北方冬麦区小麦春化基因的组成与分布及其与冬春性的关系
2022-11-03刘洪泉傅永斌顾玉章孙丽静胡梦芸张颖君
赵 杰,刘洪泉,赵 芸,杨 锴,3,傅永斌,顾玉章,孙丽静,胡梦芸,李 辉,张颖君
(1.河北省农林科学院 粮油作物研究所,河北省作物遗传育种实验室,河北 石家庄 050035;2.河北宏瑞种业有限公司,河北 石家庄 050035;3.河北工程大学 园林与生态工程学院,河北 邯郸 056038;4.张家口市农业科学院,河北 张家口 075000)
小麦春化作用是指经过适宜的低温处理从而启动或促进小麦开花的现象[1-2]。根据小麦春化过程中对低温的需求强度和持续时间,将其划分为强冬性、冬性、半冬性、弱冬性、弱春性和春性[3-4]。
小麦的春化作用主要由VRN1[5-6]、VRN2[7-8]、Vrn-B3[9]和Vrn-D4[10]基因控制。其中,VRN1位点包括Vrn-A1、Vrn-B1和Vrn-D13个同源基因,分别位于小麦5A、5B、5D染色体上[11],这3个同源基因中任意一个基因为显性时,小麦的发育特性即为春性[12],且显性等位基因Vrn-A1的效应强于Vrn-B1和Vrn-D1[11]。VRN2位点位于小麦5A染色体[7],该位点包括2个串联重复的基因ZCCT1和ZCCT2[8,13]。VRN2可以调控VRN1和Vrn-B3的表达[9]。利用中国春的染色体置换系群体,在小麦7B染色体短臂上定位到控制春化作用的基因Vrn-B3[9],该基因是拟南芥FLOWERINGLOCUST的同源基因,其显性等位基因促进小麦开花,参与小麦春化途径的支路[9]。Vrn-D4位于小麦5D染色体短臂上,通过与VRN1和Vrn-B3的互作参与调控小麦的春化进程[10]。研究表明,VRN1是小麦春化过程中的关键基因[14],且对VRN2基因具有显性上位作用[15]。当3个VRN1基因全为隐性,且VRN2为显性等位基因时,小麦发育特性为春性[15]。
