矩镰荚苜蓿地下茎次生结构异常发育的解剖学研究
2022-11-26李永强杨艳丽刘济泽张京萍方强恩
李永强,杨艳丽,刘济泽,张京萍,方强恩
(甘肃农业大学 草业学院,草业生态系统教育部重点实验室,甘肃省草业工程实验室,中-美草地畜牧业可持续发展研究中心,兰州 730070)
青藏高原气候寒冷干旱,恶劣环境在影响植物外部形态的同时,还改变了植物的内部结构[1]。植物为了适应高原环境往往会发生组织结构上的改变,例如,高原植物硬叶柳(Salixsclerophylla)主要通过增加叶片厚度来适应高海拔地区的寒旱环境[2]。藏荠(Hediniatibetica)具有发达的木栓层与角质层,是其适应高寒环境的显著特征[3]。苹果属(Malus)的几种高山海棠在响应高原干旱环境时,在解剖结构上会产生多种不同的抗旱特征[4]。植物应对寒冷干旱环境的结构特征不仅表现在地上部分[5],地下器官在逆境下也会发生明显变化[6],其中,一些植物的根茎等部位在极端环境下可能会产生异常分裂的现象。郭伟娜等[7]研究发现秦艽(Gentianamacrophylla)根在生长中会发生分裂,这种分裂会产生加厚的木栓层。侯艳伟等[8]发现干旱缺水的环境会促进棉刺(Potaniniamongolica)的分裂生长。杨瑞丽等[9]认为长叶红沙(Reaumuriatrigyna)茎基部的分裂可以使植株在干旱环境中提高吸水效率。植物在生长过程中产生的分裂现象对其应对恶劣环境具有重要的生态意义。
矩镰荚苜蓿(Medicagoarchiducis-nicolai)别名青藏扁蓿豆[10],广泛分布于青藏高原及其边缘地带的高海拔寒冷干旱地区[11-12],具有发达的地下茎系统[13-14]。矩镰荚苜蓿是青藏高原特有的一种豆科植物[15],其对高寒环境的适应机理尚不清晰。根据上述植物结构特征对逆境的适应性分析,本研究提出一种假设,认为矩镰荚苜蓿也可能通过地下器官产生异常结构来应对青藏高原的寒旱环境。为了验证该假设,本研究采用石蜡切片法对矩镰荚苜蓿不同部位的地下茎进行解剖,观察其初生结构与次生结构的变化,以期在组织解剖学层面探明矩镰荚苜蓿对高原环境的适应机理。
1 材料和方法
1.1 试验材料
试验所用材料为祁连山东段高山草甸(E103°46′,N 37°55′)自然生长的矩镰荚苜蓿。该植物地下横走茎顶端幼嫩部分呈白色,从顶端到基部(接近根颈处),颜色由白色逐渐变为黄褐色、黑褐色,顶端明显粗于基部。
1.2 试验方法
