肉葡萄球菌表面展示脂肪酶全细胞固定化
2022-12-13王华敏严佳佳尤田薛鲜丽王德培高强
王华敏,严佳佳,尤田,薛鲜丽,王德培,高强
(工业发酵微生物教育部重点实验室,天津市微生物代谢与发酵过程控制技术工程中心,生物工程国家级实验教学示范中心,天津科技大学生物工程学院,天津 300457)
脂肪酶(EC 3.1.1.3)又称三酰甘油水解酶,能够水解长链脂肪酸三酰甘油的一类酶的总称,是常用的生物催化酶制剂之一[1],催化甘油三酯水解生成甘油和脂肪酸[2]。脂肪酶广泛存在于动物、植物组织以及多种微生物中,种类最多的脂肪酶来源于微生物,据统计,产脂肪酶的微生物属包括细菌28个属、真菌23个属[3]。工业用脂肪酶主要来源于微生物[4],具有生产成本低、生产周期短、稳定性好等优点[5],而且其反应条件温和、对原材料要求低、成品质量高[6],不需要辅酶、副反应少、可用于有机溶剂等[7];因此,微生物脂肪酶被广泛地应用于食品、医药及生物柴油等领域[8-9]。脂肪酶虽然在工业上有着广泛的应用,但是大多数游离脂肪酶寿命短且在有机相中不溶解,反应过程中容易结块,且受温度、pH值、溶剂等因素的影响,游离酶的催化活性很低甚至容易失活,这大大降低了酶的水解效率[6,10]。而物理或化学固定化脂肪酶又有着固定化过程复杂、制备成本较高、固定化过程中酶活力或理化性质易损失等缺点。因此,以全细胞的形式利用脂肪酶是降低生物转化成本最有前景的方法之一[11]。
纤维素作为分布最广、含量最多的多糖,是比较廉价的材料,可以作为实用的固定化载体[12]。人们在研究纤维素酶降解纤维素时发现,纤维素酶由纤维素结合域(cellulose binding domain,CBD)和纤维素催化域组成,两者之间通过连接桥连在一起[12-13]。其中,纤维素结合域能够牢固锚定在纤维素底物[14],使邻近的催化域更容易接近底物,促进底物的水解[15]。将表面展示真菌CBD的重组肉葡萄球菌固定于棉花纤维上[16]以及将粪碱纤维单胞菌(Cellulomonas fimi)的CBD锚定在大肠杆菌表面[17]等试验证实了整个细胞与纤维素材料的特异性黏附。根据氨基酸序列、结构和结合特异性,CBD可分为14个家族,大部分属于Ⅰ、Ⅱ、Ⅲ家族,来源于C.fimi的CBD属于Ⅱα家族[18]。
