夏玉米粒位效应对增密的响应及其碳氮代谢特征
2022-11-03翟立超张丽华郑孟静吕丽华申海平姚海坡贾秀领
翟立超,张丽华,郑孟静,吕丽华,申海平,姚海坡,贾秀领
(河北省农林科学院 粮油作物研究所,河北省作物栽培生理与绿色生产重点实验室,河北 石家庄 050035)
玉米是我国第一大粮食作物,作为粮、经、饲兼用的多用途作物,玉米生产对我国粮食安全与整个国民经济发展具有重要的影响。河北夏玉米区位于黄淮海夏玉米区的最北端,该区玉米平均种植密度为6.27万株/hm2[1],较低的种植密度是限制当地玉米产量潜力发挥的主要因素。随着玉米品种耐密性的增强,合理增加种植密度已成为当前提高玉米产量的重要途径[2-4]。由于玉米籽粒发育普遍存在一定程度的粒位效应(即与玉米果穗中、下部强势粒相比,上部弱势粒灌浆速率慢,灌浆充实不充分,最终粒质量低)[5],种植密度的增加往往会进一步加剧这种粒位效应,限制密植玉米产量潜力的发挥[6]。因此,深入分析增密对玉米粒位效应的影响,阐明增密影响玉米粒位效应的生理机制,对今后采取有效措施缓解密植玉米粒位效应、实现玉米密植高产挖潜具有重要的意义。
碳、氮代谢是作物植株体内最基本的两大代谢过程,二者的协调对籽粒发育和产量形成至关重要[7-8]。前期研究表明,蔗糖-淀粉合成相关酶活性低是导致籽粒灌浆充实差的主要原因[5,9]。但也有研究显示,籽粒中氮代谢关键酶活性也影响籽粒的发育与灌浆[10]。 种植密度对玉米籽粒灌浆影响已有不少报道[11-13],但这些研究多数是围绕玉米中部强势粒为对象得出的结论,关于种植密度对玉米不同粒位籽粒灌浆的影响及其碳氮代谢生理特征的研究则鲜有报道,这限制了人们对增密影响玉米粒位效应生理机制的深入理解。
本研究拟通过大田试验,设置不同的种植密度,重点探讨夏玉米粒位效应对增密的响应及其碳氮代谢生理特征,以期为河北夏玉米密植高产挖潜的栽培调控途径提供理论依据与技术指导。
1 材料和方法
1.1 试验地概况