在2021年 6月,随机挖取生长良好的矩镰荚苜蓿多株,剪取生长状态一致的地下茎5根。依据地下茎颜色不同分4段取样:顶端白色茎段、中部黄褐色茎段、中后部黑褐色茎段和最基部茎段(最细部分)。使用双面刀连续切段并编号记录,每段切取长度5 mm,5个重复。切取时避开生长节。
采用常规石蜡切片法制片。材料统一使用FAA(70%乙醇溶液∶冰乙酸∶甲醛=18∶1∶1)固定,低温4 ℃保存。使用乙醇进行梯度脱水,经透明、浸蜡、包埋等过程,选择厚度范围10 μm进行切片。烘片后采用番红-固绿染色,中性树胶封片。使用Motic Panthera U生物显微镜观察分析,Motic Images Plus 3.0软件拍照、测量、处理。放大倍数为10×、40×、100×。
2 结果与分析
2.1 矩镰荚苜蓿地下茎的初生生长
矩镰荚苜蓿地下茎顶端白色幼嫩茎段横切面卵圆形,由外向内依次为表皮、皮层和维管柱。表皮细胞小而紧密,排列整齐(图1,Ⅰ、Ⅲ)。皮层约由10层薄壁细胞构成,细胞较大且充盈,中部皮层细胞大,细胞间隙明显。维管柱横切面整体呈卵圆形,包括髓与维管束。髓射线连接着皮层和中央髓,清晰可见,把维管柱分隔出10个大小不一、形状不规则的维管束,呈环状排列。在初生生长过程中,表皮和髓变化不大。维管束细胞切向和径向生长明显,相邻维管束彼此逐渐靠近,其中有一个维管束往往生长较快,两侧髓射线较宽(图1,Ⅰ、图1,Ⅲ圆圈内)。维管束内部(图1,Ⅱ、Ⅳ),初生木质部生长快于初生韧皮部,其导管增加至2到3层,导管管壁加厚,初生韧皮部变化不明显。

E.表皮;C.皮层;PI.髓;PIR.髓射线;VB.维管束;PVB.初生维管束;PX.初生木质部;PP.初生韧皮部;FC.束中形成层;PE.周皮;IFC.束间形成层;X.木质部;SX.次生木质部;P.韧皮部;MV.后生木质部导管;PF.韧皮纤维;V.导管分子;S.筛管分子;CC.木栓形成层;PH.木栓层;PD.栓内层;CA.形成层。下同;Ⅰ示地下茎距茎尖1.5 cm处白色芽横切结构,圆圈示突出的维管束(Ⅲ同);Ⅱ示Ⅰ的1个维管束;Ⅲ示地下茎距茎尖4.5 cm处白色芽横切结构;Ⅳ示Ⅲ的1个维管束及其髓射线;Ⅰ、Ⅲ圆圈示生长较快的维管束
2.2 矩镰荚苜蓿地下茎的次生生长
次生生长前期,地下茎表皮细胞仍保持小而紧密的正方形状态,但从整体来看,有些部位的表皮逐渐向内凹陷(图2,Ⅰ、Ⅲ),外皮层薄壁细胞不再保持充盈状态,而是发生皱缩,形状极不规则。随着次生生长的进行,凹陷区域及其凹陷深度逐渐增加,皮层进一步皱缩并变薄,皮层细胞扁平。次生生长初期,地下茎的髓变化不大,但其相对于初生结构的比例有所降低。地下茎维管柱次生生长明显,伴随着各维管束的生长连接,维管柱形成一个较大的环状结构。