肉葡萄球菌通常被认为是一种安全性的菌株[19],由于其缺乏毒素、溶血素以及极低的胞外蛋白水解活性[20],从而使其成为有吸引力的生产分泌重组蛋白的宿主[21]。本研究通过基因工程的方法,将来源于变形杆菌(Proteus sp.)的脂肪酶基因(lip)和来源于 C.fimi的CBD结合域融合表达并锚定于肉葡萄球菌细胞表面,探究全细胞酶固定化的最适条件及其酶学性质,为后续全细胞固定化脂肪酶的应用提供参考。
1 材料与方法
1.1 材料
1.1.1 质粒与菌种
Staphylococcus carnosus TM300菌株(食品级):德国图宾根大学微生物遗传学研究所Friedrich Goetz教授赠予;表达奇异变形杆菌(Proteus mirabilis)MPE4069菌株脂肪酶(GenBank ID:CP053718.1)的 Escherichia coli-Staphylococci穿梭质粒pBT2-SPP-LIP-XM:天津科技大学生化过程与技术实验室前期构建并保存;E.coli DH5α感受态细胞:天津科技大学生化过程与技术实验室制备保存;粪碱纤维单胞菌(Cellulomonas fimi)的CBD 结合域(GenBank ID:M15823.1):苏州金唯智生物技术有限公司全基因合成。
1.1.2 培养基
LB 培养基(g/L):胰蛋白胨 10.0 g/L、NaCl 10.0 g/L、酵母浸粉5.0 g/L、琼脂粉20.0 g/L(固体培养基加入),1×105Pa灭菌 20 min。
1.1.3 主要试剂与仪器
溶菌酶:北京索莱宝科技有限公司;限制性内切酶HindⅢ、BglⅡ、T4 DNA连接酶:宝生物工程(大连)有限公司;多功能DNA纯化回收试剂盒、高纯质粒小量制备试剂盒:天根生化科技(北京)有限公司;2×Rapid Taq Mster Mix:南京诺唯赞生物科技有限公司;十二烷基硫酸钠聚丙烯酰胺凝胶电泳快速制备试剂盒(12.5%):上海雅酶生物科技有限公司。
pHSJ-4A pH计:梅特勒-托利多仪器有限公司;F-7100紫外可见分光光度计:北京普析通用仪器有限责任公司;Master Cycler Nexus聚合酶链式反应(polymerase chain reaction,PCR)仪:德国 Eppendorf公司;TGL-16高速冷冻离心机:赛默飞世尔(中国)科技公司。