试验于2020—2021年在河北省石家庄市藁城区的河北省农林科学院粮油作物研究所堤上试验站进行(37°95′N,114°71′E)。该地区属于暖温带半湿润大陆性季风气候,年平均气温12.8 °C,年平均降雨约484 mm,其中约70%的降雨集中在6—9月,年均日照时数为2 711 h。试验地土壤类型为沙质壤土,试验前耕层土壤(0~20 cm)的有机质含量为20.04 g/kg,全氮0.81 g/kg,全磷0.60 g/kg,全钾7.24 g/kg,碱解氮20.16 mg/kg,速效磷64.12 mg/kg,速效钾178.38 mg/kg。
1.2 试验设计
以玉米品种豫单9953为供试材料,设置3个不同的种植密度(PD1:6.0万株/hm2;PD2:7.5万株/hm2;PD3:9.0万株/hm2),每个处理设3次重复,共9个小区,小区面积76 m2,等行距种植,行距为60 cm,株距按照不同密度计算确定。肥料施用采用沃夫特缓控释复合肥(N-P2O5-K2O,28-8-8,养分总含量≥44%)全部基施,施用量810 kg/hm2。基肥于小麦收获灭茬后玉米种植前通过人工均匀撒施于每小区,然后旋耕入0~20 cm耕层土壤,采用人工点播方式进行。2020,2021年播期分别为6月14日和6月13日,播后浇蒙头水。其他田间管理措施同常规高产田。
1.3 测定项目及方法
1.3.1 籽粒灌浆动态及灌浆速率 吐丝前,每小区选取生长一致的单株挂牌并将雌穗套袋120株,散粉高峰期取下套袋辅以人工饱和授粉。授粉后至生理成熟期,每5 d取挂牌果穗6个,并分离果穗上部中间3~6环籽粒(弱势粒)和果穗中部中间3~6环籽粒(强势粒),数取100粒经 105 ℃杀青30 min,80 ℃烘箱烘至恒质量后称质量,测定籽粒增质量动态。
以授粉后天数为自变量,测算出的单粒粒质量为因变量,参照朱庆森等[14]的方法,用Richard方程拟合籽粒的增质量动态:
①
进一步求导数得出籽粒灌浆速率(G):
