氨基葡萄糖产生菌的筛选鉴定及其发酵优化
2022-12-13黄娟蔡俊
黄娟,蔡俊
(发酵工程教育部重点实验室,工业微生物湖北省重点实验室,工业发酵湖北省协同创新中心,湖北工业大学,湖北 武汉 430068)
氨基葡萄糖(glucosamine,GlcN)是由葡萄糖的一个羟基被氨基取代形成的一种重要的功能性单糖[1-4],其广泛存在于细菌、酵母、丝状真菌、植物和动物中[5]。作为生物体内多糖的重要单体之一,GlcN可有效治疗关节炎,参与肝肾解毒和护肝[6-8],还能抗衰老、刺激婴儿肠道内双歧杆菌的生长、调节内分泌[9]。
目前国内外生产GlcN主要有由虾蟹壳或者真菌菌丝体酸、酶解[10-14],工程菌发酵与真菌发酵这3种方法,但酸、酶解存在原料来源会引起过敏反应[15]以及酶成本高等问题。工程菌发酵产GlcN[16-19]已有大量研究,产量高且发酵周期短,在经济上占有优势,但工程菌有着表达系统不稳定,表达产物无活性、易被降解,食品市场难以接受等普遍性缺点。Hsieh等[5]比较发现野生菌株Aspergillus sp.BCRC 31742是产GlcN的最佳菌株,通过对该菌株发酵条件、菌丝体状态以及刺激因子等多方面提高其生物量[20-21]或者利用生产柠檬酸的废弃菌丝体[22]从而获得菌丝体中GlcN的方法,对于不接受用工程菌产GlcN的医药、食品市场而言具有可行性,但从丝状真菌菌丝体中提取GlcN仍存在纯度低、酶成本高以及发酵周期长等问题。Papagianni等[23]研究发现黑曲霉产柠檬酸发酵早期的发酵液中存在GlcN,在丝状真菌发酵初级阶段,细胞内大量合成的GlcN会分泌到胞外,该现象为GlcN的生产提供了新思路。因此本文提出从丝状真菌早期发酵液中提取获得GlcN。
本研究从养虾厂采集土壤样品进行筛选分离,得到一株胞外GlcN含量为1.537 g/L的菌株,进行形态学观察和分子学鉴定后确定为Aspergillus piperis,发酵12 h后发酵液中GlcN含量达到2.687 g/L,实现了GlcN胞外生产和积累。该策略解决了丝状真菌发酵周期长、GlcN提取工艺中存在环境污染以及工程菌危害的问题,为绿色便捷获得GlcN提供了理论依据。
1 材料与方法
1.1 材料与试剂
在湖北省潜江市小龙虾养殖池中采集土样。葡糖胺(分析纯)、对二甲氨基苯甲醛(分析纯):上海麦克林生化科技有限公司;D-氨基半乳糖盐酸盐(分析纯):上海源叶生物科技有限公司;10×扩增缓冲液(10×PCR buffer,含 Mg2+)、脱氧核糖核苷三磷酸(deoxy-ribonucleoside triphosphate,dNTP):美国赛默飞世尔科技公司(Thermo Fisher Scientific);引物 ITS1(10 μmol/L)、ITS4(10 μmol/L):南京金斯瑞生物科技股份有限公司;其他试剂均为国产分析纯。
