速生槐叶蘋入侵风险评估及管理措施分析
2022-12-14张国良袁至立王忠辉付卫东
张 岳,张国良,袁至立,宋 振,王忠辉,付卫东
中国农业科学院农业环境与可持续发展研究所,北京 100081
由外来植物引起的生物入侵已成为全球性的生态和经济问题(闫小玲等,2012)。入侵植物严重影响农业生产及其生态系统功能,并且破坏物种栖息地,影响生物多样性。建立外来植物入侵风险评估体系可对外来物种传入过程中产生的不确定事件进行识别、评估和处理,以最小的成本将各种不利后果减少到最低程度。完整的外来植物风险评估包括对外来入侵植物的各种特性、环境因素和人为因素进行分析,评价其传入、扩散的可能性,预测其对本地居民健康、经济活动、生态环境和社会生活等产生的影响,最后从社会、经济和政治等方面确认危害可以接受的程度,制定和实施降低风险措施(张瑞海等,2019;张国良等,2020)。
速生槐叶蘋SalviniamolestaD.S.Mitchell,隶属于槐叶蘋科Salviniaceae槐叶蘋属Salvinia,又称人厌槐叶蘋、蜈蚣蘋、山椒藻(万方浩等,2012;闫小玲等,2020),是一种原产于巴西东南部的多年生漂浮型水生植物(Forno,1983)。作为一种观赏和水族箱的造景植物,其被广泛引种至世界各地后逃逸至野外,侵入湖泊、河流和稻田等各种水生生境。基于对生物多样性和人类活动产生影响的严重性,以及能否阐明生物入侵的关键性问题等2个主要筛选标准(Loweetal.,2000),速生槐叶蘋被列为“世界100种恶性外来入侵物种”之一,是仅次于水葫芦Eichhorniacrassipes(Martius) Solms的世界第二大有毒水生植物(Luqueetal.,2013,2014)。
速生槐叶蘋在中国的分布范围有限,但其潜在的高入侵风险及危害不可忽视(闫小玲等,2020;冼晓清等,2022)。本文通过查阅梳理文献,对速生槐叶蘋开展定性和定量风险评估分析,建立风险评估体系,确定入侵我国的风险值,为其检疫决策及防治提供科学依据。
1 材料与方法
1.1 文献资料收集
速生槐叶蘋的地理分布数据来源于全球生物多样性信息网络(global biodiversity information facility,GBIF)(https:∥www.gbif.org/species/5274863)、国际应用生物科学中心(centre agriculture bioscience international,CABI)的入侵物种大全(invasive species compendium,ISC)(https:∥www.cabi.org/isc/datasheet/48447)和《中国外来入侵植物名录》(马金双和李慧茹,2018)。定性分析和定量分析的相关资料主要以“速生槐叶蘋”“人厌槐叶蘋”“Salviniamolesta”“kariba weed”为关键词检索中国知网和web of science数据库,并参考欧洲和地中海植物保护组织(European and Mediterranean Plant Protection Organization,EPPO)的EPPO全球数据库(EPPO global database)(https:∥gd.eppo.int/taxon/SAVMO)、《中国外来入侵植物志第一卷》(闫小玲等,2020)及《生物入侵:中国外来入侵植物图鉴》(万方浩等,2012)。
1.2 风险评估方法
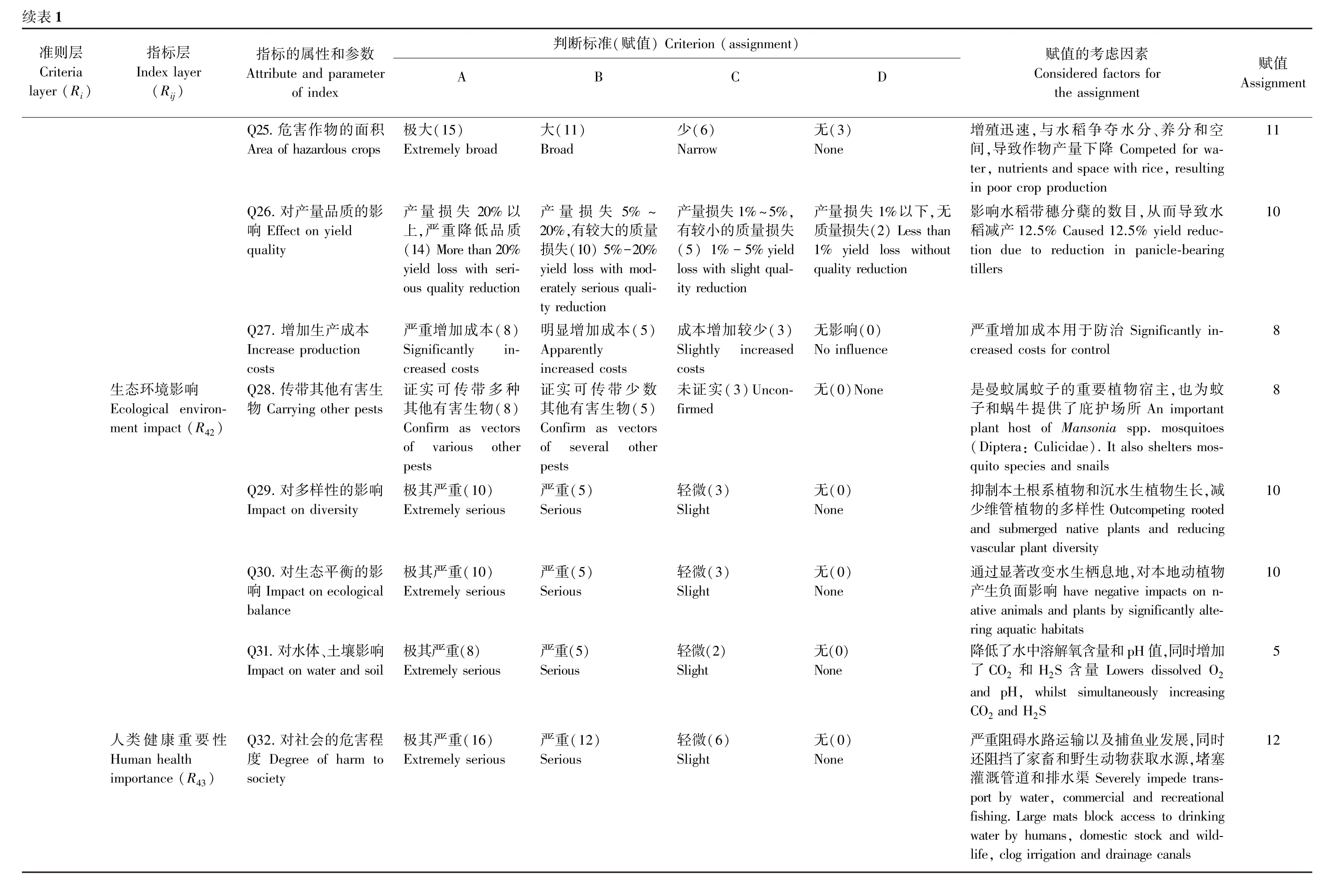