②
其中,W为籽粒的实际质量,a为籽粒的终极生长量,t为授粉后的天数(d);b、c、d为回归方程所确定的相关灌浆参数。
1.3.2 植株干物质积累量测定 在开花期和生理成熟期测定干物质积累,每小区取生长均匀的植株3株,按茎、叶、鞘、苞叶、籽粒、穗轴6个部分进行分解,并采用烘干称质量法测定。
花后物质积累量=生理成熟期的干物质积累量-开花期的干物质积累量
③
1.3.3 籽粒碳氮代谢相关酶活性的测定 在授粉后30 d取不同处理果穗的强、弱势粒,置于液氮中冷冻5~10 min后放入-70 ℃超低温冷冻箱保存,用于测定碳氮代谢相关酶活性。参照Yang等[15]的方法测定腺苷二磷酸葡萄糖焦磷酸化酶、蔗糖磷酸合成酶(SPSase)、蔗糖合成酶(SSase)、淀粉合成酶(SSSase)的活性;按照李太贵等[16]的方法测定(SBEase)的活性;参照邹琦[17]的方法,测定硝酸还原酶(NR)、亚硝酸还原酶(NiR)、谷氨酰胺合成酶和NADH谷氨酸脱氢酶(NADH-GDH)的活性。
1.3.4 可溶性总糖、淀粉、游离氨基酸以及可溶性蛋白含量的测定 采用蒽酮比色法[18]测定籽粒的可溶性总糖和淀粉含量,采用茚三酮比色法[17]测定游离氨基酸含量,采用考马斯亮蓝G-250染色法测定蛋白质含量[17]。
1.4 数据统计与分析
用 Excel 2010 对数据进行整理,采用SPSS 20.0 软件进行单因素方差分析,采用 Duncan 检验法进行多重比较及差异显著性分析,利用SigmaPlot 14.0软件绘制图表,数据以平均值±标准偏差表示。
2 结果与分析
2.1 增密对夏玉米弱、强势粒灌浆及灌浆速率的影响
如图1 所示,2 a研究结果均表明,与强势粒相比,种植密度对弱势粒粒质量与灌浆速率影响较大。增密明显降低了灌浆中后期弱势粒的粒质量与灌浆速率,而对强势粒没有明显影响。2020年自授粉后35 d开始,PD3的弱势粒粒质量与PD1和PD2的弱势粒粒质量之间的差异逐渐增加,在授粉后55 d,与