1.2 方法
1.2.1 表达载体pBT2-SPP-LIP-CBD-XM的构建
以前期构建保存的质粒pBT2-SPP-LIP-XM为模板,使用引物 L-F(CCAAGCTTATGAGCACGAAATACCCGATT)和 L-R(GCTCTAGACTAATGGTGATGGTGATGATGAAGCTGTTTGGAGGCAAGATACT)扩增 lip基因;使用引物C-F(GGCTAGCCATGTCCACCCGCAGAACC)和 C-R(CAGATCTGCTTATCGTCG TCATCCTTGT AATCG)扩增CBD基因;以琼脂糖凝胶电泳回收纯化后的lip和CBD片段为模板,使用引物L-F和C-R扩增融合片段lip-CBD基因,下划线为酶切位点。聚合酶链式反应产物经琼脂糖电泳检测,纯化回收并送至华大基因测序分析。用限制性内切酶HindⅢ和BglⅡ分别对质粒pBT2-SPP-LIP-XM和融合片段lip-CBD进行双酶切,纯化回收,Solution Ⅰ连接酶中16℃连接40 min。
1.2.2 基因工程菌S.carnosus TM300/pBT2-SPP-LIPCBD-XM的构建
将重组质粒pBT2-SPP-LIP-CBD-XM用化学转化法转化至E.coli DH5α的感受态细胞中,涂布至含有氨苄青霉素的LB抗性平板上进行筛选,37℃培养12h~16h后,挑取单菌落至上述抗性LB培养液的试管中,温度37℃、180 r/min培养12 h~16 h,取适量菌液提取质粒扩增lip-CBD基因测序分析(测序委托深圳华大基因科技有限公司完成)。选择测序正确的质粒电转至肉葡萄球菌的感受态细胞中,涂布至含有氯霉素的LB抗性平板上进行筛选,37℃培养14 h~16 h后,挑取单菌落至上述抗性LB培养液的试管中,37℃、180r/min培养14 h~16 h,取适量菌液提取质粒,测序分析。
1.2.3 目的蛋白的提取和纯化
挑取单菌落S.carnosus TM300/P-SPP-LIP-CBDXM接种到5 mL含有氯霉素的LB试管中培养,然后按接种量1%转接至含有相同抗性的50 mL LB液体培养基中,37℃、180 r/min培养18 h~24 h。