其中,部分韧皮薄壁细胞转化为极发达的韧皮纤维,几乎相连接成环。在初生韧皮部以内的相邻部分,可见4到7个筛管分子(图2,Ⅳ箭头指向)。在木质部与韧皮部之间,束中形成层处于持续分裂分化状态,细胞呈狭长状。形成层内侧,大部分木质部导管分子尚未发育成熟。在维管柱外侧,特殊的环状木栓形成层紧靠韧皮部(图2,Ⅱ~Ⅳ),其细胞逐渐变为狭长状(图2,Ⅳ),约有2到3层,处于持续分裂状态,形成完整的环状。
随着矩镰荚苜蓿地下茎的进一步次生生长,其内部结构发生较大变化。木栓形成层后于维管形成层环出现,在次生生长中逐渐分裂分化,最终形成周皮(图2,Ⅴ箭头指向),伴随着皮层与表皮的脱落,这层周皮将成为地下茎的最外层。切片显示(图2,Ⅴ、Ⅵ),随着维管束次生生长与扩张,地下茎内部变化较大。木栓形成层分裂分化而形成成熟的周皮,周皮外的死细胞不断堆积加厚,形成一圈完整的保护层。在这个次生结构中,由于表皮及皮层的脱落消失,木质部与形成层等的发育及加厚使得维管柱不断扩大,维管柱中可见大面积被染成红色的发达木质部,大体呈扇形(图2,Ⅵ圆圈内)。木质部层层加厚,外侧可见排列整齐的次生木质部导管。中间的次生木质部导管散乱排列,导管较大,管壁明显加厚。髓开始发生变形(图2,Ⅵ)。
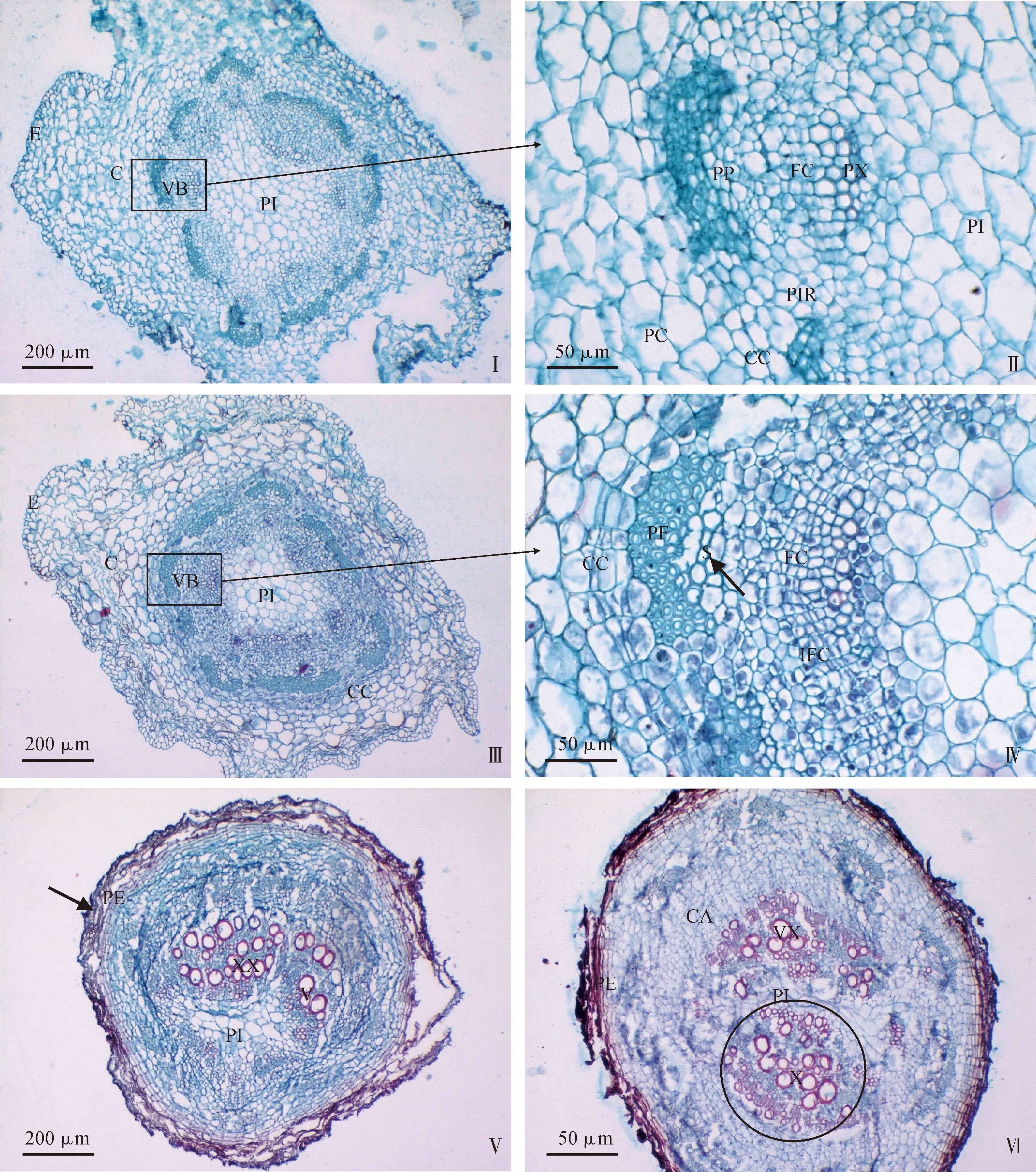
Ⅰ、Ⅲ示地下茎次生生长前期的变化过程;Ⅱ、Ⅳ分别示Ⅰ、Ⅲ的1个维管束的次生生长变化,箭头指向为筛管分子;Ⅴ示地下茎次生生长后期1个维管束正常发育的结构,箭头指向为成熟的周皮;Ⅵ示地下茎次生生长后期2个维管束正常发育的结构,圆圈示1个维管束的发达木质部,可见散乱的大导管
2.3 矩镰荚苜蓿地下茎次生结构的异常发育
矩镰荚苜蓿地下茎周皮的发生方式较为特殊(图3,Ⅰ、Ⅱ)。外周皮来源于内皮层薄壁细胞发育而来的木栓形成层,在地下茎表皮与皮层脱落消失后成为地下茎的最外层,并能进一步次生生长,向内部弯曲延伸。随着地下茎次生生长程度进一步加深,其内部结构开始表现出多个异常现象。通过解剖结构由外到内来看,首先,由于维管束不规则的次生生长,髓被挤压成椭圆状或条状(图2,Ⅴ、Ⅵ)。髓薄壁细胞进一步分化形成了环状的内周皮,部分地下茎随次生生长而产生了裂隙(图3,Ⅴ、Ⅵ)。矩镰荚苜蓿的两种周皮均可卷曲延伸,分裂地下茎,且在此过程中可产生极厚的木栓层(图3,Ⅱ、Ⅲ)。