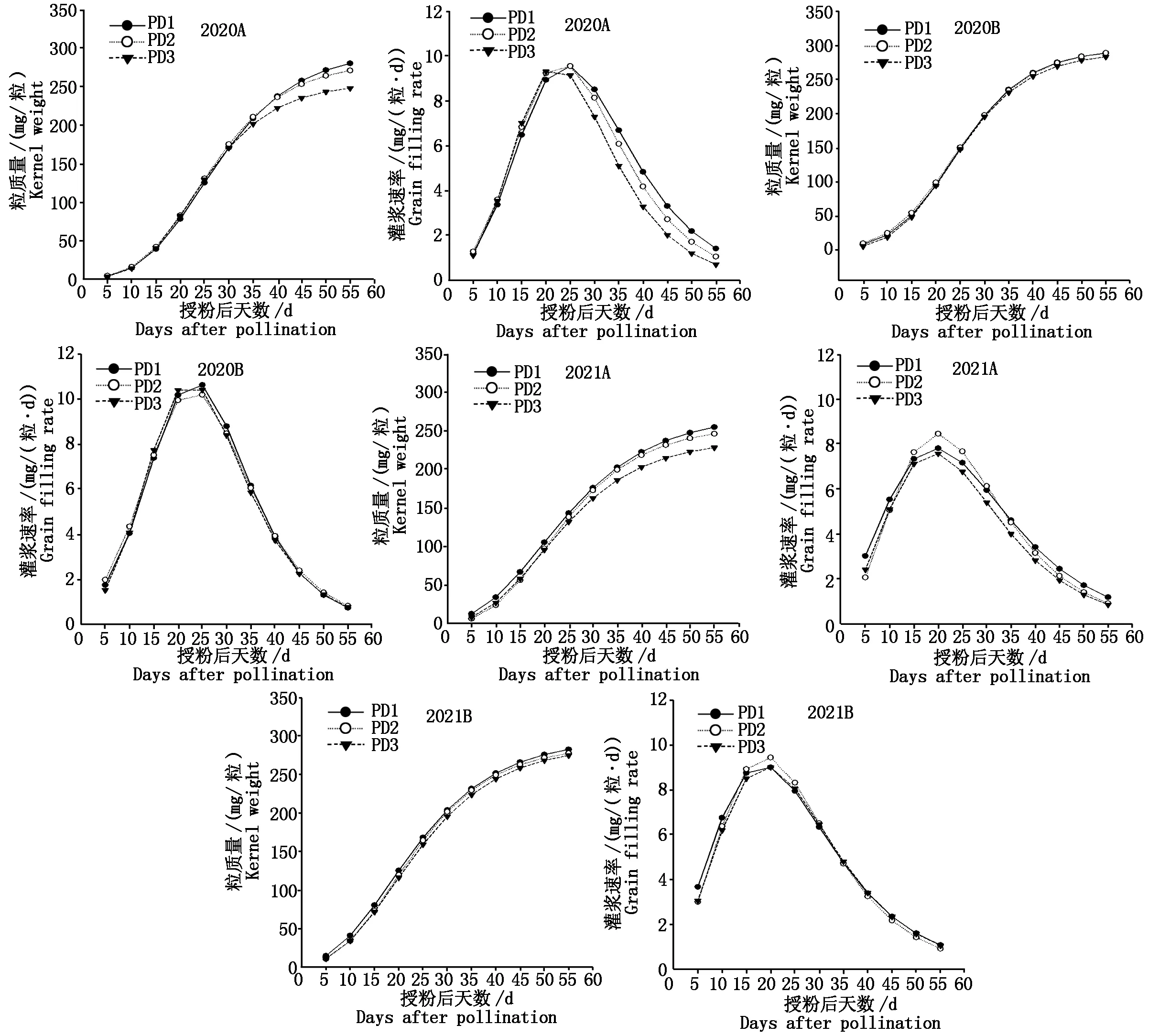
A.弱势粒;B.强势粒。A.Inferior kernel;B.Superior kernel.
PD1相比,PD2与PD3的弱势粒粒质量分别降低3.3%和11.5%。弱势粒的灌浆速率自花后25 d开始在不同密度处理间开始呈现差异,表现为PD1>PD2>PD3,与PD1相比,PD2和PD3的平均灌浆速率分别降低3.7%和11.9%。2021年自授粉后30 d开始,弱势粒粒质量在不同密度处理间开始呈现差异,表现为PD1和PD2显著大于PD3,且随着灌浆进程的推进差异逐渐加剧;与PD1相比,PD2和PD3的弱势粒粒质量分别降低3.4%和10.4%。不同密度处理的最大灌浆速率出现在授粉后20 d,最大灌浆速率表现为PD2>PD1>PD3,而平均灌浆速率表现为PD1>PD2>PD3。
2.2 增加种植密度对弱、强势粒粒质量比的影响
弱、强势粒粒质量比是反映玉米粒位效应的重要指标。由图2可知,增密降低了玉米弱、强势粒粒质量比,但在不同年度间存在差异。2020年,与PD1相比,PD2和PD3的弱、强势粒粒质量比分别显著降低了3.6%和9.7%。2021年,PD1与PD2之间不存在显著差异,但是,与PD1相比,PD3的弱、强势粒粒质量显著降低7.2%。同一密度处理下,2021年的弱、强势粒粒质量比普遍低于2020年。与PD1相比,2020,2021年PD3的弱、强势粒质量比均明显降低, 平均降幅为8.45%。
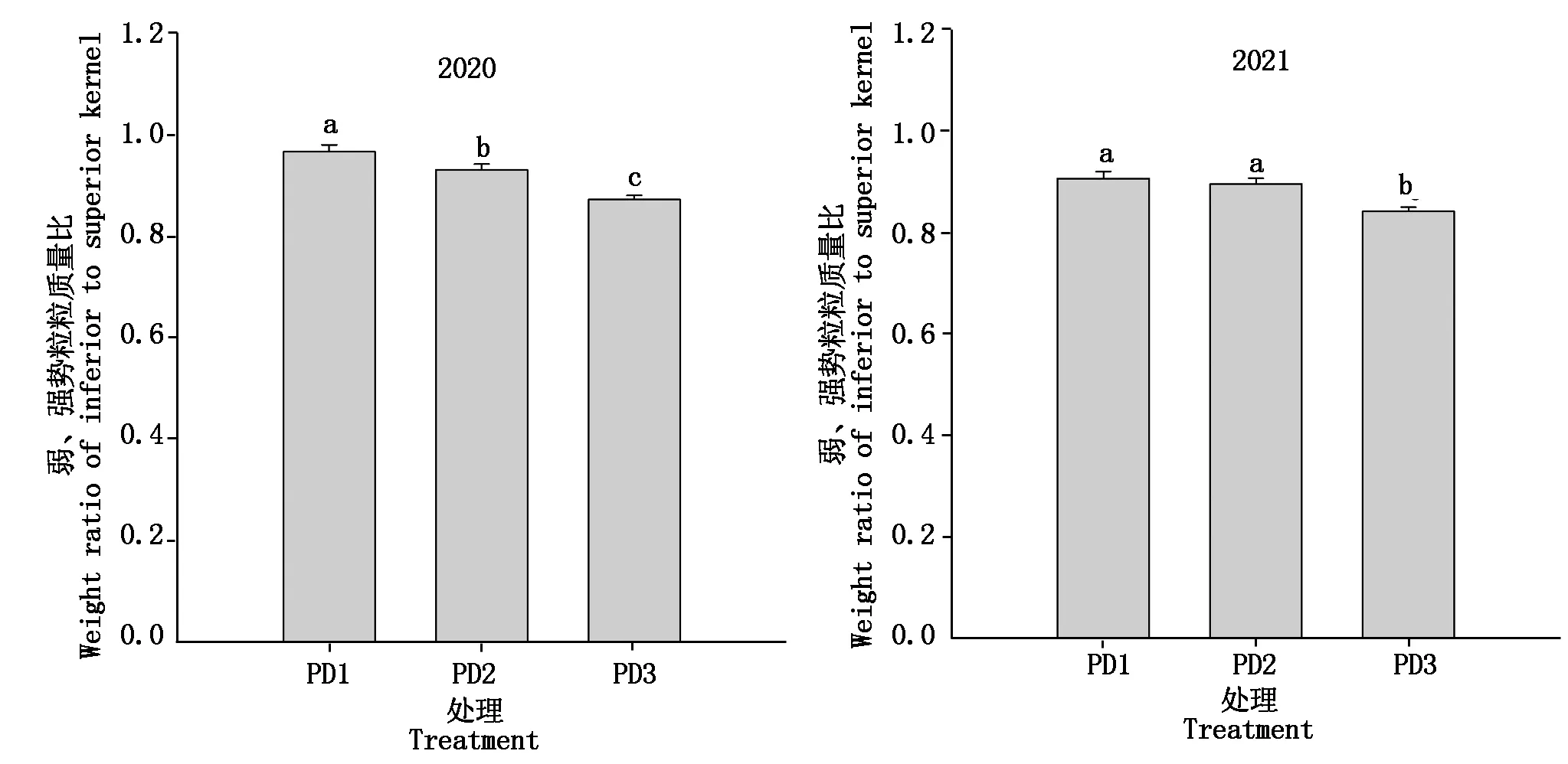
不同小写字母表示差异显著(P<0.05)。图3—4同。Different lowercase letters indicate significant difference at the 0.05 probability level.The same asFig.3—4.