将发酵后的菌液倒入50mLEP管中,4℃、5500r/min离心10 min收集菌体置于-80℃12 h。收集冰冻的菌体至研钵中,用液氮将其研磨至粉末,收集粉末大约0.2 g,用 1 mL 0.05 mol/L PBS(pH7.4)缓冲液溶解粉末。4℃、12 000 r/min离心15 min,收集上清液提取蛋白。将收集到的蛋白提取液过镍柱纯化,使用80 mmol/L的咪唑洗脱杂蛋白,再使用300 mmol/L的咪唑洗脱目的蛋白并收集,得到纯化后的目的蛋白进行十二烷基硫酸钠聚丙烯酰胺凝胶电泳试验验证纯度。
1.2.4 分光光度计法测脂肪酶酶活力
在一定范围内,OD410随着对硝基苯酚浓度的升高而增长,而对硝基苯酚的浓度又体现了脂肪酶的活力。配制不同浓度的对硝基苯酚(p-nitro phenol,p-NP)溶液,混匀后测定OD410,绘制OD410与p-NP摩尔关系曲线见图1。按照表1的反应体系测定脂肪酶的活性,终止液选用10%的十二烷基硫酸钠(sodium dodecyl sulfate,SDS)。
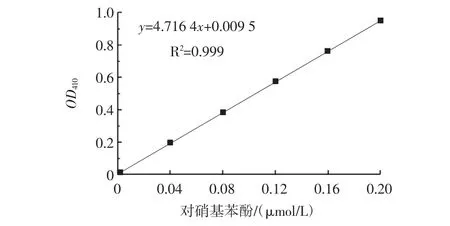
图1 OD410中p-NP的标准曲线Fig.1 The standard curve of p-NP in OD410

表1 测酶活力的反应体系Table 1 Reaction system for measuring enzyme activity
1个酶活力单位定义为室温(25℃)下每分钟释放1 μmol对硝基苯酚(p-NP)所需要的酶量。
1.2.5 高酶活转化子的筛选
对经验证正确的转化子按照1.2.4的方法进行酶活力的测定,从中筛选出酶活力较高的转化子用于后续试验。
1.2.6 溶菌酶处理菌体
挑取单菌落S.carnosus TM300/P-SPP-LIP-CBDXM接种到5 mL含有氯霉素的LB试管中振荡培养18 h~24 h 后,180 r/min 离心 1 min,倒掉上清液,菌体用1 mL PBS洗涤2次。加入1 mL不同浓度的溶菌酶,37℃水浴锅中放置1 h后加入1 mL PBS洗涤2次,收集菌体沉淀测酶活力。