1.2 培养基的配制
牛肉膏蛋白胨培养基[24]:牛肉膏5 g、蛋白胨10 g、NaCl 5 g、pH7.0。
高氏一号培养基[25]:可溶性淀粉20 g、KNO31 g、K2HPO40.5 g、MgSO4·7H2O 0.5 g、NaCl 0.5 g、FeSO4·7H2O 0.01 g。用少量冷水将淀粉调成糊状,倒入煮沸的水中,加入其他成分,溶化后补水至1 L。
马铃薯葡萄糖琼脂(potato dextrose agar,PDA)培养基[21]:土豆200 g、葡萄糖20 g。土豆去皮切块煮沸0.5 h,然后用纱布过滤,再加葡萄糖并定容至1 L。
水琼脂培养基平板[26]:琼脂20 g、水1 L。
查氏培养基[27]:NaNO32 g、K2HPO41 g、KCl 0.5 g、MgSO40.5 g、FeSO40.01 g、蔗糖 30 g。
1.3 仪器与设备
SYNERGY2酶标仪:基因生物技术国际贸易(上海)有限公司;Veriti实时荧光定量聚合酶链式反应(polymerase chain reaction,PCR)仪:美国赛默飞世尔科技公司;沃特世2767-2545液质联用仪(liquid chromatograph mass spectrometer,LC-MS):沃特世科技(上海)有限公司。
1.4 产GlcN菌株的分离筛选
1.4.1 菌株分离纯化
取1 g土样接入50 mL 0.9%NaCl溶液中并于摇床30℃,180 r/min培养2 h。将土样液体进行稀释,梯度稀释至 10-9倍,取 200 μL 10-7、10-8、10-9倍的稀释液涂布在牛肉膏蛋白胨、PDA及高氏培养基的平板上,分别置于37、30、30℃内培养。平板划线分离直到划分出单菌落。
1.4.2 产GlcN菌株初筛
将无菌滤纸贴在水琼脂培养基平板上,纯化后得到的菌株点种[28]在滤纸后培养1 d,将滤纸取出晾干进行茚三酮显色[29]。观察点样点及其周围有无紫红色斑点出现,将显色菌株标记并进行菌株复筛。
1.4.3 产GlcN菌株复筛
将复筛的细菌活化后接入牛肉膏蛋白胨培养基,30℃、180 r/min培养16 h。然后按照接种量4%接入牛肉膏蛋白胨培养基,30℃、180r/min发酵培养24 h。复筛的真菌活化后接入PDA培养基,37℃、180 r/min培养20h后以接种量4%接入查氏培养基,于37℃、180r/min发酵48 h。用Elson-Morgon法与LC-MS联用仪分别对初筛菌株的发酵液进行GlcN定量与定性分析。