2.3 增密对夏玉米植株干物质积累和收获指数的影响
增密明显影响玉米花前、花后、单株总干物质积累量与收获指数(表1)。 随着种植密度的增加,花前、花后及总干物质积累量均呈降低趋势。与PD1相比,PD2和PD3分别降低花前干物质积累量7.3%和14.0%(2 a平均)、降低花后干物质积累量11.3%和25.3%(2 a平均)、降低总干物质积累量 8.9%和20.9%(2 a平均)。2020年,PD1和PD2之间收获指数不存在显著差异,但PD3的收获指数显著低于PD1和PD2;2021年,与PD1相比,PD2和PD3分别降低收获指数4.6%和6.5%,且差异达到显著水平。与2020年相比,2021年不同密度处理的玉米花前、花后及总干物质积累量普遍低于2020年,但2021年的收获指数均高于2020年。
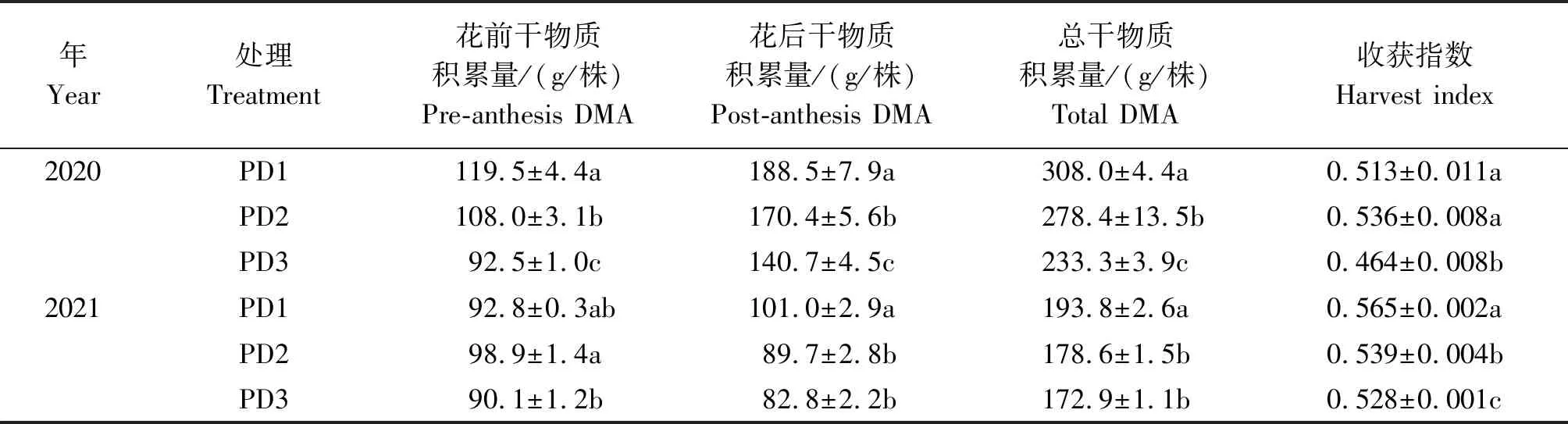
表1 增密对夏玉米单株干物质积累量与收获指数的影响Tab.1 Effect of increasing plant density on plant dry matter accumulation and harvest index of summer maize
2.4 增密对夏玉米弱、强势粒碳代谢酶活性的影响
玉米灌浆中期弱、强势粒碳代谢相关酶活性对增密的响应存在差异(表2)。在相同密度处理下,强势粒的SPSase活性(以鲜质量计)显著高于弱势粒,种植密度对相同粒位籽粒的SPSase活性影响不显著。但与PD1相比, PD2与PD3处理的弱、强势粒之间的差异分别增加了64.0%和67.5%。SSase活性(以鲜质量计)在同一密度不同粒位之间不存在显著差异,除了PD2的弱势粒显著低于PD1和PD3之外,PD2的强势粒显著低于PD3,其余处理之间没有显著差异。同一密度处理下,强势粒ADPGase活性(以鲜质量计)明显高于弱势粒;增密不仅降低了不同粒位籽粒的ADPGase的活性,也加剧了弱、强势粒之间ADPGase的差异。SSSase(以鲜质量计)在同一密度不同粒位之间存在显著差异,与PD1相比,PD3显著降低了不同粒位的SSSase活性。弱、强势粒之间的差异大小则表现为PD3>PD1>PD2。在同一密度处理下,弱势粒的SBEase活性(以鲜质量计)显著低于强势粒,但增密对弱、强势粒的SBEase活性没有显著影响。
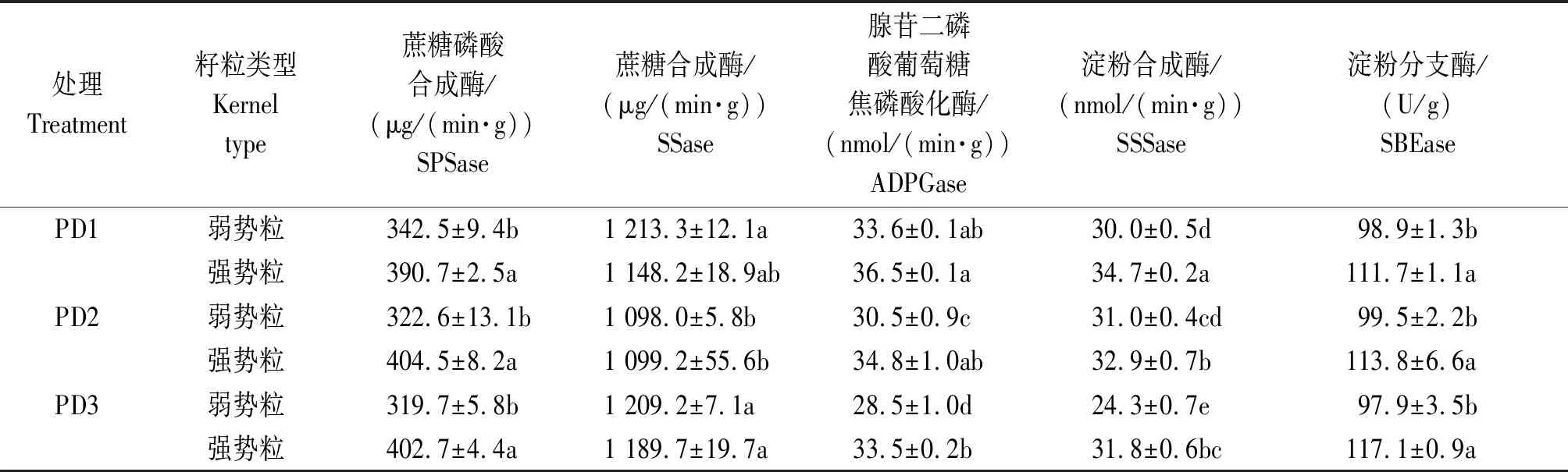