北方冬麦区由北部冬麦区和黄淮冬麦区组成,是我国小麦主产区[16-17]。北部冬麦区主要包括辽宁省南端、北京、天津、河北省长城以南与保定和沧州以北地区、山西省中部与东南部、陕西省北部和甘肃省陇东地区;黄淮冬麦区包括山东、河南、河北省中南部、陕西关中平原、山西省南部、甘肃省天水市及平凉与定西区域、江苏和安徽省的淮河以北区域[18-19]。北方冬麦区约占全国小麦总产的75%左右[19],保障该麦区的小麦产量对我国粮食生产起着至关重要的作用。
本研究以271份北部冬麦区和黄淮冬麦区小麦品种为试验材料,研究4个主效春化基因Vrn-A1、Vrn-B1、Vrn-D1和Vrn-B3的组成特点,阐明两大冬麦区VRN基因组成与小麦冬春性的关系,旨在为小麦品种遗传改良提供理论支撑,对小麦生产具有重要的指导意义。
1 材料和方法
1.1 试验材料
本研究以北方冬麦区271份小麦主栽品种(包括北京14份、天津1份、河北79份、河南71份、山东57份、山西24份、陕西15份、甘肃1份、江苏9份)为试验材料进行4个春化基因的分子标记检测。
1.2 小麦材料田间表型鉴定和冬春性确定
将这271份小麦品种于2020年12月初播种于河北省农林科学院粮油作物研究所三亚试验站,2021年1月下旬调查记录田间抽穗情况。当地气温数据于2345天气预报网(http://tianqi.2345.com/)进行查询。同时,结合中国作物种质信息网(https://www.cgris.net/)和《中国小麦遗传资源目录》的查询结果确定检测小麦材料的冬春性。
1.3 小麦基因组DNA提取和春化基因检测
利用CTAB法提取小麦叶片基因组DNA。春化基因的序列特异性检测参照Yan等[20]、Fu等[21]和Yan等[9]的方法。引物由通用生物系统(安徽)有限公司合成,引物序列见表1。序列扩增试剂采用康为世纪生物科技股份有限公司(https://www.cwbio.com/home)的2×Es Taq MasterMix,PCR扩增程序如下:95 ℃预变性5 min;95 ℃变性30 s,50~61 ℃退火30~60 s,72 ℃延伸1~2 min,35个循环;72 ℃延伸10 min。扩增产物检测利用1%或2%的琼脂糖凝胶电泳,200 V电压电泳30 min,缓冲体系为1×TAE溶液。
2 结果与分析
2.1 小麦品种VRN基因的显隐性分析