1.2.7 纤维素滤纸固定细胞
按照1.2.3的方法对目的蛋白进行表达后离心收集菌体,用PBS洗涤2次,取适量菌体重悬于缓冲液中至OD600=1.0。在一次性培养皿中加入10 mL菌悬液,再加入一张纤维素滤纸振荡孵育。孵育结束后取培养皿中的菌悬液测定OD600(B),以未加入纤维素滤纸的菌悬液的OD600(A)作为对照。
1.2.8 细胞对纤维素滤纸的吸附率计算方法
细胞对纤维素滤纸的吸附率计算公式如下。
吸附率/%=(A-B)/A×100
式中:A为未加入纤维素滤纸的菌悬液在600nm处的吸光度;B为加入纤维素滤纸的菌悬液在600nm处的吸光度。
1.2.9 单因素固定化条件优化
对收集到的菌体用PBS洗涤2次后按照1.2.7的方法对菌悬液进行孵育,结束后按照1.2.8的方法计算吸附率,分别考察固定化温度(4、16、25、30、37 ℃)、pH值(6、7、8、9、10、11)、时间(2、4、6、8、10 h)和转速(60、80、100、120 r/min)对吸附率的影响。
1.2.10 正交试验
在分别探究温度、pH值、时间和转速对吸附率的影响之后,设计四因素三水平的正交试验,获得较优的吸附条件。正交试验因素水平见表2。
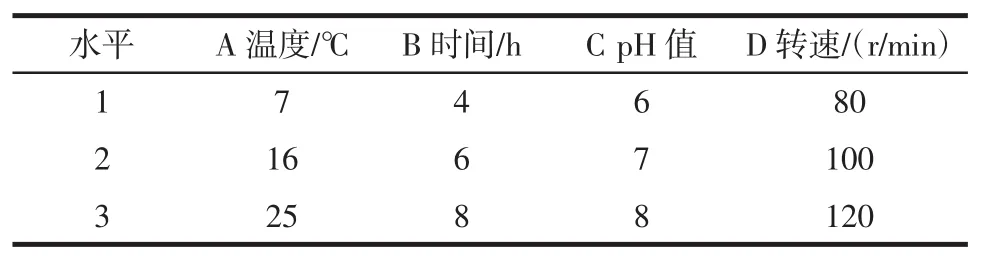
表2 正交试验因素水平Table 2 Factors and levels of orthogonal experiments
1.2.11 固定化细胞稳定性研究
取1.0 g微晶纤维素,用无菌水清洗3遍,与1.2.7处理后的菌悬液在最适的条件下进行固定化。按照酶活力的测定方法,将固定化细胞加入反应体系中测定酶活力,然后1 200 r/min离心1 min倒去上清液,用PBS将固定化细胞在相同条件下多次离心洗涤过滤,除去表面残留未反应底物和反应后的产物,然后将其重新加入到反应体系中测定残留的酶活力,如此反复操作10次,以初始酶活力为100%,考察随着使用次数的增加,固定化细胞表面酶活力的保留情况。
1.2.12 固定化细胞酶学性质探究