参照张国良等(2020)提出的外来植物风险指标体系和判断标准,分别对4个指标层(入侵性R1、适应性R2、扩散性R3和危害性R4)的17个指标和32个问题进行量化赋值,并计算出某植物的风险值R,即R=0.40R1+0.15R2+0.20R3+0.25R4
若R>61,则该植物应禁止引入;若29.5 2.1.1 起源、入侵扩散历史及分布情况 速生槐叶蘋原产于巴西亚热带地区(McFarlandetal.,2004)。1943年,斯里兰卡科伦坡大学植物系引入的速生槐叶蘋逃逸至野外,第一次在原产地之外建立野外种群(Oliver,1993)。在澳大利亚,于1952年首次发现逃逸,至1976年暴发,其危害程度超过了包括水葫芦在内的其他水生植物(Cronk &Fuller,2001)。1972年引入巴布亚新几内亚后,在塞皮克河泛滥。在非洲主要发生在湖泊-河岸系统,1962年在卡里巴湖其种群暴发,超过1/4的湖泊被其覆盖(McFarlandetal.,2004)。速生槐叶蘋于1995年在美国南卡罗来纳州的野外被监测到(Jacono &Pitman,2001)。随后于1998年入侵德克萨斯州和路易斯安那州,1999年入侵到佛罗里达州、阿拉巴马州、密西西比州、夏威夷州、亚利桑那州、加利福尼亚州和佐治亚州。 速生槐叶蘋目前已广泛分布于非洲(贝宁、博茨瓦纳、布基纳法索、喀麦隆、刚果、斯威士兰、肯尼亚、莱索托、马达加斯加、马拉维、马里、玛丽塔尼亚、毛里求斯、莫桑比克、纳米比亚、尼日利亚、塞内加尔、南非、坦桑尼亚、赞比亚、津巴布韦)、亚洲(印度、印度尼西亚、以色列、日本、马来西亚、巴基斯坦、菲律宾、新加坡、斯里兰卡、泰国)、欧洲(奥地利、比利时、法国、德国、以色列、意大利、荷兰、葡萄牙、西班牙、瑞士)、北美洲(墨西哥、美国、古巴、瓜德罗普、危地马拉、马提尼克、特立尼达和多巴哥)、南美洲(阿根廷、巴西、哥伦比亚、圭亚那)和大洋洲(澳大利亚、斐济、新西兰、巴布亚新几内亚)(EPPO,2022)。 在我国,速生槐叶蘋作为观赏植物最早由台湾引入,后逸生于野外。目前已分布于江苏、台湾、浙江、海南等省及香港特别行政区,也常见于各地花卉市场、植物园与水族馆中(马金双和李慧茹,2018)。 2.1.2 国外重要地位及管控措施 速生槐叶蘋于2007年被列入EPPO预警名单,在2019年被列入EPPO A2类检疫性有害生物名录(EPPO,2022)。澳大利亚将其列入国家有毒杂草清单,各州都出台了相应的管控措施(Australian Weeds Committee,2016)。2014年,南非将其列为1b类入侵物种,即“必须受到管控,并在可能的情况下铲除和销毁。严禁任何形式的贸易或种植”(http:∥www.environment.gov.za)。美国将其列入联邦有害杂草名单(未经许可在美国各州之间进口或运输该植物是非法的),被亚利桑那州、加利福尼亚州、科罗拉多州、佛罗里达州等州政府列为入侵物种(McFarlandetal.,2004)。 2.1.3 传播方式及传入途径 速生槐叶蘋可通过风、水流和动物活动(鸟类、水豚、河马和水牛等)进行自然传播(Forno &Smith,1999)。但主要通过人类有意引入(作为观赏植物或饲料)或无意引入(依附在船舶或掺杂在水生植物和鱼类的货物中)进行远距离传播(Gewertz,1983;Parsons &Cuthbertson,1992)。 2.1.4 检验鉴定难度 速生槐叶蘋隶属于美洲槐叶蘋复合体。