2.1.1Vrn-A1基因检测结果 利用引物VRN1AF和VRN1-INT1R对271份小麦品种的基因组DNA进行特异性扩增,结果显示,所有品种均能扩增出734 bp的片段,表明这些品种不含Vrn-A1a或Vrn-A1b等位变异。进一步利用引物Intr1/A/F2和Intr1/A/R3对这些小麦品种的基因组DNA进行扩增,所有小麦品种均未扩增出目标条带,而利用引物Intr1/C/F和Intr1/AB/R均可扩增出1 068 bp的特异性片段,推断供试小麦品种中Vrn-A1第1内含子区域均不含大片段缺失,其Vrn-A1的基因型为隐性vrn-A1。由此表明,271份小麦品种只含有隐性基因vrn-A1。
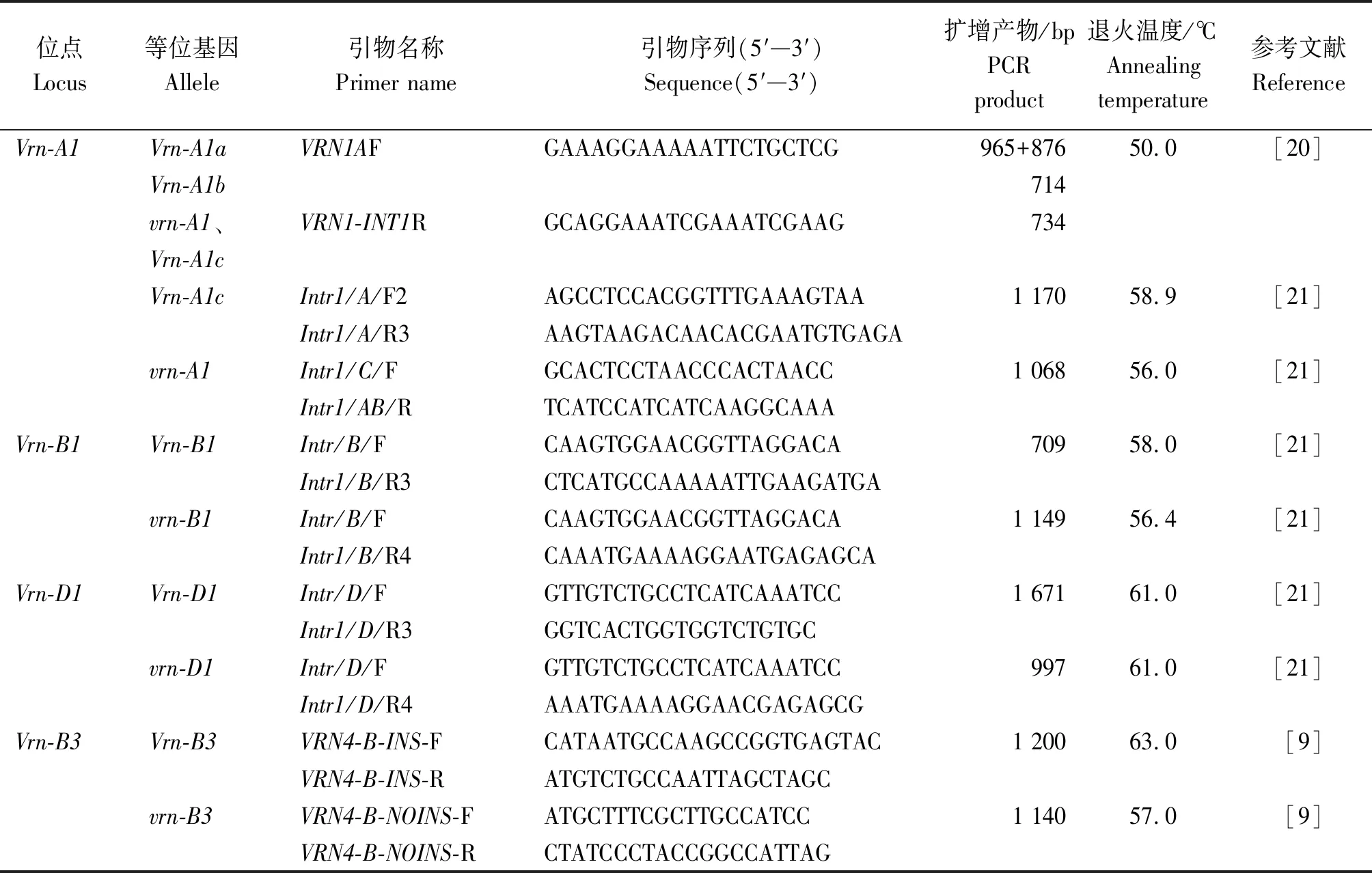
表1 小麦春化基因检测相关引物Tab.1 Primers used for detection of genes responding to vernalization of wheat
2.1.2Vrn-B1基因检测结果 本研究利用引物组合Intr/B/F、Intr1/B/R3和Inr/B/F、Inr1/B/R4对271份小麦品种的Vrn-B1基因组成进行检测。结果显示,有8份品种(豫麦34、郑麦9023、先麦10号、丰川6号、科信9号、陕253、陕农78和西农538)利用引物Intr/B/F和Intr1/B/R3可以扩增出709 bp片段,而引物Inr/B/F和Inr1/B/R4未扩增出目标条带,表明这8份品种含有显性基因Vrn-B1,占供试品种的3.0%。而其余小麦品种只有引物Inr/B/F和Inr1/B/R4可以扩增出1 149 bp片段,引物Intr/B/F和Intr1/B/R3未扩增出条带,表明这些品种含有隐性基因vrn-B1(占97.0%)。
2.1.3Vrn-D1基因检测结果 利用引物Intr/D/F和Intr1/D/R3对这271份小麦品种进行检测,结果显示,有75份小麦材料可以扩增出1 671 bp特异性条带,而引物Intr/D/F和Intr1/D/R4未扩增出条带,表明这些小麦品种含有显性基因Vrn-D1(占27.7%)。其余的196份小麦材料检测结果正好相反,表明这些材料含有隐性基因vrn-D1(占72.3%)。