表2 增密对夏玉米弱、强势粒SPSase、SSase、ADGPase、SSSase与淀粉分支酶活性的影响Tab.2 Effect of increasing plant density on SPSase,SSase,ADPGase,SSSase,and SBEase activities of summer maize
2.5 增密对夏玉米弱、强势粒氮代谢酶活性的影响
如表3所示,除了NiR之外,同一密度处理下弱势粒的GS、NR和NADH-GDH的活性(以鲜质量计)均显著低于强势粒,增密对夏玉米弱、强势粒不同氮代谢相关酶活性的影响则存在一定的差异。PD2处理的弱、强势粒GS活性显著高于PD1和PD3处理,NR活性随着种植密度的增加则显著降低,但种植密度对弱、强势粒的NiR(以鲜质量计)和NADH-GDH的活性没有显著影响。此外,随着种植密度的增加,玉米弱、强势粒之间GS活性的差异逐渐增加。
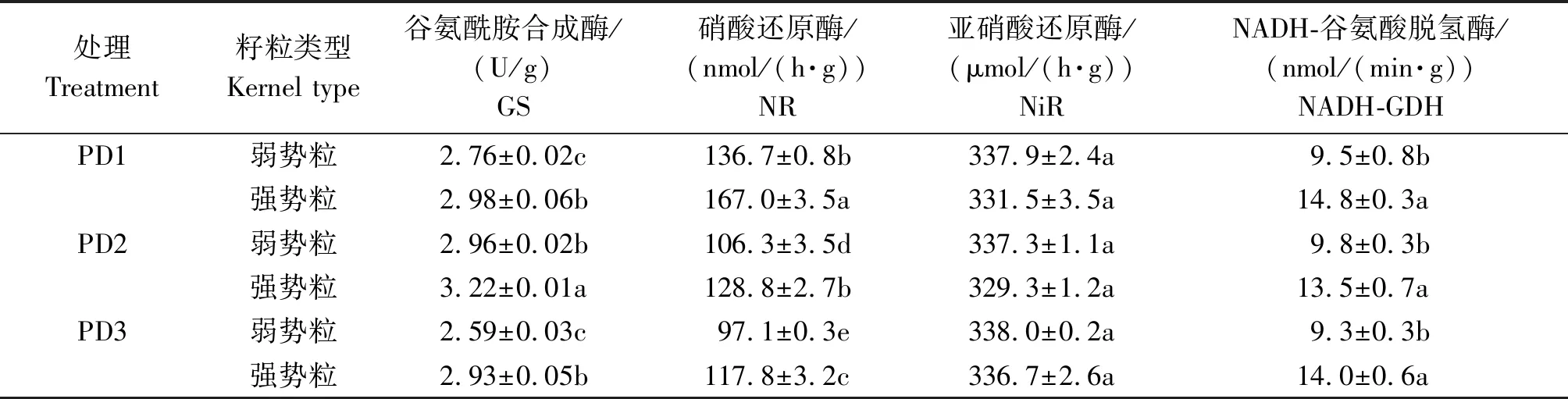
表3 增密对夏玉米弱、强势粒GS、NR、NiR、NADH-GDH活性的影响Tab.3 Effect of increasing plant density on GS,NR,NiR,and NADH-GDH activities of inferior and superior kernels of summer maize
2.6 增密对夏玉米弱、强势粒可溶性糖和淀粉含量的影响
种植密度影响弱、强势粒可溶性糖和淀粉含量(图3)。 随着种植密度的增加,弱势粒的可溶性糖含量逐渐降低。与PD1相比,PD2和PD3降幅分别为10.4%和33.8%,且PD1和PD3之间的差异达到显著水平。强势粒中可溶性糖含量在不同种植密度处理下表现为PD2>PD1>PD3,且相互间差异均达到显著水平。此外,增密加剧了弱、强势粒之间的可溶性糖含量的差异。弱势粒中淀粉含量在PD1和PD2处理之间没有显著差异,但与PD1和PD2相比,PD3显著降低了弱势粒的淀粉含量;强势粒中淀粉含量在不同密度处理之间差异不显著。随着种植密度的增加,弱、强势粒之间淀粉含量的差异也逐渐增加;与PD1相比,PD2与PD3处理的弱、强势粒之间的差异分别增加18.0%和129.7%。
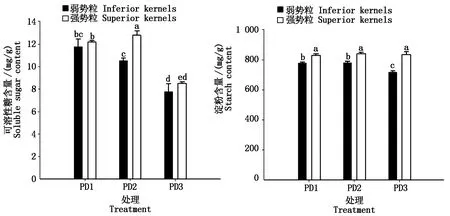
图3 增密对夏玉米弱、强势粒可溶性糖和淀粉含量的影响Fig.3 Effect of increasing plant density on soluble sugar and starch content of inferior and superior kernels of summer maize
2.7 增密对夏玉米弱、强势粒氨基酸和蛋白质含量的影响