该复合体包含4个物种,即速生槐叶蘋、耳形槐叶蘋S.auriculataAubl、S.bilobaRaddi和S.herzogiide la Sota (Mitchell,1972)。上述4个物种营养器官的形态特征极其近似,只能通过孢子果等生殖器官的细微差异进行识别。另外,速生槐叶蘋与槐叶蘋S.natans(Linnaeus) Allioni也极为近似,但前者浮水叶表皮毛顶端的分叉在末端闭合,后者的分叉不闭合,是区分二者的主要特征(Booyetal.,2015)。 2.1.5 防治难易程度 速生槐叶蘋的防治效率主要取决于入侵程度,当它处于低密度时,许多防治手段均可在短时间内有效根除(Koncki &Aronson,2015),因此规范引种行为、早日发现逃逸的植株并及时处理是控制其泛滥最有效的方法。当该种已大面积泛滥时,化学、物理防治效果甚微且耗资巨大,目前防治效果最佳的仍是以槐叶蘋象甲Cyrtobagussalviniae(Mart.) Solms为代表的生物防治。 化学防治:速生槐叶蘋叶片上表面密被笼状结构的毛,可形成大多数除草剂无法穿透的屏障。只能通过添加润湿剂或表面活性剂来增强除草剂的渗透作用(Oliver,1993)。目前已知有效的除草剂包括草甘膦(Nelsonetal.,2001)、2,4-二氯苯氧乙酸与十二烷基苯磺酸钙的混合制剂(Julian,1984)、百草枯、利谷隆、敌草隆等(Wellsetal.,1986)。 2.4 相关性分析 在观察组孕妇中,直线相关性分析显示,阴道微生态状况异常与早产、胎膜早破、绒毛膜羊膜炎、产褥感染、胎儿窘迫、新生儿感染呈现明显正相关性(r=0.432、0.384、0.552、0.384、0.392、0.362,P<0.05)。 物理防治:人工清除速生槐叶蘋可作为一种持续控制方法,但当其定殖且生物量达到8×106kg·km-2,速生槐叶蘋将进入快速生长发育阶段,机械收割和人工清除的成本将远超化学防治。 生物防治:在目前已知的所有天敌性昆虫中,仅有一种原产于巴拉圭、巴西和玻利维亚的半水生甲虫——槐叶蘋象甲被证明有效,并且成本低于化学或物理防治(Chikwenhere &Keswani,1997),已成功被引入16个国家进行生物防治(Julien &Griffiths,1998;Julienetal.,2002,2009),包括南非、博茨瓦纳、印度(Cilliers,1991;Creagh,1991)、巴布亚新几内亚(Thomas &Room,1986)、斯里兰卡(Room,1990)等,使速生槐叶蘋的发生面积锐减至原发生面积的1%~20%。但在澳大利亚新南威尔士州和北领地却引入失败,主要原因是槐叶蘋象甲对温度较敏感,水温过高或过低均不适合其生长发育(Oliver,1993)。此外,槐叶蘋象甲的摄食量与水体含氮量呈正相关,在低氮水平下,槐叶蘋象甲无法维持自身种群数量(Room,1990)。 2.2.1 气候适宜度 速生槐叶蘋的分布范围主要由海拔和最冷季的平均温度决定,全球气候变化将有利于其逐渐向温带地区传播定殖(Koncki &Aronson,2015)。由此推测,中国台湾以及华南的大部分地区都有遭受该种入侵的风险。 2.2.2 生存环境条件 速生槐叶蘋喜光照强、水温较高且营养丰富的水域(Mitchell &Tur,1975)。最佳生长发育pH值为6.0~7.5,水温为20~30 ℃(Cary &Weerts,1983)。在富营养化栖息地,如磷矿复垦湿地和池塘,特别适合其生长发育(Oliver,1993)。 