光照减少对海草的影响研究进展
2022-11-11廖立国刘应帅陈石泉
符 妙,廖立国,刘应帅,毛 伟,陈石泉,张 翔
(1.海南大学生态与环境学院,海口 570228;2.海南省海洋与渔业科学院,海口 570125)
海草是生活在温带和热带海域中的单子叶植物,是地球上唯一一类可以在海水中完成生活史的高等被子植物[1]。受光照等环境因子的限制,海草大多数分布在浅海海域内[2],分布区域一般在6 m以内的潮间带和潮下带区域,目前记载的海草分布的最大深度为90 m[3]。全球范围内记录的海草种类有6科13属74种[4],我国有4科10属22种[5],占全球海草种类的30.5%。
尽管全球海草的生长面积不到全球海洋总面积的0.2%,但其年平均初级生产量约为1 012 g·DW·m-2,每年的碳封存量达到甚至超过27.4 TgC,相当于全球海洋碳封存总量的10%~15%[6],在全球碳汇中扮演着重要角色。海草不仅能为许多海洋生物(如儒艮Dugong dugon、绿海龟Chelonia mydas及海牛Trichechu等)提供栖息、觅食和繁育的场所[7],还可作为附着基,为附生生物提供足够的生活空间及营养物质[8]。此外,海草能够有效吸附悬浮颗粒物质,减弱海浪能,具有消除污染、改善水质以及维护海岸等作用[8]。但由于人类活动和全球气候变化等影响,全球海草床每年以接近7%的速度在消失,海草生态系统已成为地球上最易受威胁的生态系统之一[9]。
人类活动引起的水体富营养化被认为是干扰海草生态系统的主要方式[10],其可通过降低水体透光率导致海草叶片的光合速率降低,进而影响海草的形态及生理学过程,表现为海草生长速率降低[11]、生物量损失[12]以及非结构性碳水化合物的消耗等[13-14]。本文梳理了国内外关于光照减少对海草生态系统影响的研究现状,对其生长的最低光照要求和光照减少的因素进行了总结与分析,并结合海草在光照减少时的生长与生理变化,探讨海草生态系统对光照减少的响应方式,以期为深入认识人为干扰引起水体透光率下降及未来环境变化对海草生态系统的影响提供参考。
1 海草生长的光照环境
1.1 海草生长的最低光照要求
光照强度对海草的生长、存活和分布起着关键作用[15]。与陆生植物不同,太阳光能在到达海草叶片前会受多种因素影响造成光能损耗,其中空气与水交界处的损失在10%左右,同时受水体溶解物质、悬浮颗粒物质以及水深的影响,太阳光能迅速衰减,因此水下环境易发生光照不足的现象[16]。研究表明,海草生活的海底环境平均要接收海水表面入射辐照度(以下称表面辐照度)约11%的光照才能维持海草的正常生长发育[17]。如波状波喜荡草(Posidonia sinuosa)的正常生长发育要求接收的光照为表面辐照度8%~14%,否则其光合作用将无法顺利进行[18]。此外,不同的海草种类对应的最低光照要求有所差别[19],研究结果详见表1。