按照1.2.10的条件获得固定化细胞,使用不同pH值的缓冲液将其洗涤3次后,40℃下在pH6.0~11.0缓冲液中反应10 min测定酶活力,确定细胞固定化后酶活力的最适pH值。在最适pH值条件下,于30℃~55℃反应10 min测定酶活力,确定细胞固定化后的最适温度。最后分别在不同的温度与pH值下反应3 h,每隔30 min取样测定酶活力,确定细胞固定化后的活力稳定性。
1.3 数据处理
用Excel处理后取平均值,图表采用Minitab软件绘制。
2 结果与分析
2.1 表达载体pBT2-SPP-LIP-CBD-XM构建及其转化子获得
转化子PCR验证见图2。

图2 转化子PCR验证Fig.2 PCR verification of transformants
由图2可知,以纯化后的lip和CBD基因为模板扩增融合片段lip-CBD,经电泳检测扩增的条带约为1 437 bp,符合预期,经测序比对,其结果与模板的基因序列匹配度为100%。分别进行HindⅢ和BglⅡ双酶切后,将融合片段lip-CBD连接至质粒pBT2-SPP-LIPXM,再将连接产物转化至E.coli DH5α感受态细胞中,单菌落PCR验证结果表明质粒pBT2-SPP-LIP-CBDXM构建成功。将该质粒转化到S.carnosus TM300的感受态细胞中,经氯霉素抗性筛选和提质粒PCR验证,测序结果与模板基因序列的匹配度为100%,证明具有pBT2-SPP-LIP-CBD-XM表达载体S.carnosus基因工程菌株构建成功。
2.2 目的蛋白的表达
肉葡萄球菌转化子纯化后lip-CBD的SDS-PAGE见图3。

图3 肉葡萄球菌转化子纯化后融合蛋白lip-CBD的SDS-PAGEFig.3 SDS-PAGE of lip-CBD after purification of S.carnosus transformants
由图3可知,将样品进行镍柱纯化后分析,试验组均只在50 kDa左右出现单一条带,而对照组则未出现任何条带,进一步证明融合蛋白表达成功。
2.3 高酶活力转化子的筛选
不同转化子脂肪酶活力见图4。

图4 不同转化子脂肪酶活力Fig.4 Lipase activity of different transformants