如图 4 所示,种植密度对弱、强势粒氨基酸和蛋白质含量以及粒位间的差异均有明显影响。与PD1和PD2相比,PD3显著降低了弱势粒和强势粒的氨基酸含量,但是PD1和PD2之间没有显著差异。 此外,与PD1和PD2相比,PD3明显加剧弱、强势粒之间氨基酸含量的差异。弱、强势粒的蛋白质含量在不同密度处理之间表现为PD2>PD1>PD3,且不同处理之间的差异均达到显著水平。但是,随着种植密度的增加,弱、强势粒之间蛋白质含量的差异呈逐渐增加的趋势。
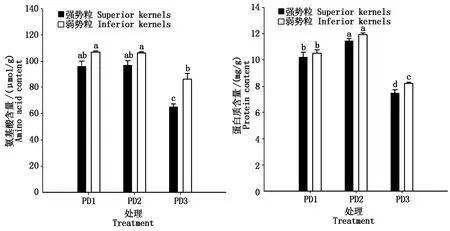
图4 增密对夏玉米弱、强势粒氨基酸和蛋白质含量的影响Fig.4 Effect of increasing plant density on amino acid and protein content of inferior and superior kernels of summer maize
3 结论与讨论
3.1 玉米弱、强势粒灌浆与粒位效应对增密的响应
在适宜的水肥等栽培管理条件下,合理增加种植密度是提高玉米产量的重要途径和必然趋势[19-21]。但是,过高的种植密度会加剧作物群体体内个体之间对光照、养分和水分的竞争,进而影响单株穗粒数和粒质量[22]。在较高的种植密度条件下,当穗粒数确定后,粒质量则成为决定产量的重要因子[11]。已有研究结果证实,增密影响籽粒的灌浆进程和粒质量[13],这与密植诱发玉米花后植株早衰存在密切关系[23],因为花后早衰不仅降低了同化物向籽粒中的分配,还缩短了籽粒的灌浆活跃期,最终影响籽粒的灌浆充实[24-25]。本研究结果显示,增密同时降低了玉米弱、强势粒平均灌浆速率和粒质量,但对弱势粒的影响较为明显,表明增密主要是影响夏玉米弱势粒的灌浆充实,这与王晓燕等[6]研究结果相似。相关研究证实,玉米果穗上部弱势粒相对于强势粒更易受到非生物逆境胁迫的影响[26]。增密通过降低冠层光分布导致弱光胁迫可能是显著影响弱势粒灌浆充实的主要因素,因为前人通过遮光试验也表明,弱光胁迫会明显影响玉米弱势粒发育甚至导致其败育[8]。关于种植密度对弱势粒灌浆影响的起始时间,前人研究认为,起始于灌浆前期,但是本研究发现,密植明显影响弱势粒粒质量的时间是自授粉后30~35 d开始,说明弱势粒受密植影响主要是从灌浆中期开始。