LEE和DUNTON[20]发现,羽叶二药草(Halodule pinifolia)吸收的光照需要超过表面辐照度的14%才能存活,LONGSTAFF和DENNISON[21]也得到了相似的研究结果。RUIZ和ROMERO[22]认为大洋波喜荡草(Posidonia oceanica)生长所需要的最低光照为表面辐照度的10%~16%,而诺氏鳗草(Zostera noltii)的最低光照要求仅为表面辐照度的2%[15]。造成最低光照要求的这种差异可能是由于种间差异,也可能是由于海草对当地光照环境的适应[23]。
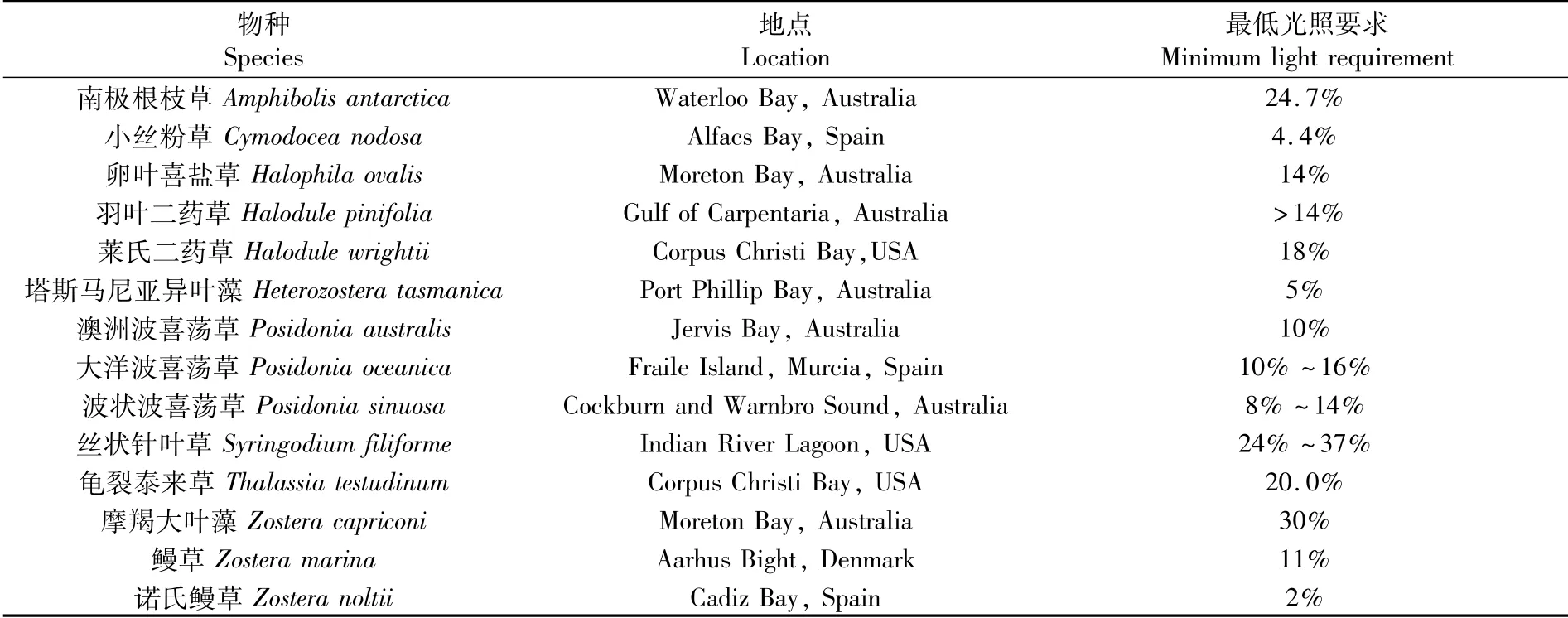
表1 海草的最低光照要求(“最低光照要求”采用表面辐照度的百分比:%SI表示)Tab.1 M inimum light requirement for seagrass(“m inimum light requirement”expressed as percentage of surface irradiance,%SI)
1.2 光照减少的因素分析
影响海草生长的干扰因素主要包括人为活动和自然因素两大类。大规模的海草床衰退主要是由于人为活动干扰引起的光照减少直接(如疏浚)或间接(如富营养化)造成的[24]。人为活动的干扰具体表现为疏浚工程、沿岸码头的修建、海水养殖、生活垃圾与污水的排放等。疏浚工程会造成沉积物悬浮[25],导致海底光照强度减弱,并且可能造成底栖生物群落的掩埋[26],对海草的生长与生存产生负面影响。沿岸码头的修建、船只的停泊等对海草进行了直接的光照遮挡[27]。海水养殖投喂的饵料、生活垃圾与污水的排放等导致海水水体透明度降低,海草可利用的光照减少,从而导致海草的光合速率降低[28]。此外,由于人为活动的干扰,浅海的富营养化环境非常适合浮游植物、附生藻类和大型藻类等大量生长[29],它们对海底弱光的利用率比海草强,生长速率较海草快,不仅会与海草竞争生活空间,还会在海草床上方形成一层加厚的吸光墙,抑制海草的光合作用[30]。这些海洋生物的生长以及自身的分解都会造成水体溶解氧的不足,进而抑制海草的呼吸作用,引起海草的大量死亡[31]。自然因素包括降雨、冻害和台风等也会影响海草的生长。暴雨或长时间的降雨不仅会改变海水的盐度,还会增加海水的浊度,从而限制海草的深度分布[24]。台风引起的风浪不仅能将海草连根拔起,还会造成水体的浑浊,使得台风过后的一段时间内海草仍然受到光照降低的影响[32]。在海洋弱光环境中,光照被认为是限制海草生长与分布的主要环境因素[22]。
2 光照减少对海草的影响
2.1 光照减少对海草生长的影响