Elson-Morgon法[30-31]:GlcN 与氨基半乳糖(galcosamine,GalN)两种氨基己糖在100℃和25℃下经碱性乙酰化与对二甲氨基苯甲醛[p-(dimethylamino)-benzaldehyde,PDABA]反应后因乙酰化程度不同而导致生色原含量不同,通过标准曲线分别计算出样品中总氨基己糖与GalN的含量,两者差值即为样品中GlcN的含量。
LC-MS联用条件[32]:发酵液经0.22 μm微孔滤膜过滤。色谱条件为AtlantisT3,5 μm,100 Å(10 mm×150 mm),检测器为Waters 2489 UV检测器。流动相为乙腈-0.01%氨水溶液梯度洗脱,流速为8 mL/min,进样量100 μL。质谱条件为电喷雾正离子化(electron spray ionization,ESI);特定离子(SIM)和质谱全扫检测。温度为600℃;锥孔电压为25 V;扫描范围为质荷比150~250。梯度洗脱条件见表1。

表1 梯度洗脱条件Table 1 The conditions of gradient elution
1.5 菌种鉴定
1.5.1 形态特征观察
通过菌株分离纯化确定曲霉11为产GlcN菌。将目的菌株在PDA固体培养基中进行稀释涂布,30℃恒温培养48 h,观察并记录其菌落特征。
1.5.2 分子学鉴定
将菌株接种于PDA液体培养基中,30℃恒温培养24h后过滤收集菌体。根据Solarbio真菌基因组DNA提取试剂盒提取获得曲霉11的全基因组。将提取的全基因组与南京金斯瑞生物科技股份有限公司合成的引物 ITS1:5’-TCCGTAGGTGAACCTGCGG-3’,ITS4:5’-TCCTCCGCTTATTGATATGC-3’进行PCR扩增。25 μL PCR 反应体系:DNA 模板 1 μL,10×扩增缓冲液(10×PCR buffer)2.5 μL,脱氧核糖核苷三磷酸(deoxy-ribonucleoside triphosphate,dNTP)1 μL,引物 ITS1、ITS4 各1 μL,rTaq 酶 0.5 μL,加超纯水补至 25 μL。PCR 扩增条件为 98 ℃ 2 min,98 ℃ 10 s,58 ℃ 10 s,72 ℃ 10 s,循环 35次,72℃5min。将 3 μLPCR产物与3 μL10×上样缓冲液混合,经0.8%琼脂糖凝胶电泳进行检测,并送至北京擎科生物有限公司进行测序。将测序结果在NCBI中用BLAST在线同源性查询软件与GenBank中已登录的ITS区基因组进行同源性比较,用系统发生推断软件包MEGA5.1进行系统发育树分析,并以Neighbor-Joinging法构建系统发育树,确定该菌株的分类地位。