2.1.4Vrn-B3基因检测结果 对271份小麦材料Vrn-B3位点进行检测,结果显示,所有品种利用引物VRN4-B-INS-F和VRN4-B-INS-R均未扩增出目标条带,而引物VRN-4-B-NOINS-F和VRN-4-B-NOINS-R都扩增出1 140 bp特异性条带,表明这些材料均含有隐性基因vrn-B3。
2.2 小麦品种不同春化基因的组合和分布情况
通过分析供试小麦材料春化基因显性位点的分布情况,表明显性等位基因Vrn-D1分布频率最高,为27.7%;显性等位基因Vrn-B1分布频率仅为3.0%;所检测材料中均不含显性等位基因Vrn-A1和Vrn-B3(表2)。以上结果表明,供试小麦品种中控制春化作用的显性等位基因主要是Vrn-D1,其中在江苏小麦品种中分布频率最高(88.9%),其次为河北小麦品种(38.0%)、陕西小麦品种(33.3%)、河南小麦品种(31.0%)、山西小麦品种(12.5%)、山东小麦品种(10.5%)、北京小麦品种(7.1%)。显性等位基因Vrn-D1在北方冬麦区分布频率呈现由黄淮冬麦区向北部冬麦区、由南向北逐渐减少的趋势。显性等位基因Vrn-B1只有在3个省份的小麦材料中检测到,分布频率为陕西最高(20.0%),其次依次为河南(4.2%)、山东(3.5%)。所检测小麦材料中甘肃和天津的品种各有1份,均不含显性等位基因Vrn-B1和Vrn-D1。
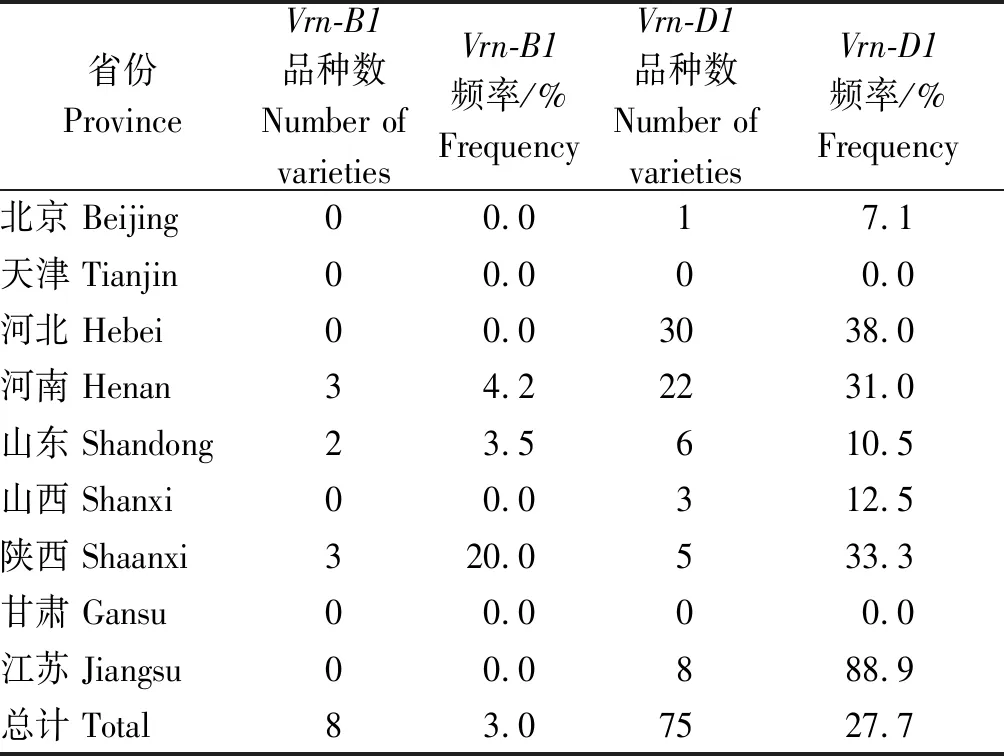
表2 显性等位基因Vrn-B1和Vrn-D1在不同省份小麦品种中的分布Tab.2 Distribution of dominant alleles Vrn-B1 and Vrn-D1 in wheat varieties of different provinces
该271份小麦材料共检测到春化基因4种组合,分别为vrn-A1/Vrn-B1/vrn-D1/vrn-B3、vrn-A1/vrn-B1/Vrn-D1/vrn-B3、vrn-A1/Vrn-B1/Vrn-D1/vrn-B3、vrn-A1/vrn-B1/vrn-D1/vrn-B3。所检测小麦材料中基因型为vrn-A1/vrn-B1/vrn-D1/vrn-B3的品种有190份,其次依次是基因型为vrn-A1/vrn-B1/Vrn-D1/vrn-B3的品种(73份)、基因型为vrn-A1/Vrn-B1/vrn-D1/vrn-B3的品种(6份),基因型为vrn-A1/Vrn-B1/Vrn-D1/vrn-B3的品种只有2份。