玉米籽粒发育普遍存在着粒位效应[5,27],而玉米弱、强势粒粒质量比是反映玉米粒位效应的一个重要指标,其值越小,粒位效应越明显。本研究结果显示,增密可显著降低弱、强势粒的粒质量比,表明增密加剧了玉米的粒位效应,这主要与密植明显降低弱势粒灌浆速率和粒质量有关。此外,密植明显降低了玉米植株花后干物质积累量,而较高的花后物质积累量是籽粒灌浆充实和高产的重要保证[28],增密影响弱势粒灌浆和粒质量可能与植株花后同化物积累显著降低有关。前人研究证实,玉米灌浆期光合同化物在果穗籽粒中的分配具有明显的粒位效应,顶部弱势粒分配到的同化物低于果穗中下部强势粒[5],通过不当的栽培措施或受逆境胁迫减少同化物的供应时,均能加剧玉米强、弱势粒之间的差异[9,26,29]。因此,种植密度的增加导致花后同化物供应受限时,光合同化物可能优先供应强势粒,弱势粒因得不到足够的同化物供应导致粒质量明显降低,进而加剧玉米的粒位效应。
3.2 玉米弱、强势粒碳氮代谢特征对增密的响应
玉米籽粒灌浆充实的过程,主要是淀粉合成与积累的过程[5]。光合同化物以蔗糖的形式从源输送到籽粒,再经一系列酶促反应形成淀粉[30]。在这一过程中,由源供应的碳水化合物和淀粉合成相关酶起着重要作用。本研究结果显示,在密植条件下,弱势粒中可溶性糖、淀粉含量及蔗糖-淀粉合成相关酶(SPSase、ADPGase、SSSase、SBEase)活性在多数情况下明显低于强势粒,这与徐云姬等[5]研究结果相似,均表明玉米弱势粒较弱的碳代谢水平是其灌浆差、粒质量低的重要原因。此外,本研究结果表明,随着种植密度的增加,玉米强、弱势粒间淀粉含量、SPSase与ADPGase活性差异也逐渐增加。因此,增密加剧玉米粒位差异与其引起强、弱势粒间SPSase与ADPGase活性进一步差异化相关,因为SPSase与ADPGase是作为淀粉合成途径中的关键酶和限速酶,其活性大小对籽粒淀粉合成至关重要[31-32]。
籽粒中氮代谢在籽粒的发育与建成上同样发挥着重要作用[10],但目前关于种植密度对玉米不同粒位籽粒氮代谢生理的研究却较少。本研究显示,在相同的密度处理下弱势粒中氮代谢相关酶(GS、NR和NADH-GDH)活性、游离氨基酸和蛋白质含量均低于强势粒,表明弱势粒的氮代谢水平同样低于强势粒。与中低密度相比,高密度不仅同时降低了弱、强势籽粒的含氮化合物的含量,还加剧了强、弱势粒间含氮化合物的差异,这可能与密植加剧玉米强、弱势粒间GS活性的差异存在密切关系,因为GS是参与籽粒代谢的关键酶,其活性高低直接关系氨基酸供体的合成与转化,进而影响籽粒中蛋白的合成[33]。 因此,增密加剧玉米粒位差异与其改变不同粒位间GS活性可能存在一定关系。
综上所述,增密加剧了夏玉米的粒位效应,这与其明显影响玉米弱势粒灌浆充实有关。增密限制玉米弱势粒灌浆充实主要体现在以下两方面:一是增密降低了花后干物质积累量,不利于为弱势粒灌浆提供充足的物质保证;二是增密明显抑制弱势粒中SPSase、ADPGase、GS活性,进而影响籽粒中淀粉和蛋白的合成与积累。