2.2.3 分布生境特点 自然生境中,速生槐叶蘋喜静水或流动缓慢的水域,如湖泊、河流或溪流、湿地、池塘等(EPPO,2022)。易在稻田、运河、泄洪渠、人工湖以及水利发电设施等扰动生境中定殖(Forno &Harley,1979)。 2.2.4 耐逆性 速生槐叶蘋的厚垫状结构,可将其对霜冻的抵抗力提高到生理耐受性水平之上(Harley &Mitchell,1981),如Owensetal.(2004)发现其可在-3 ℃的试验池塘中存活48 h以上。而芽由于没有厚垫状结构的保护,暴露在低于-3 ℃或高于43 ℃的温度中超过2~3 h后即死亡(Whiteman &Room,1991)。速生槐叶蘋对盐度的耐受性较低,但可在盐度约为20%的低盐度海水中缓慢生长(Harley &Mitchell,1981)。 速生槐叶蘋是五倍体,因此减数分裂异常,产生的孢子败育,只能进行营养繁殖(Mitchell,1972),Barrett(1989)推测速生槐叶蘋是由槐叶蘋属其他物种杂交产生的,且入侵至全世界的所有速生槐叶蘋种群均是同一个遗传个体的克隆体。但目前尚未有相关的遗传学研究证明。 2.4.1 经济影响 速生槐叶蘋与水稻OryzasativaL.争夺水分、养分和空间,影响水稻带穗分蘖的数目,从而导致水稻减产12.5%(Azmi,1988),是印度、斐济、斯里兰卡等国家的主要稻田有害生物(Sundaresan &Reddy,1979;Thomas &Room,1986)。 2.4.2 生态环境影响 速生槐叶蘋可在水面形成1 m厚的垫状物,严重妨碍其他沉水植物的光合作用,从而抑制本土根系植物和沉水生植物生长,减少维管植物的多样性(Sculthorpe,1967)。垫状物可在较宽阔的水域中形成漂浮的小岛(Thomas,1981),显著改变水生栖息地,对本地动植物产生负面影响(Carpenter &Lodge,1986)。垫状物的形成还降低了水中溶解氧含量和pH值,同时增加了CO2和H2S含量,导致底栖动物数量减少(McFarlandetal.,2004;Mitchell,1969)。在印度,速生槐叶蘋已经侵入湿地并替代了本土的植物群落(Gopal,1988)。 2.4.3 对社会的影响 速生槐叶蘋是传播西尼罗病毒、圣路易型脑炎病毒、委内瑞拉马脑炎病毒和农村象皮病的曼蚊属蚊子的重要宿主植物(Chowetal.,1955;Lounibosetal.,1990;Pancho &Soerjani,1978;Ramachandran,1960),也为传播脑炎、疟疾和登革热的蚊子(Creagh,1991)和传播血吸虫病的蜗牛(Holmetal.,1977)提供了庇护场所。 速生槐叶蘋厚垫状结构使船只无法通过,严重阻碍水路运输以及捕鱼业发展。同时还堵塞灌溉管道和排水渠(Oliver,1993;McFarlandetal.,2004)。在巴布亚新几内亚的塞皮克河沿岸,速生槐叶蘋的入侵使当地居民的医疗卫生、教育和粮食安全等民生问题无法得到保障,被迫整体搬迁(Gewertz,1983)。 一些研究利用速生槐叶蘋生产沼气(Thomas &Room,1986),其能量生产潜力约为4.2×105J·km-2(Abbasietal.,1990)。速生槐叶蘋能够有效富集水体中的磷、氮、钾、锰、铁、锌等富营养化物质(Parsons &Cuthbertson,1992;Shimadaetal.,1988)及铬等重金属(Gupta &Devi,1995)。