2.3 北方冬麦区小麦春化基因组成与冬春性的相关性分析
2020年12月将这271份小麦材料播种于海南省三亚市,2021年1月调查田间抽穗情况。该生长周期,三亚市最低气温高于11℃(图1),因此,田间正常抽穗的品种为春性小麦。
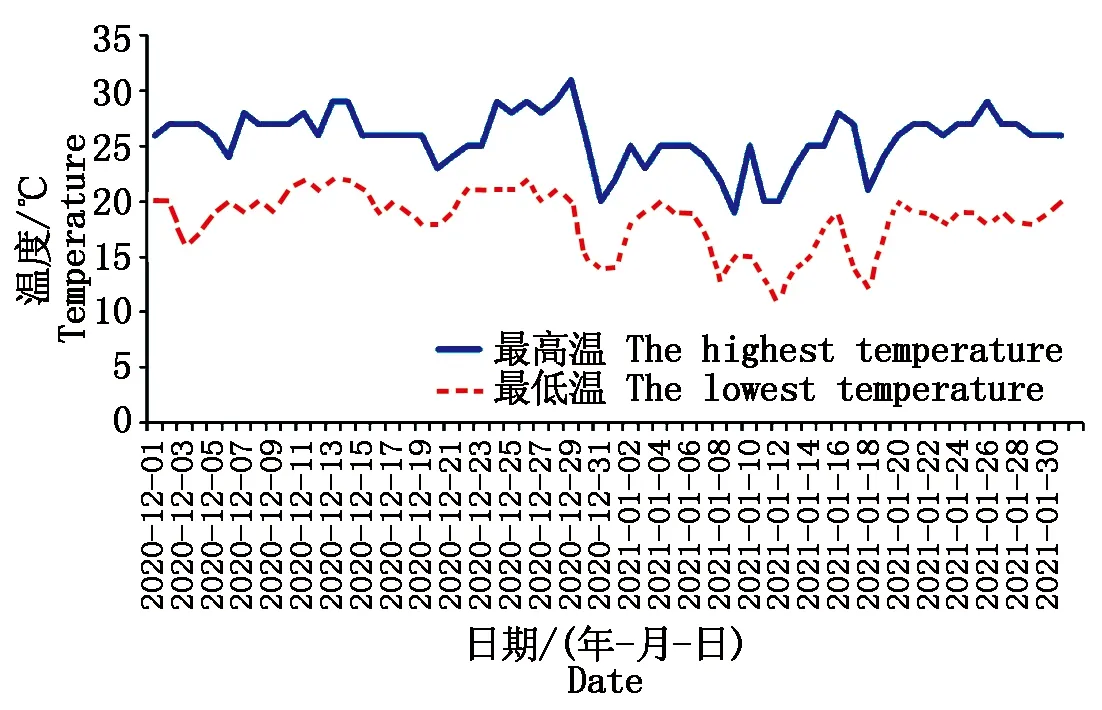
图1 三亚市气温变化Fig.1 Temperature change in Sanya City
进一步结合资料记载和表型调查结果,所检测材料中偏冬性和半冬性品种最多,有184份,其次依次为冬性品种(53份)、弱春性品种(15份)、弱冬性品种(10份),春性品种只有9份。
通过分析春化基因与小麦冬春性的关系,显示在含有显性春化基因Vrn-B1的8份品种中,半冬性品种3份、弱春性2份、春性3份,Vrn-B1基因型推测的表现型与冬春性的一致性为62.5%(表3)。含有显性春化基因Vrn-D1的品种冬春性记载以半冬性为最多(53份),其次为冬性(8份)、弱春性(7份)、春性(6份)、弱冬性(1份),Vrn-D1基因型推测的表现型与冬春性的一致性仅为17.3%(表3)。供试小麦材料主效春化基因组成为全隐性(即基因型为vrn-A1/vrnB1/vrn-D1/vrn-B3),冬春性记载中半冬性和偏冬性为最多(129份),其次为冬性(45份)、弱冬性(9份)、弱春性(6份),春性品种有1份,基因型推测的表现型与冬春性的一致性为96.3%。
以上结果表明,显性等位基因Vrn-B1和Vrn-D1推测的表现型与冬春性的一致性较低,隐性等位基因组合vrn-A1/vrn-B1/vrnD1/vrn-B3推测的表现型与记载的冬春性一致性较高。产生这一现象的原因可能是显性等位基因Vrn-B1和Vrn-D1对小麦的春化作用较弱,或在这些小麦材料中含有其他未检测的春化基因。
