;金盏银盘的各参数分别下降58.02%、50.00%和27.08%。白花鬼针草和金盏银盘的Y(NO)分别上升16.07%和14.29%。二者的Fv/Fm和Y(NPQ)则无明显变化。
3 讨论
本研究结果表明,入侵植物白花鬼针草在短期涝渍胁迫下生物量和相对生长速率下降占比低于本地近缘种金盏银盘,表明涝渍对白花鬼针草的负面影响较小,涝渍有助于入侵植物白花鬼针草形成生长优势。该结果与陆生入侵植物三叶鬼针草B.pilosaL.的试验结果相似(Yueetal.,2019)。尽管二者分布范围有较大差异,但由于二者均起源美洲热带地区(田兴山等,2010),该地区雨水充沛,这可能使其形成了较强的涝渍适应能力。
涝渍对植物的影响作用于根系,快速形成不定根以代替因缺氧死亡的初生根,是耐淹植物应对涝渍胁迫的主要适应方式之一(Chenetal.,2016)。新生不定根可在伸长区形成发达的通气组织,使根内部组织孔隙度大幅提高,改善根系氧气摄取和运输能力,从而适应涝渍的低氧环境(Dominickj &Ninal,2005)。在本研究中,入侵植物白花鬼针草表现出更快的不定根生成和生长能力。这与以往耐水淹的植物常表现出快速不定根生成能力相一致(Vidozetal.,2016)。涝渍诱导形成的不定根也有利于水位回落后植物在土壤中的固定(Almeidaetal.,2003)。因此,涝渍胁迫下白花鬼针草不定根的快速生长不但有利于其快速的适应水淹低氧环境,而且有助于其在水位回落后恢复生长。植物在淹水的环境下可通过增加乙醇脱氢酶、乳酸脱氢酶等脱氢酶的活性启动无氧呼吸,以应对缺氧环境(Hawrylak-Nowaketal.,2015;Lietal.,2010)。本研究中入侵植物白花鬼针草的根系脱氢酶的活力在涝渍处理过程中始终高于本土植物金盏银盘。白花鬼针草更高的根系脱氢酶活力有利于其在缺氧状态下进行能量代谢,维持正常生长。可见,在涝渍胁迫下,入侵植物白花鬼针草不但可通过快速生成不定根来获得氧气,更强的脱氢酶活力也缓解了缺氧对根系造成的危害,有利于其在涝渍下生长发育。