1.6 曲霉11生长曲线的绘制
曲霉11斜面经PDA液体培养基活化后以接种量4%接入50 mL的PDA液体培养基中,30℃、180 r/min培养,周期取样测干重。以培养时间(h)为横坐标,干重(g)为纵坐标绘制曲霉11的生长曲线。
1.7 曲霉11发酵条件优化
通过对产GlcN菌种的筛选确定曲霉11的发酵液中GlcN含量为1.537 g/L,对曲霉11的发酵条件即发酵周期、发酵pH值、摇床转速、装液量、发酵温度及接种量等方面进行优化以进一步提高发酵液中的GlcN含量并探究发酵优化过程中菌体的干重与发酵液中GlcN含量的关系。
1.8 数据处理
取3次试验数据的平均值并计算其误差,用Origin 8.0软件画图。
2 结果与分析
2.1 产GlcN菌株的分离筛选
2.1.1 菌株初筛结果
共分离得到菌种16株,其中在牛肉膏蛋白胨培养基筛得菌株4种,高氏培养基筛得4种,PDA培养基种筛得8种。采用茚三酮显色法对分离得到的16株菌种进行产GlcN能力的初筛,结果见图1。
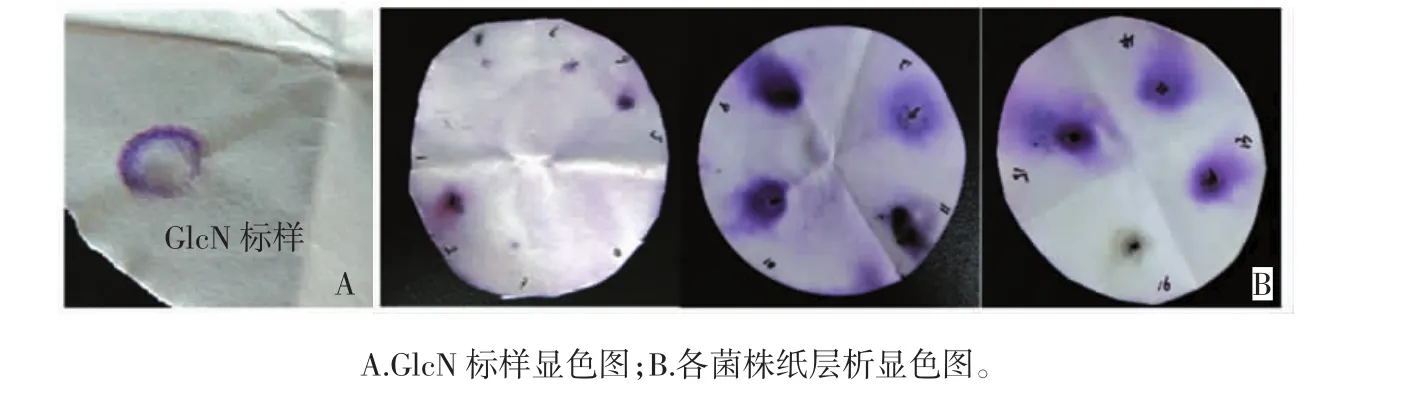
图1 GlcN标样显色图与各菌株纸层析显色图Fig.1 Color diagram of strains and GlcN standard sample
由图1可知,共有10株菌种的周围有紫红色,这表明其产物能与茚三酮发生颜色反应。因此选择这10株菌进行复筛实验。
2.1.2 菌株复筛结果
氨基己糖在100℃与25℃反应下的标准曲线见图2,根据标准曲线计算出各菌株胞外GlcN含量。
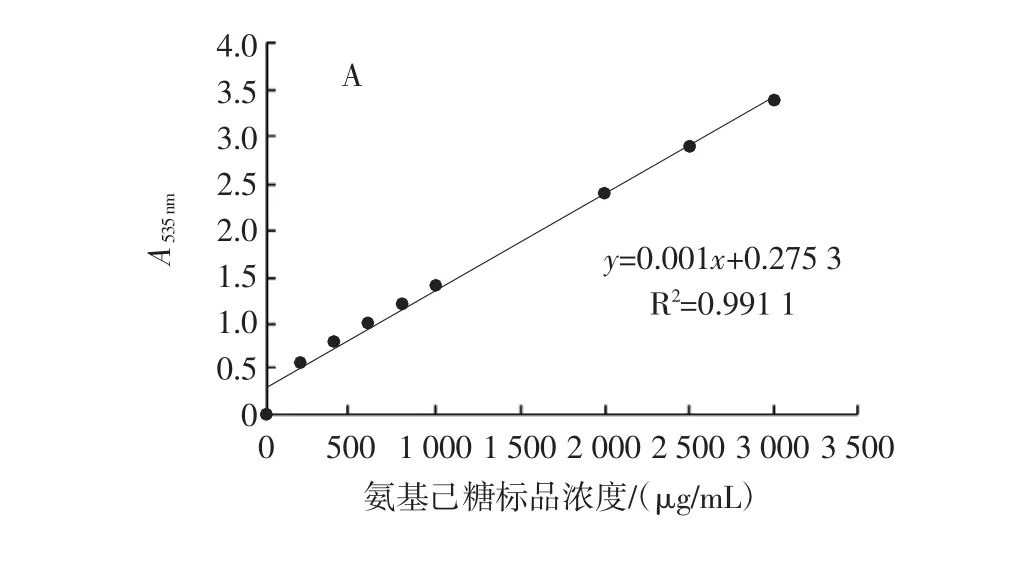

图2 氨基己糖的标准曲线Fig.2 Standard curve of hexosamine
初筛菌株胞外GlcN产量见表2。

表2 初筛菌株胞外GlcN产量Table 2 Yield of extracellular GlcN in selected strains
由表2可知,曲霉11胞外GlcN含量最高,为1.537 g/L,对曲霉11的发酵液进行GlcN定性分析。
用LC-MS联用仪对曲霉11的发酵液进行检测,结果见图3。
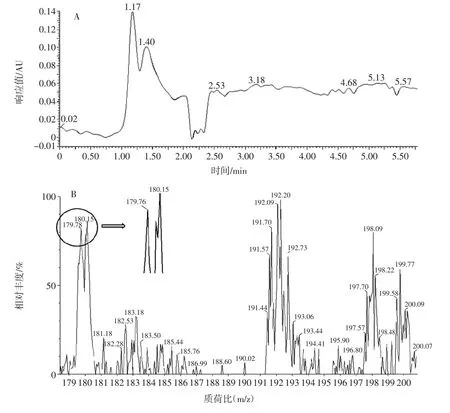
图3 发酵液液相色谱-质谱图Fig.3 Liquid chromatograph-mass spectrometer of fermented liquid
由图3可知,SIR检测出0.9 min~1.3 min内有质荷比为180的离子峰,减去H+的质量1,该时间段检测物质与GlcN标品的分子量179相符,且该时间段质谱出峰时间与液相色谱出峰时间对应良好。