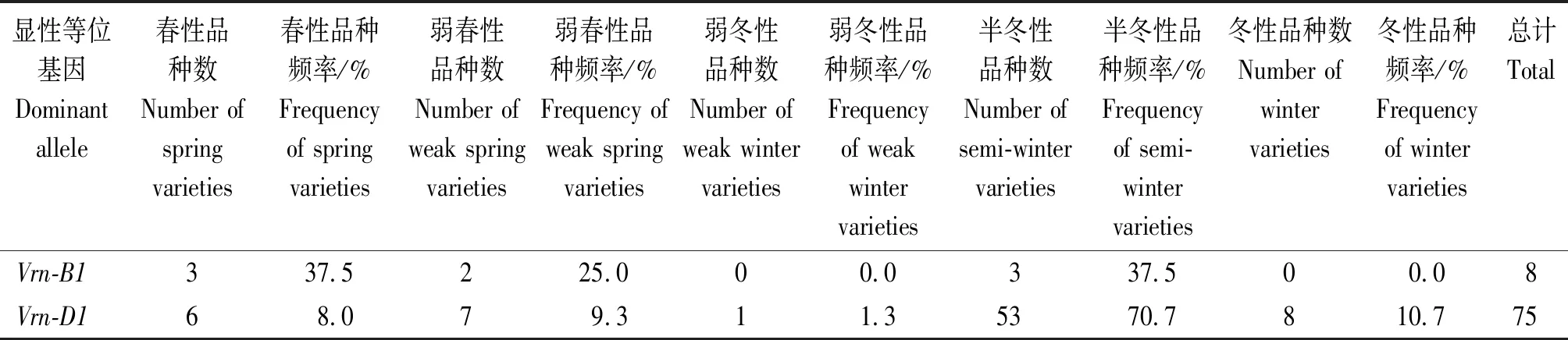
表3 显性等位基因Vrn-B1和Vrn-D1在不同冬春性品种中的分布Tab.3 Distribution of dominant Vrn-B1 and Vrn-D1 allele in different winterness-springness types
3 结论与讨论
Yan等[6]通过利用单粒春小麦G2528和冬小麦G1777构建F2群体,发现VRN1与编码MADS-box转录因子的基因AP1和AGLG1完全连锁。进一步通过序列分析,发现AP1启动子区的自然变异导致小麦的冬春特性不同[6],并根据其启动子区的序列差异开发出Vrn-A1基因在六倍体冬小麦和春小麦中的特异性检测引物[20]。通过对春性小麦品种完整VRN1基因进行测序,发现其显性等位基因VRN1在第一内含子区有4 kb的片段缺失,其中2.8 kb序列在隐性等位基因vrn1中具有高度保守性,并由此开发出春化基因VRN1的特异性检测引物[21]。Yan等[9]研究发现,春性小麦品种在Vrn-B3的启动子区存在一个逆转录转座子插入,并根据该插入缺失设计出小麦冬春性特异性检测引物。基于前人研究结果,本研究利用冬春性小麦VRN1和Vrn-B3启动子区或内含子区插入缺失设计的特异性引物对271份北部冬麦区和黄淮冬麦区小麦主栽品种进行分子标记检测,结果显示,所检测品种中均不含显性等位基因Vrn-A1和Vrn-B3。
中国小麦地方品种中4个主效的春化基因分布频率和分布地域差异较大。显性等位基因Vrn-A1和Vrn-B1主要出现在东北和西北等春麦区;显性等位基因Vrn-D1分布频率呈现从南向北依次递减趋势;而显性等位基因Vrn-B3在中国小麦品种中分布频率极低[14,22]。姜莹等[14]对中国小麦地方品种VRN1基因进行检测发现,中国小麦地方品种中显性春化基因Vrn-D1分布频率较高,达60.8%;而显性等位基因Vrn-B3在所检测的品种中分布频率为0。李哲等[22]对中国小麦主产区品种春化基因进行分子检测,发现显性等位基因Vrn-D1分布频率高达44.6%,明显高于显性等位基因Vrn-B1(18.1%)和Vrn-A1(11.2%),显性等位基因Vrn-B3分布频率仅为0.8%。