其次,维管束存在不均衡发育的现象。在次生生长后期,矩镰荚苜蓿的地下茎逐渐变细,其次生结构维管束大小不一,只有少数次生维管束正常发育。切片显示,成熟的地下茎内只有1~3个维管束正常发育(图2,Ⅴ、Ⅵ;图3,Ⅴ),形成明显的次生结构,其余维管束保持着初生状态,正常发育的次生维管束占据其原维管柱的大部分,木质部发达,导管密集。地下茎中心部分的髓均受到挤压开始发生变化,位于次生维管束的中间或一侧(图2,Ⅴ、Ⅵ)。未进行次生生长的维管束的木质部几乎与初生结构时相同,仅见少量较小的木质部导管。
此外,随着上述异常结构进一步生长,可以看到地下茎发生了分裂,这种分裂有两种方式。一种是周皮向内隘缩造成内卷(图3,Ⅰ~Ⅲ),表现为一个维管束超常发育,占据横切面大部,但偏向一侧,另一侧的部分维管束被挤压至维管柱之外,形成层产生大量薄壁细胞填充至二者之间,这些细胞可以分化形成周皮,使得一端或两端的周皮向内凹陷卷曲,不断延伸,将维管束包裹(图3,Ⅰ、Ⅲ箭头指向),并逐渐从外到内分裂为两个大小不一的地下茎(图3,Ⅳ)。另一种是维管柱中心髓的分化造成的分裂(图3,Ⅴ),表现髓薄壁细胞进一步分化产生环状内周皮,并使髓逐渐空心化,产生裂隙(图3,Ⅵ)。内周皮逐渐向外延伸,包裹部分正常发育的维管束,与外周皮相连后,将地下茎从内到外分裂为多部分。这两种分裂方式结果相似,都会将原地下茎分裂为多束。但暂未观察到两种分裂方式同时进行的现象。
3 讨 论
3.1 矩镰荚苜蓿地下茎周皮的特殊发生方式及意义
周皮是由木栓形成层向内分生栓内层、向外分生木栓层所形成的次生组织[16]。通常情况下,茎的木栓形成层发生于邻近表皮的外皮层薄壁细胞,根的木栓形成层一般来源于维管柱鞘[17-18]。在本研究中,矩镰荚苜蓿地下茎的木栓形成层较为特殊,其来源于维管柱外侧、紧邻韧皮部的内皮层薄壁细胞,这与豆科植物甘草(Glycyrrhizauralensis)的地下茎木栓形成层发生方式相似[19]。内皮层木栓化后,将地下茎的维管柱与皮层和表皮隔开,在次生生长过程中皮层薄壁组织逐渐皱缩并最终脱落,木栓形成层分化为周皮,成为地下茎新的外部保护结构。此外,本研究还发现矩镰荚苜蓿地下茎中央髓薄壁细胞也会发生分化,形成木栓形成层,并发育为环状的周皮,周皮内留下裂隙。郭伟娜等[7]在研究秦艽根发育时也发现了类似现象,秦艽根次生生长时,来自根维管柱中心的木薄壁细胞会脱分化产生木栓化细胞,形成内周皮。可以看出,矩镰荚苜蓿地下茎与秦艽一样,在次生生长过程中会产生内、外两种周皮。