在不同光照环境下,海草具有一定的形态可塑性,即海草的形态特征在不同的光照环境下会发生一定的改变[33]。叶片是海草进行光合作用的主要场所,其呼吸速率高,能量消耗大[34]。研究表明,泰来草(Thalassia hemprichii)经过遮光处理(仅接收海水表面入射辐照度1%的光照时)74 d后,其叶长和叶宽显著减小[33]。当遮光率大约为57%和80%时,为期3个月的遮光处理使卵叶喜盐草(Halophila ovalis)的叶长、叶宽和叶面积随着遮光率的增加不断减小,各光照处理组之间具有显著性差异[35]。这种形态的调整有助于降低海草在弱光环境下的呼吸消耗,是其对弱光环境的一种适应[36]。但是也有研究表明,在整个遮光过程中,海草的形态特征没有发生明显变化[37],例如,当处理时间仅为3 d时,卵叶喜盐草的叶片形态在80%的遮光处理下没有发生显著变化[38]。由此可见,研究光照减少对海草生长形态的影响应该结合多种因素进行分析(如遮光持续的时间和遮光强度等),从而更好地理解海草对光照减少的响应。
2.2 光照减少对海草生物量的影响
海草通过光合作用进行有机物质的积累,光照减少使其光合速率降低[39],进而影响其生产力[40],甚至造成植株密度和生物量的减少[41-42]。LONGSTAFF等[43]的研究表明,光照减少导致卵叶喜盐草的生物量迅速下降,在完全遮光24 d后,其生物量降低了64.6%~71.7%。MACKEY等[44]的遮光处理使根枝草(Amphibolis griffithii)实际接收的光照强度大约减少了80%,3个月后其密度和生物量分别较对照组减少了50%和70%。相关研究还表明,遮光处理使地上部分生物量的减少较地下部分生物量要多[33],说明地上部分对遮光的响应较地下部分更加敏感。而FOKEERA-WAHEDALLY和BHIKAJEE[45]对针叶草(Syringodium isoetifolium)进行了为期3个月75%的遮光处理后,无论是夏季还是冬季,遮光区域海草的地上部分密度与对照组没有显著性差异,而且遮光区域海草的生长速率高于对照组。这可能是由于当地光照强度过高,超过了适合海草生长的光照强度范围,从而产生了光抑制现象。
2.3 光照减少对海草光合特性的影响
光合色素在植物的光合作用中起着不可替代的作用。其中,叶绿素能够有效地吸收光能,其含量直接影响海草的光合作用[46]。目前研究的大部分结论是弱光环境能够引起海草叶绿素浓度的增加[47],而且由于叶绿素b对蓝光的吸收能力较叶绿素a强,光照强度的降低会引起叶绿素a/b比值的降低[21]。为了揭示不同光照强度对海草光合作用的影响,SILVA等[13]对鳗草(Zostera marina)和小丝粉草(Cymodocea nodosa)进行了原位遮光处理,其中,75%的遮光处理使两种海草的叶绿素浓度均较对照组显著增加。类似的,当莱氏二药草(Halodule wrightii)可利用的光照强度仅为表面辐照度的10%~13%时,其叶绿素浓度显著高于对照组[48]。光照减少对海草叶绿素浓度的影响还有另一种结论,即过低的光照环境会导致海草叶绿素浓度的减少。LONGSTAFF等[43]的研究表明,卵叶喜盐草对光照的微小变化非常敏感,完全黑暗处理30 d后,其叶绿素浓度较对照处理组显著降低。由此可见,光照强度降低能够促进海草叶绿素浓度的增加,从而有效地吸收海底入射光线[20],提高光捕获效率[18],但是过低的光照强度不利于叶绿素的合成。