光合作用是植物生长的决定性因素之一。本研究表明,在涝渍胁迫过程中白花鬼针草的Pn显著高于金盏银盘;与对照处理相比,白花鬼针草的Pn下降比例显著低于金盏银盘,这与白花鬼针草在胁迫过程中保持较高的生长速率相一致。植物生理代谢的变化与植物形态结构的改变密切相关(Gaseckaetal.,2012)。本试验中白花鬼针草能够保持更大的叶面积和SLA。SLA在一定程度上反映了叶片截获光的能力和在强光下的自我保护能力,能够反映植物在不同生境下资源获取能力(齐威等,2012)。涝渍条件下植物的叶片常常变小,以减少光的获得量来适应生理代谢(Fanetal.,2015)。本试验结果表明,白花鬼针草在涝渍下依然可通过更大的叶面积和SLA来获取等多的光能来维持生理代谢。此结果与涝渍胁迫下白花鬼针草和金盏银盘Pn的变化一致。可见,在涝渍胁迫下,白花鬼针草表现出更高的光合能力,这有利于白花鬼针草积累更多的碳水化合物来维持快速生长。
白花鬼针草和金盏银盘光能利用效率的改变也可以从其叶绿素荧光参数的变化反映出来。本研究中,白花鬼针草的Fv/Fm相对比较稳定,始终维持在0.80以上,而金盏银盘的Fv/Fm在涝渍处理后的5、10、15 d均低于0.80。一般来讲,植物在正常状态下的Fv/Fm为0.80~0.84。当植物的Fv/Fm小于0.8时,说明植物的光系统Ⅱ遭到显著破坏(Van Kooten &Snel,1990)。因此,在涝渍处理过程中白花鬼针草的光系统Ⅱ受到的破坏相对较轻,而金盏银盘的光系统Ⅱ受到明显破坏。
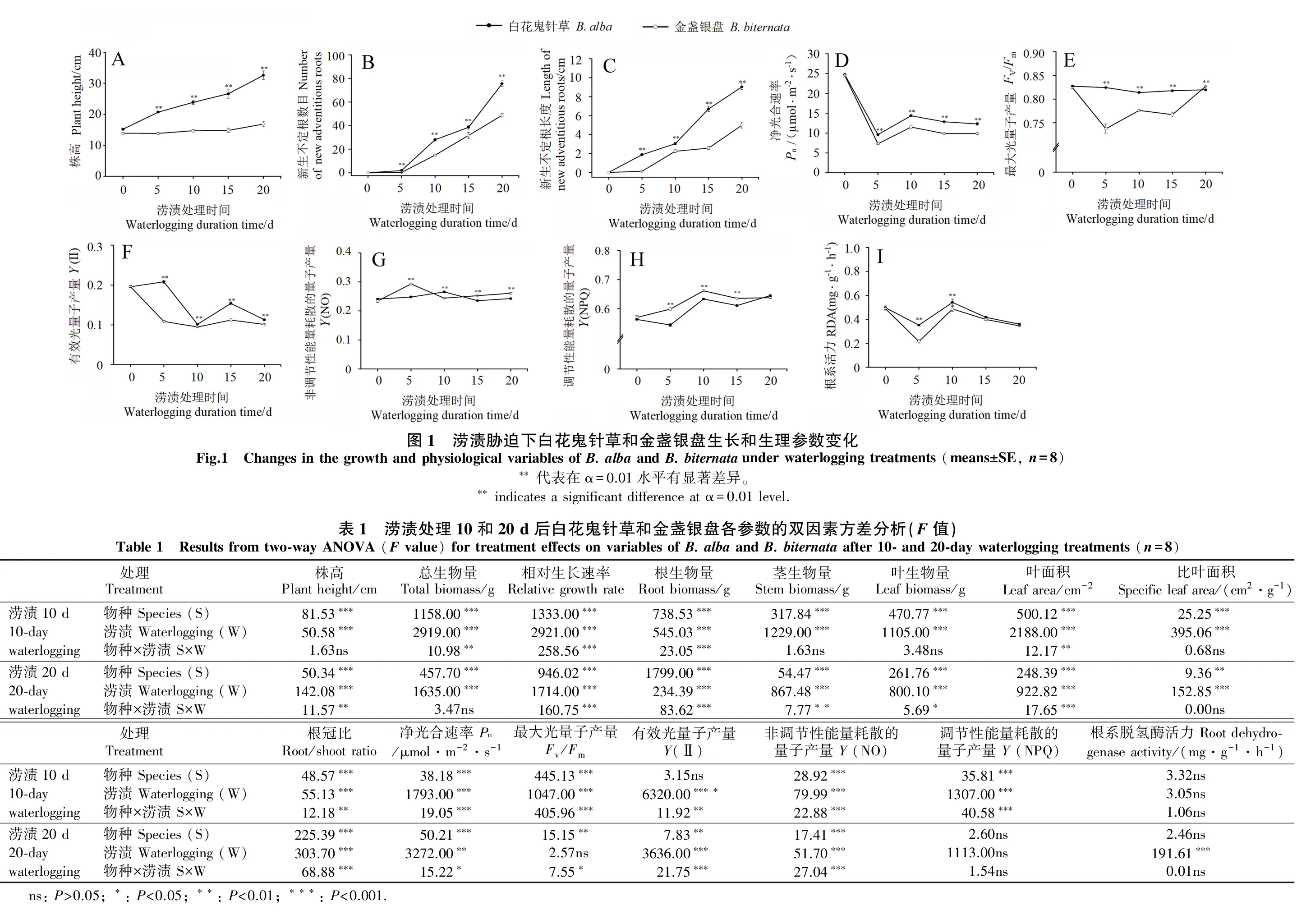
本试验中,除涝渍胁迫后10 d外,其他检测时间金盏银盘Y(NO)均显著高于白花鬼针草。Y(NO)是光损伤的重要指标,若Y(NO)较高,则表明光化学能量转换和保护性的调节机制不足以将植物吸收的光能完全消耗掉,植物可能已经受到损伤或者(尽管还未损伤)继续光照的话植物将受到损伤。可见,在涝渍胁迫过程中,本地种金盏银盘的光系统受到的破坏更加严重。与此同时,白花鬼针草Y(Ⅱ)在20 d内始终显著高于金盏银盘,说明白花鬼针草可把更加多的光能用于光合作用,以维持生长。
综上所述,白花鬼针草能够在涝渍条件下快速生成不定根缓解涝渍对其影响,并保持更高的净光合速率以维持快速生长,从而形成生长优势。白花鬼针草及其本地近缘种金盏银盘对涝渍的响应表明,我国南方地区雨季集中降水所造成的短期土壤涝渍有利于白花鬼针草的入侵。近期有研究表明,源于热带美洲地区的薇甘菊MikaniamicranthaH.B.K.表现出极强的耐水淹能力,但这种耐水淹能力在其入侵过程中发挥的作用尚不明确(Yueetal.,2019b)。全球气候变化下极端降水对源于美洲热带地区外来植物入侵能力的影响值得进一步关注。