一般认为,周皮是植物的次生保护组织,在根、茎次生加厚时替代表皮,作为植物的第一道防线保护植物免受生物和非生物胁迫[20]。闫成才[21]发现典型高山耐寒植物藏荠具有发达的根木栓层与角质层,可以限制热量的传递,以保护植株抵御寒冷胁迫。李小燕等[22]对欧李(Cerasushumilis)的研究也认为发达的周皮能够起到抗寒的作用。Baljeet[23]认为,马铃薯(Solanumtuberosum)块茎周皮木栓化程度的增加保持了块茎的完整性和不透水性,最终减少了蒸腾作用。另有研究表明,内皮层木栓化可以防止水分流失,其木栓化程度的增加可以保护内部维管组织免受干旱胁迫,帮助植株度过干旱期[24]。本研究中,矩镰荚苜蓿外周皮由靠近韧皮部的内皮层细胞形成,内周皮由靠近木质部的髓细胞形成,内、外周皮形成后,完全取代了初生结构中的主要薄壁组织—皮层和髓,形成了由具有发达木栓层的周皮构成的严密的防护系统。从前人对周皮功能的认识来看,这种特殊的周皮防护系统对于青藏高原自然生长的矩镰荚苜蓿而言,具有重要的适应性意义。
3.2 矩镰荚苜蓿地下茎的分裂生长与适应性
植物的根或茎在干旱逆境下会发生分裂(split)[25],这种现象多出现在一些强旱生植物中。分裂具有多种形成方式。龙胆属(Gentiana)植物根的分裂是由于维管柱内部的分裂与周皮向内缢缩共同作用,将根分裂为不同部分[26]。棉刺的根与长叶红沙的茎基部也具有相似的分裂方式,其木质部内侧的木薄壁细胞脱分化形成的异常形成层与断裂的维管形成层相连接并产生周皮,将木质部分割包围[8-9]。半日花(Helianthemumsongaricum)茎基部的木质部会出现内空,并持续延伸分裂[27]。藏锦鸡儿(Caraganatibetica)的根具有周皮缢缩,逐渐分裂的现象[25],这与本研究中矩镰荚苜蓿地下茎的分裂方式相似,不同的是,矩镰荚苜蓿地下茎除外周皮向内缢缩外,维管柱内部也会出现空心化。
前人的大量研究普遍认为植物根茎等器官的分裂是对干旱或其他不良环境的一种适应现象。杨瑞丽等[9]研究发现长叶红沙茎基部的分裂会导致维管束之间相互分离,一个集中的木质部被分割为多个分散的木质部,增加了与土壤的接触面积,这种结构特征可以在干旱环境中提高植株的吸水效率,这与侯艳伟等[8]对棉刺分裂的观察结果相似。王迎春等[28]研究表明,发生劈裂的绵刺通过渗透调节和高的组织弹性两条途径来共同保持膨压,以抵抗不良的生存环境,同时对环境水分胁迫产生较高的敏感性。王继和等[29]认为,绵刺分裂生长是由于严重干旱、较大温差及风蚀沙埋等外部因素作用的结果,分裂生长可以有效应对不良环境[30]。本研究发现,矩镰荚苜蓿具有两种异常的分裂特征,其地下茎通过两种不同的分裂方式来应对高原环境,这在具有分裂特征的植物中较为少见。地下茎分裂产生两种周皮,形成了一套复杂的新防护系统,对地下茎起到保护作用。矩镰荚苜蓿地下茎在次生生长中出现分裂,是该植物对高原冬季漫长的寒旱环境的一种适应性策略。