2.2 菌种鉴定结果
2.2.1 形态特征观察
曲霉11菌落形态和显微镜下形态见图4。

图4 曲霉11形态Fig.4 Morphology of the strain 11
如图4所示,曲霉11在PDA固体培养基上30℃培养48 h后,菌落开始为白色,菌落中央逐渐出现很淡的黄色,最后变为粗绒状黑色或黑褐色,背面淡黄色。曲霉11在显微镜下观察菌体菌丝发达多分枝。分生孢子梗自基质中伸出,顶部形成黑褐色放射状球形顶囊。
2.2.2 分子学鉴定
曲霉11的PCR电泳见图5。
由图5比对可得曲霉11的ITS区基因片段大致为600 bp。

图5 曲霉11的PCR电泳Fig.5 PCR electropherogram of strain 11
曲霉11的进化树分析结果见图6。
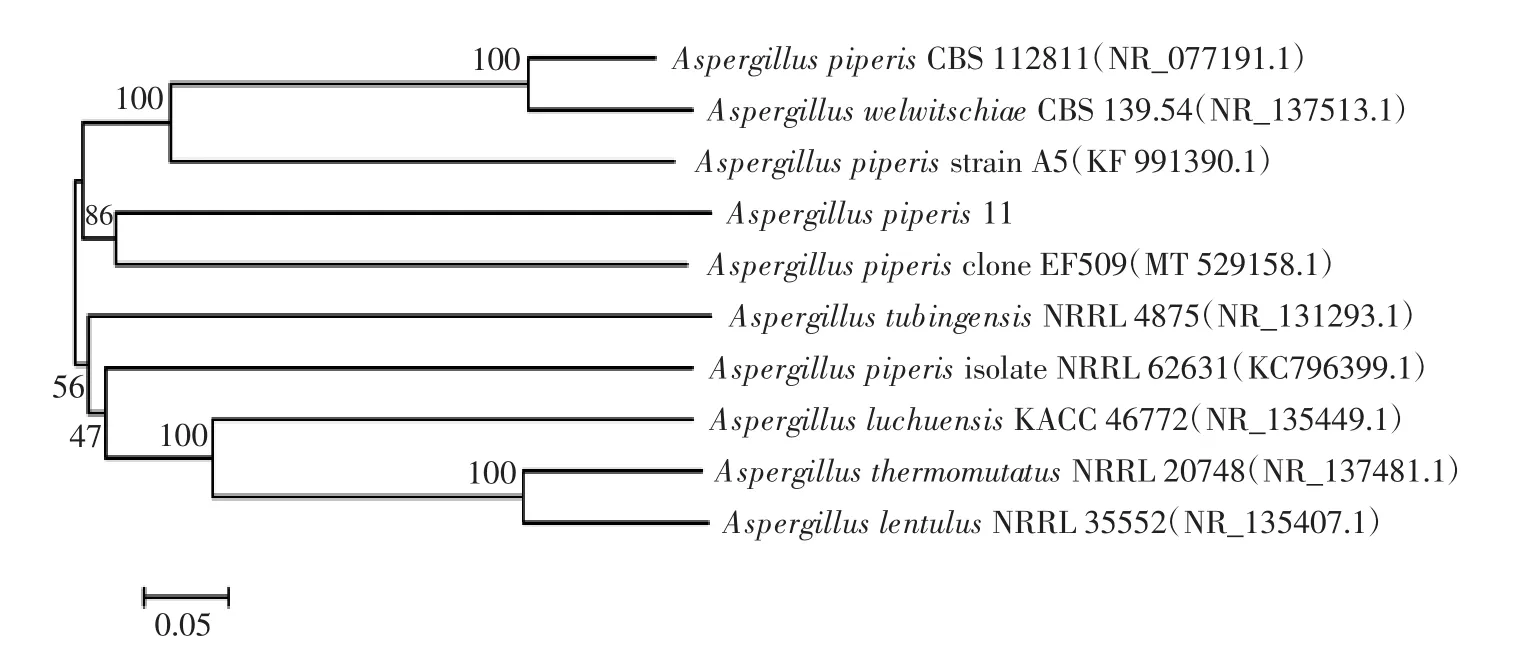
图6 曲霉11的进化树分析结果Fig.6 Phylogenetic tree of strain 11 based on ITS DNA sequences
由图6可知,该菌株的ITS序列与Aspergillus piperis的基因序列同源性最高,为100%。结合该菌株的形态学特征和分子学鉴定结果,可初步判定该菌株为曲霉属。
2.3 曲霉11生长曲线
曲霉11的生长曲线见图7。
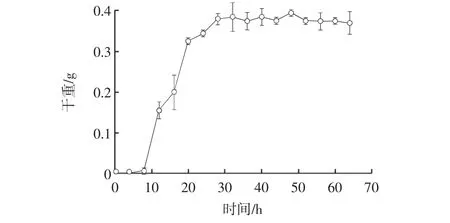
图7 曲霉11的生长曲线Fig.7 The growth curve of Strain 11
如图7所示,8 h~32 h为此菌株的对数生长期,选择生长到第32小时的菌液做为种子转接于发酵培养基。
2.4 曲霉11发酵条件与产GlcN含量的关系
发酵条件对曲霉11产氨基葡萄糖的影响见图8。

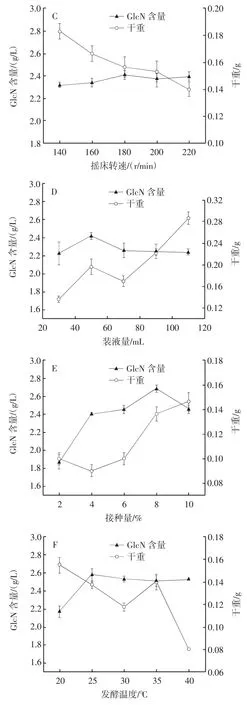
图8 发酵条件对曲霉11产氨基葡萄糖的影响Fig.8 Effect of fermentation conditions on producing glucosamine by Aspergillus 11
由图8A可知,发酵0~12 h时曲霉11发酵液中GlcN的含量急速增长,发酵12 h后GlcN含量迅速减少直至发酵72 h速度减缓。曲霉11的干重从8 h开始增长,到24 h后增长速度减缓直至96 h进入平稳期。发酵过程中测得12 h时发酵液中GlcN含量最高,为2.362 g/L,此时干重为0.16 g,在此基础上进一步优化其他发酵条件。