另外,光照减少对海草的类胡萝卜素浓度也会产生一定的影响[49]。类胡萝卜素对光系统具有一定的防御和保护作用,还可以将捕获的光能传递给叶绿素。但是目前关于光照减少对海草光合色素的影响研究多集中在叶绿素浓度的变化上,对于其他光合色素受弱光环境的影响研究仍待加强。
2.4 光照减少对海草生态系统固碳能力及碳封存的影响
长期的光照减少可能会造成海草生态系统的退化或丧失[50],从而引起海草床碳储量的降低[51],还可能导致其沉积物中储存的碳由于失去海草冠层的庇护而无法长期稳定存在[52],最终造成海草生态系统的固碳能力降低,甚至使其从碳汇转变为碳源[53]。PENDLETON等[54]结合全球面积、土地利用方式的改变和近地表碳储量对海草床退化引起的碳逃逸进行估算,结果表明在1.5%的全球年退化率下,海草生态系统的碳逃逸高达0.15 Pg(Pg=1015g)。但是目前的研究大多数是关注光照减少对海草床及其沉积物固定的碳量的变化,较少考虑它们退化或丧失时产生的碳逃逸和碳排放。例如,澳洲波喜荡草(Posidonia australis)和大洋波喜荡草两个海草床的大规模损失导致海草床沉积物碳储量分别减少了72%[55]和11%~52%[56],对其进行有效恢复后则可以显著提升海草生态系统的固碳能力[52]。然而一项在坦桑尼亚Chwaka海湾进行的同类实验结果表明,遮光和收割处理对沉积物有机碳含量无显著影响[57]。考虑到全球海草生态系统退化与丧失可能带来的潜在威胁,未来应该深入探究海草生态系统的衰退和损失对其固碳潜力及其碳封存的影响。表2对上述国内外有关光照减少对海草的影响研究进行了汇总。
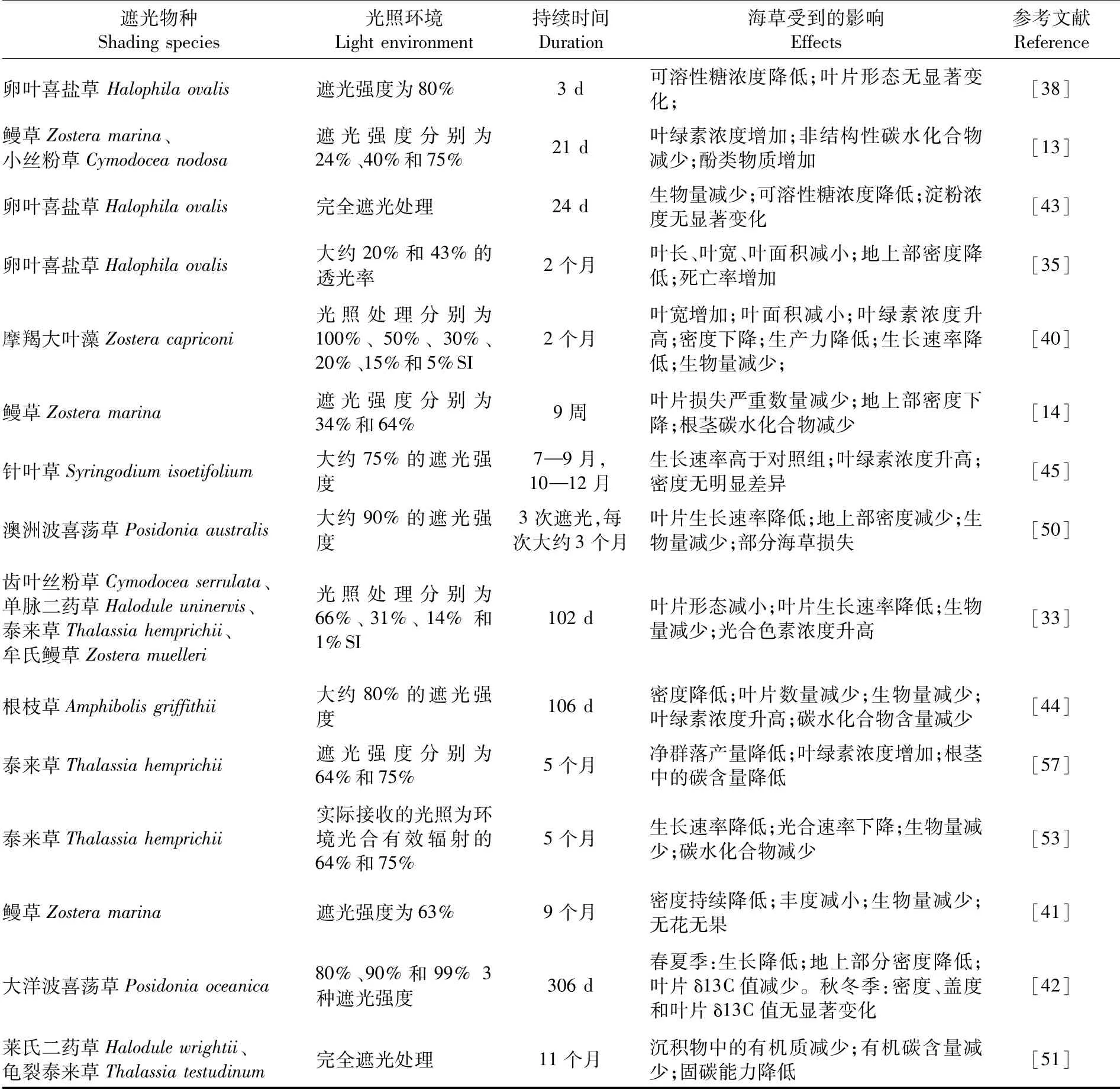
表2 光照减少对海草的影响Tab.2 Effects of light reduction on seagrass
3 海草对光照减少的响应分析
现有的研究表明,海草对光照减少表现出了不同的生长及生理响应。常见的生长响应包括叶片形态、生长速率、植株密度及生物量的变化[40]。在弱光环境下,海草的叶长、叶宽和叶面积通常会有所减小,这能够更好地减少自身的呼吸需求,增加对弱光环境的适应[58]。然而,并非所有的海草物种在不同的光照环境下都具有较高的形态可塑性。形态较小、生长较快的喜盐草属(Halophila)能够通过调整叶片形态使其迅速适应新的光照环境[35]。而形态较大、生长缓慢的波喜荡草属(Posidonia)在光照胁迫环境下对自身形态的调节能力较低,这类海草物种的根茎部分通常含有丰富的碳水化合物储备[59],在光照减少时能够依赖内部碳水化合物的调节来抵御弱光环境。研究表明,波状波喜荡草可以在弱光环境下存活140 d以上[60],其根茎部分的碳水化合物发挥了重要的作用。