由图4可知,5转化子因为CBD结构域蛋白的导入而对表面展示的脂肪酶的活力有促进作用,其余转化子则因CBD结构域的导入而对酶活力有抑制作用,但促进和抑制的作用并不明显。将5转化子命名为M5,该转化子表面展示脂肪酶的酶活力高达(1 193.41±11.50)U/g湿菌体。
2.4 不同浓度溶菌酶处理M5转化子结果
溶菌酶处理转化子M5后的菌体脂肪酶活力见图5。
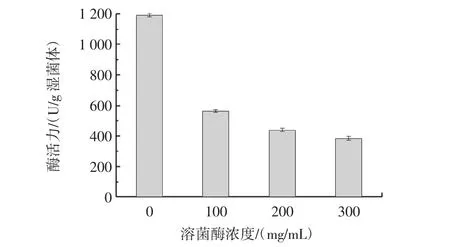
图5 溶菌酶处理转化子M5后的菌体脂肪酶活力Fig.5 Bacterial lipase activity after lysozyme treatment of transformant M5
由图5可知,与无溶菌酶处理的M5转化子相比,用300 mg/mL的溶菌酶处理过的M5转化子的酶活力损失巨大,仅为对照组的32%左右。进而表明溶菌酶对lip-CBD融合蛋白的表面展示影响很大,说明溶菌酶破坏肽聚糖结构而造成表面展示的lip-CBD丢失,证明肉葡萄球菌表面展示lip-CBD成功。
2.5 转化子M5固定化条件分析
对影响固定化吸附率的4个单因素进行分析,结果如图6所示。
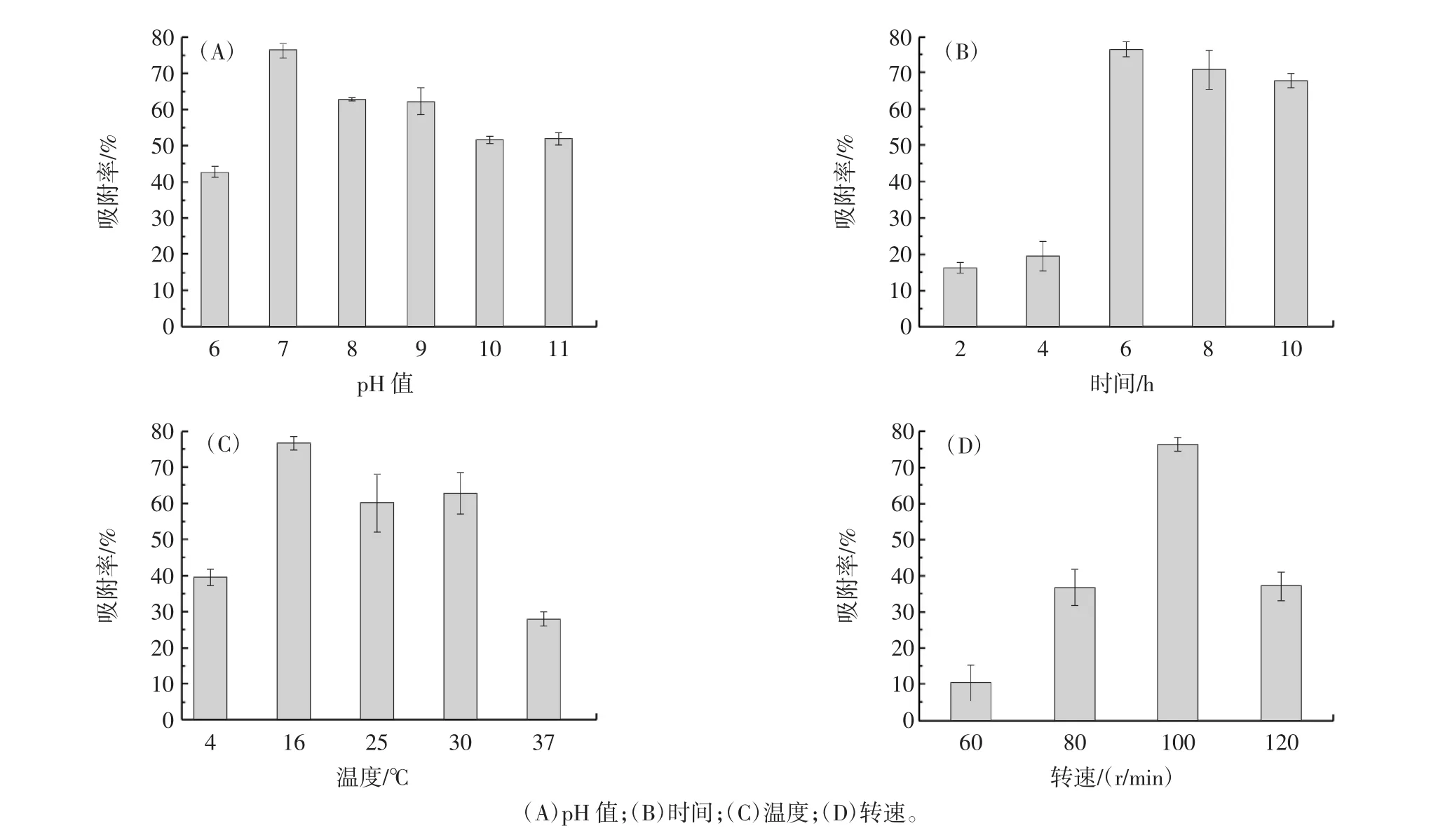
图6 单因素试验结果Fig.6 Single factor test results
由图6可知,随着pH值增大吸附率先上升后下降,pH10.0~11.0时,pH值对吸附率的影响不明显。随着时间的延长吸附率先上升后下降,固定化6 h时,吸附率最高达72%左右,可能由于达到吸附饱和后靠自然吸附的细胞会脱离纤维素基质。当固定化温度为16℃时,吸附率最高。对转速进行单因素分析发现,转速为100 r/min时纤维素基质与细胞的结合最牢固,因此吸附率最高。
2.6 正交试验确定固定化最佳条件
对影响吸附率的4个单因素(温度、时间、pH值、转速)进行四因素三水平的正交试验,结果如表3所示。
由表3可知,纤维素基质吸附细胞的最优条件为pH8.0、时间 6 h、温度 16℃、转速 100 r/min。经验证,在最优条件下纤维素基质对细胞的吸附率可达76%,高于试验9组。
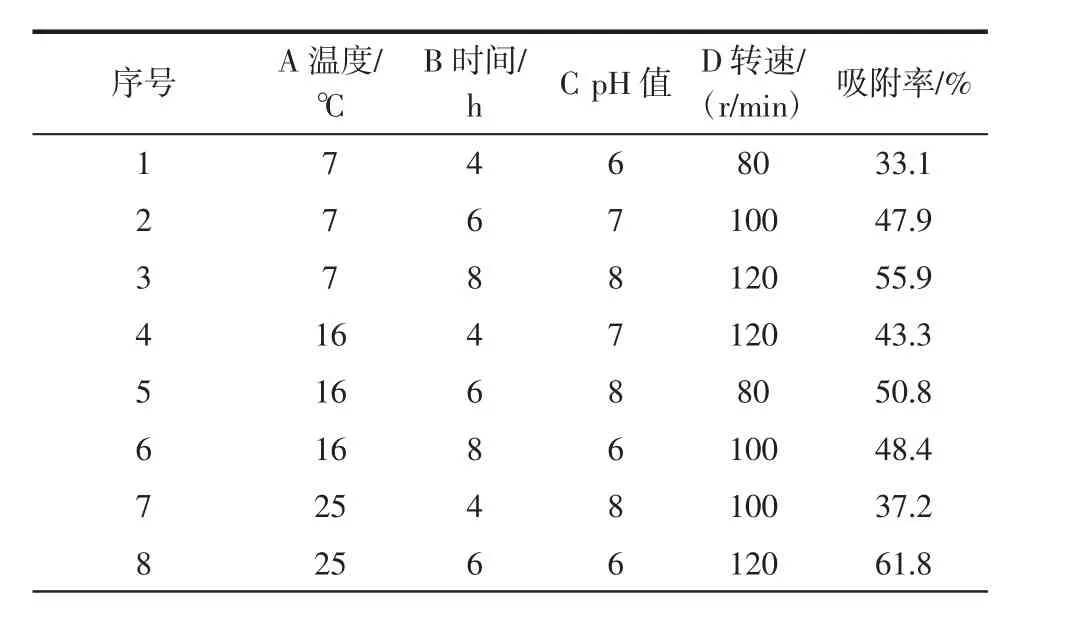
表3 正交试验设计结果及极差分析Table 3 Orthogonal experimental design results and range analysis