由图8B、图8C、图8D、图8E、图8F可知,随着发酵pH值、摇床转速、装液量、接种量、发酵温度的增加,发酵液中GlcN的含量都先增后减,其中接种量对GlcN含量影响最明显,摇床转速几乎无明显影响,其他条件对GlcN含量影响较小。在优化过程中,装液量与发酵温度对干重的影响比发酵pH值和摇床转速对干重的影响要大。最终确定在发酵时间12 h、发酵 pH3.0、摇床转速 180 r/min、装液量 50 mL(250 mL)、发酵温度25℃、接种量8%时,发酵液中GlcN的含量最高为2.687 g/L,干重为0.134 g。
发酵初期GlcN的大量合成导致干重的迅速增加,随着发酵时间的延长、干重的增加,发酵液中GlcN含量逐渐下降。发酵pH值过低会影响菌株的生长状态以及物质合成速度,当发酵pH值逐渐适于菌株生长时,菌株的干重会逐渐增大,合成的GlcN会大量应用于菌体生长。所以随着发酵pH值的增加,发酵液中GlcN含量逐渐减少而干重增加。高转速产生的剪切力、传质速度、溶氧度和高温等条件都会影响菌体的生长状态以及代谢产物的合成与积累,甚至会导致菌体自溶,因此摇床转速、装液量以及发酵温度会明显影响干重,但几乎不影响GlcN含量。干重与GlcN含量随着接种量的增大而明显增大,但接种量过大会导致GlcN含量的减少且干重增长速度减缓。
3 结论与展望
本文通过茚三酮显色初筛以及Elson-Morgon、液质联用的复筛试验从土壤中分离筛选得到一株胞外产GlcN量为1.537g/L的曲霉11,对该菌株进行形态学观察以及分子学鉴定,将其鉴定为Aspergillus piperis。通过测定曲霉11的生长曲线以及优化曲霉11的发酵条件,最终使得曲霉11在发酵时间12 h、发酵pH3.0、摇床转速180 r/min、发酵温度25℃、接种量8%的条件下GlcN产量为2.687 g/L,是优化前的1.75倍。
在曲霉11发酵初期,细胞内大量合成积累GlcN以用于曲霉11干重的增长,干重的增长相对于GlcN含量的增长具有滞后性而且干重的增长会导致GlcN胞外含量的减少。条件优化时,发现在不利于菌体生长的条件(发酵pH值过低、发酵温度较高)下,菌体的干重虽然较少但胞外GlcN含量并未减少。因此猜想胞外GlcN含量在不利于菌体生长的条件下也会有所提高。
本文根据黑曲霉产柠檬酸发酵早期的发酵液中存在GlcN这一现象,通过菌种筛选及GlcN的定量、定性试验,实现了丝状真菌胞外提取GlcN并通过发酵条件优化提高了胞外GlcN产量。在优化过程中提出了胞外GlcN含量会在不利于菌体生长的条件下有所提高这一新想法,因此后续可从GlcN合成途径的强化、GlcN合成途径支路的抑制、抑制细胞壁合成等不利于菌体生长的条件下进一步提高胞外GlcN的产量。