光照强度的变化能够直接影响海草的新陈代谢,从而引起生长发育的变化[61]。因此,海草对不同光照环境的生理响应发生在生长响应之前。例如,弱光环境下,海草的叶绿素浓度增加[47]。随着水深的增加,叶绿素a吸收的红光比叶绿素b吸收的蓝光减少得更快,为了更有效地利用海底入射能力较强的蓝光,叶绿素b的增加速度较叶绿素a快[20]。但是当光照环境过低或者过高时,都不利于叶绿素的合成。此外,光照减少还会导致海草光合变量的变化。14 d的遮光处理使卵叶喜盐草和摩羯大叶藻(Zostera capriconi)的最大电子传输速率下降幅度高达40%,饱和辐照度的下降幅度高达70%[39]。
目前关于海草对光照减少的响应研究主要集中在生长和生理指标上,利用分子生物学技术进行的相关研究较少[62]。现有的研究表明,在弱光环境下,大洋波喜荡草的RuBisCo大亚基的表达量下调了30%,而1-果糖-二磷酸醛缩酶、核苷二磷酸激酶及β-淀粉酶等参与碳水化合物分解的酶表现为上调[63]。此外,DATTOLO等[64]研究发现,大洋波喜荡草的光保护基因在弱光环境下的表达量上调,表明海草在光照减少时能够启动光保护响应。
4 展望
目前国内外关于光照减少对海草生态系统的影响研究仍然存在不足,一些较为基础性的科学问题还未得到解决,关于海草在弱光环境下的一系列响应机制也尚不明确。鉴于当前的研究现状,本文提出对未来研究的展望,具体如下:
1)增加研究范围,丰富研究对象
由于种间差异或海草分布地区光照环境等影响,不同海草物种生长的最低光照要求不同,其对光的耐受性也存在一定的差异。增加光照减少对不同种类海草的影响研究,有助于全面地了解不同地区不同海草生长的最低光照要求,完善关于光照减少对海草生态系统影响的基础数据。
2)改进研究方法,加强多种因素交互影响的研究
生物与非生物因素的相互作用会影响海草对光照减少的响应。目前国内外的研究多集中在不同遮光强度对海草生长发育的影响,而将其与遮光持续时间、遮光起始时间和遮光时的水环境温度等其他因素结合的研究很少。单因素的模拟实验容易忽略海草生态系统复杂性,今后的研究可以尝试从多因素的交互影响出发。
3)拓宽研究方向,深化机制研究
关于弱光环境对海草产生的影响,研究学者大多数从生长及生理特性等方面进行阐述,这些研究结果体现了海草细胞水平的变化,而分子水平上的机制研究则很少见。为了更好地评估海草对弱光环境的响应能力,有必要加强海草分子水平的研究,从而了解海草在不同光照环境下的早期变化,进一步探讨海草对光照减少的响应机制,对海草开展多层面的综合研究。