郭总总等[23]对黄淮麦区91份小麦材料进行分子标记检测,发现所检测材料中不含显性等位基因Vrn-A1和Vrn-B3,显性等位基因Vrn-D1分布频率为41.8%,明显高于显性等位基因Vrn-B1(6.6%)。游银等[24]对黄淮麦区部分骨干亲本的春化基因进行分析,发现所检测材料中显性春化基因Vrn-D1分布频率为20.3%,显性基因Vrn-B1分布频率为4.7%,未检测到显性基因Vrn-A1和Vrn-B3的分布。本研究结果显示,显性等位基因Vrn-D1在271份供试小麦品种中分布频率为27.7%,其次为Vrn-B1(3.0%),Vrn-A1和Vrn-B3没有分布,这可能是冬麦区小麦品种对春化作用的需求较强造成的。本研究结果与前人研究结果基本一致,说明显性春化基因Vrn-D1和Vrn-B1是控制北部冬麦区和黄淮冬麦区小麦冬春性的主要基因。
据报道,冬麦区是小麦春化基因的基因型推测表现型与实际表现型一致性较差的区域,因此,进一步分析该麦区春化基因对小麦冬春性的影响尤为重要。姜莹等[14]研究发现,我国春麦区春化基因推测的冬春性与实际小麦的冬春性一致性较高,冬麦区的一致性较低。赵彦坤等[25]对黄淮麦区北片小麦材料进行检测发现,春化基因鉴定的基因型与表现型一致性较低,主要与Vrn-D1的显性变异有关。本研究检测了4个春化基因位点(Vrn-A1、Vrn-B1、Vrn-D1和Vrn-B3)在小麦品种的分布,春化基因组合vrn-A1/vrn-B1/vrn-D1/vrn-B3和vrn-A1/Vrn-B1/vrn-D1/vrn-B3推测的表现型与冬春性的一致性较高,vrn-A1/vrn-B1/Vrn-D1/vrn-B3推测的表现型与冬春性一致性较低,其原因可能是由显性春化基因Vrn-A1、Vrn-B1和Vrn-D1对低温春化的敏感性和需求不同造成的[14],显性春化基因Vrn-B1对小麦春化发育特性的作用强于Vrn-D1、弱于Vrn-A1。由此可见,部分含有显性等位基因Vrn-D1的材料需要春化才能开花结实,而包含显性等位基因Vrn-B1的小麦材料对春化的需求明显较弱。北部冬麦区和黄淮冬麦区小麦材料中可能存在其他控制春化作用的基因。