目前,速生槐叶蘋已经广泛应用于矿山生态修复(Sukumaran,2013)及污水处理(Mustafa &Hayder,2020,2021)。此外,速生槐叶蘋还是滞留空气层稳定性和减阻效应研究的仿生学模式材料(郑亚雯,2018;Barthlottetal.,2010)。 根据外来植物风险指标体系和判断标准,对速生槐叶蘋进行量化赋值(表1)。 根据表1 ,入侵性R1为72,适应性R2为53,扩散性R3为82,危害性R4为81。根据风险值计算公式R=0.40R1+0.15R2+0.20R3+0.25R4,计算出风险值R为73.4。因此,速生槐叶蘋应属于严格禁止引入的植物。 本文通过查阅梳理文献,对速生槐叶蘋开展定性和定量风险评估分析,建立风险评估体系,确定速生槐叶蘋入侵我国的风险值高。为防止其进一步扩散,建议相关管理部门采取如下措施加强管理。 (1)摸清速生槐叶蘋在我国的入侵现状。目前只知道速生槐叶蘋已经逸生到香港、江苏、台湾、浙江、海南等省市自治区,其具体的发生区县乡镇范围、发生面积、生境、危害对象及危害程度等信息仍处于空白状态,因此,应尽快通过问卷调查、实地踏查、样地调查等方式,摸清其在我国的入侵现状。 (2)预测速生槐叶蘋在我国的潜在适生区范围。考虑到速生槐叶蘋在我国仍有作为观赏植物和水族箱造景植物的市场需求,应尽快开展我国潜在适生区范围的预测研究。基于其在我国的潜在适生区范围,规范引种制度,划分允许销售区域和禁止销售区域,并且严禁与禁止销售区域开展跨区贸易往来。 (3)加强速生槐叶蘋的检疫、监测和监管力度。由于速生槐叶蘋是无性繁殖,逃逸至野外的一株个体也足以在其适生区快速定殖、生长分裂并造成危害。因此,加强对可能携带速生槐叶蘋的载体,如水生植物、热带鱼和水族箱等其他相关产品进行检疫工作,仔细检查是否有速生槐叶蘋掺杂其中。考虑到速生槐叶蘋较小且近缘种较多,难以通过形态特征进行有效鉴定,建议开发适用于现场检测的分子鉴定试剂盒。另外,应加强适宜生存区域开展重点监测工作,如湖泊、河流、湿地、稻田、灌溉渠道、沟渠、池塘和运河、人工湖以及水利发电设施等区域,研发集成生物芯片传感和环境DNA技术的高通量、高灵敏度、实时精准监测技术,及时发现逃逸的植株。 (4)研发速生槐叶蘋应急处置技术。在速生槐叶蘋处于低密度时,可通过多种防治手段在短时间内有效根除,对低密度、零星发生区采用物理防治进行人工摘除或机械清除;对成片发生的区域可采用物理防治与化学防治相结合的方法开展根除工作。目前仅有国外速生槐叶蘋的化学防治资料可供参考,考虑到不同国家的农药管理制度和允许使用的农药种类存在差异,建议在我国《农药管理条例》的框架下,使用已取得中国农药登记的农药组成成分,尽快研制适用于我国速生槐叶蘋化学防治的农药组合和有效剂量。 (5)建立速生槐叶蘋联防联控综合防治体系。在防控原则上,采取“预防为主,综合防治”的原则,在速生槐叶蘋的潜在发生区,加强风险评估、监测预警,强化植物检疫措施,防止其向未发生区传播扩散;在速生槐叶蘋的发生区以农业防治为基础,协调化学防治、物理防治、生物防治等措施,进行综合治理。目前,对于区域防治效果最佳的仍是以槐叶蘋象甲为代表的生物防治,但我国目前尚未开展相关研究,亟需因地制宜开展研究和评估。2 速生槐叶蘋入侵风险的定性分析
2.1 速生槐叶蘋的入侵性
2.2 速生槐叶蘋的适应性
2.3 速生槐叶蘋的扩散性
2.4 速生槐叶蘋的危害性
2.5 速生槐叶蘋利用价值
3 速生槐叶蘋入侵风险的定量分析



4 速生槐叶蘋管理措施建议