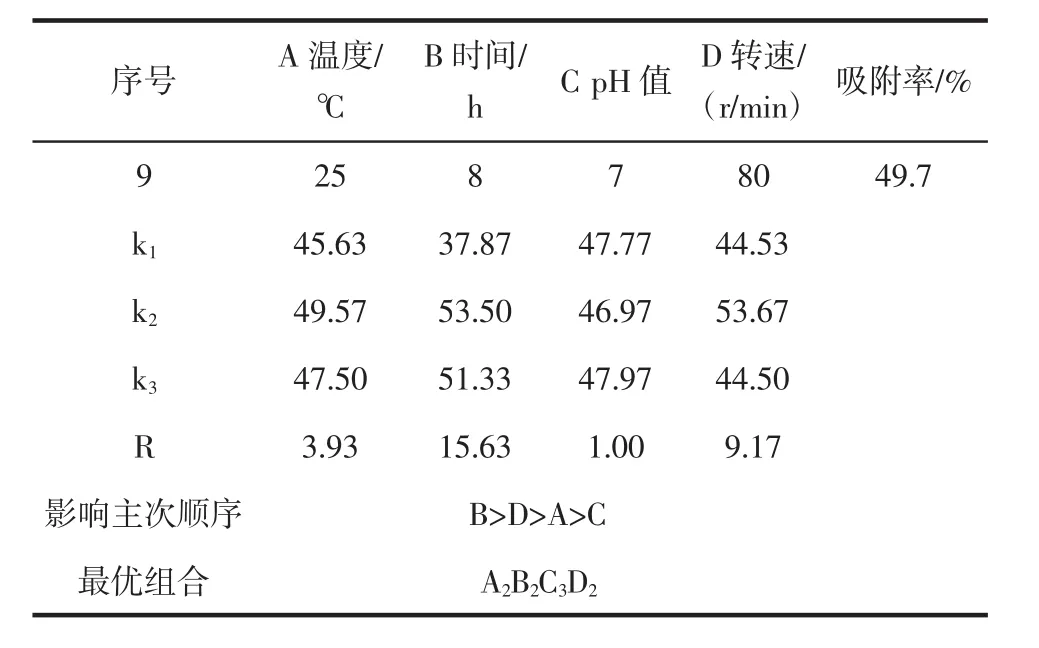
续表3 正交试验设计结果及极差分析Continue table 3 Orthogonal experimental design results and range analysis
2.7 固定化细胞稳定性分析
固定化细胞重复使用次数的分析结果见图7。
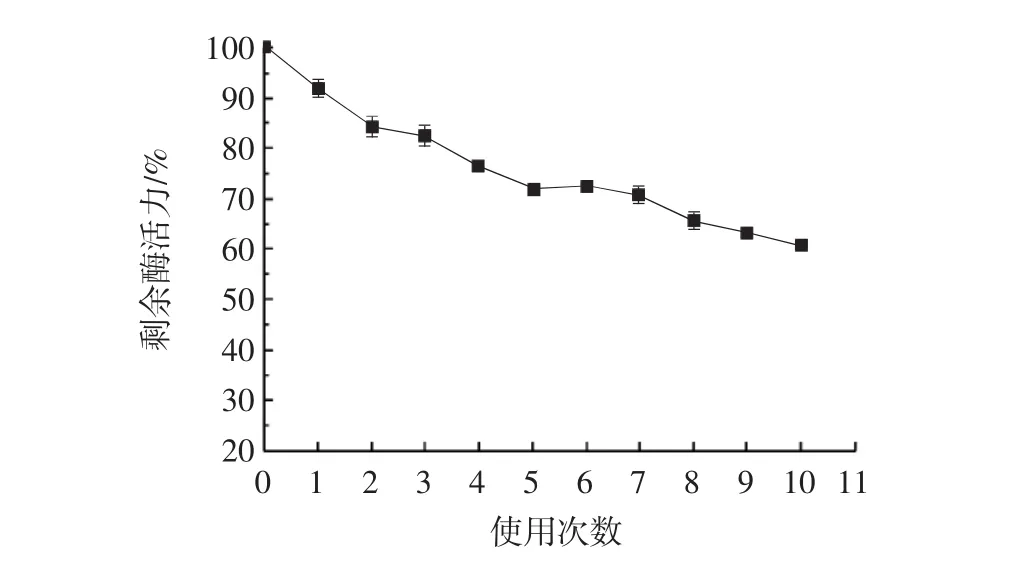
图7 固定化细胞重复使用次数的研究Fig.7 Exploration of the number of repeated uses of immobilized cells
如图7所示,固定到微晶纤维素上的细胞在连续使用10次后酶活力虽然有所下降,但是仍然能够保持原始酶活力的60%左右。可见利用微晶纤维素固定化全细胞酶的效果较好,重复使用率也相对较高。
2.8 全细胞固定脂肪酶酶学性质分析
转化子固定化的酶学性质研究见图8。
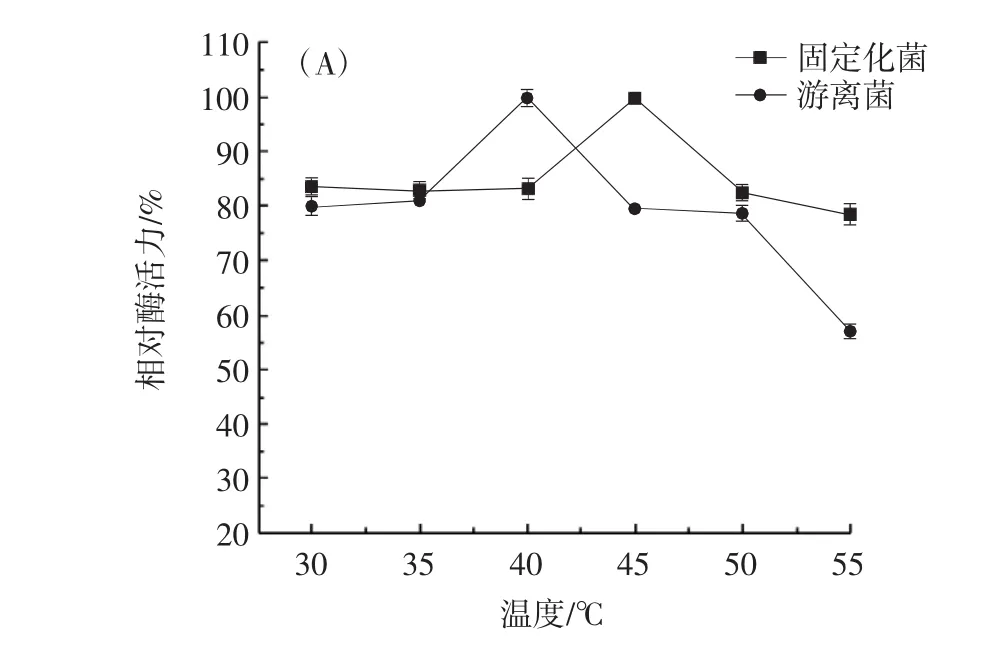
由图8可知,将最高酶活力定义为100%,与游离菌相比,全细胞酶固定化后其脂肪酶的最适温度可达到45℃,比游离细胞的脂肪酶更能够耐受高温,并且在40℃保温3 h后仍能保留80%左右的酶活力,温度稳定性较好;而最适pH值则由10变为9,因为在pH值大于9的缓冲液里,部分纤维素结合域被碱性环境破坏,因此pH值大于9的酶活力明显下降,但其在pH8~9之间稳定性较好,处理3 h后仍能保留70%左右的酶活力。
3 结论
本研究利用食品级的肉葡萄球菌为宿主,将脂肪酶和纤维素结合域共同锚定于细胞壁表面;利用纤维素结合域与纤维素基质的特异性结合以获得全细胞固定化脂肪酶,提高脂肪酶的重复利用率。结果表明:在最优的固定化条件(16 ℃、100 r/min、pH8.0、6 h)下,细胞对纤维素基质的吸附率可达76%,且固定化全细胞酶在连续使用10次后酶活力仍然保留60%左右,固定化后的全细胞脂肪酶更能够耐受更高温度,pH值和温度的稳定性也较好。本研究可明显提高脂肪酶在食品、医药等工业领域的重复利用率,从而降低生物转化的成本。
