花期低温对甘蓝型油菜叶片生理及雌雄蕊发育的影响
2022-11-03李浩东秦梦凡王茸茸朱云林徐爱遐
李浩东,秦梦凡,张 淼,王茸茸,徐 宇,宋 葭,朱云林,黄 镇,徐爱遐
(西北农林科技大学 农学院,陕西 杨凌 712100)
倒春寒是指在春季之后(3,4月份)气温回升,随后气温大幅度下降,甚至到0 ℃以下的天气现象。近年来,春季低温(倒春寒)时有发生,而甘蓝型冬油菜的花期从北向南依次发生在春季的3,4月份,倒春寒发生时冬油菜正处于生殖生长的旺盛阶段,遭受倒春寒低温影响后,油菜的生长发育尤其是幼嫩的生殖器官受冻严重,影响授粉结实和角果发育,有效结角率、每角粒数和种子含油量下降,直接影响到产量和产油量[1-2]。2018,2021年在黄淮和长江下游发生大面积的倒春寒,导致较多不耐春寒的油菜品种结实受到严重影响,产量损失严重。因此,研究油菜花期对低温胁迫的响应,对于深入研究油菜花期寒害机理,筛选和创制花期耐寒的油菜种质,培育耐春寒的冬油菜品种具有理论和现实意义。
油菜苗期对冬季寒冷响应的相关研究较多,蒲媛媛等[3]测定白菜型冬油菜陇油6号和天油4号在越冬期的一系列生理指标,发现耐寒性强的陇油六号在低温条件下有着较高的SOD、CAT、SP含量与较低的MDA含量,因此,认为这些指标在一定程度上可以反映出白菜型冬油菜的抗寒性强弱。朱惠霞等[4]通过在越冬期测定8个白菜型冬油菜品种的SOD、POD、Pro、SS、SP,发现越冬率高的品种酶活性和可溶性物质含量均比越冬率低的品种高。在对温敏性不同的油菜低温胁迫后发现,冬性品种的SOD、POD、CAT、APX活性强于春性品种[5-6]。闫蕾等[7]研究发现,在低温环境下,抗寒性强的材料其生物量、叶绿素和脯氨酸的累积量都高于敏感材料,而相对电导率无显著差异。李世成等[8]对油菜进行零上低温胁迫(20,15,10,5 ℃),发现丙二醛、可溶性糖和游离脯氨酸含量上升。但油菜花期受低温胁迫的相关研究较少,Larden等[9]通过对油菜低温胁迫((-3±0.5)℃,4 h),发现植株出现主花序弯曲,长3 mm的花蕾开花后的花粉活力丧失80%,并且随着种子的发育,抵抗低温的能力越强。涂玉琴等[1]在田间低温阴雨条件下对19个油菜品种进行分析,发现有效结角率、每角粒数、种子含油量均会降低,而千粒质量则增加。作物在生殖生长阶段遭受低温胁迫的研究大多集中于水稻和小麦,刘红杰等[10]通过研究冬小麦田间气温变化规律,发现在不同高度出现0 ℃以下温度的时间、持续时间以及最低气温都不相同,表现在小麦冠层处气温更低、持续时间更长。高芸等[11]通过对小麦倒二叶出生期、孕穗期和开花期进行低温处理,发现处理后倒二叶出生期花粉败育率和穗位数降低的幅度最大。Zhang等[12]研究表明,对冬小麦孕穗期低温处理,会引起脱落酸积累,改变蔗糖代谢酶的活性,导致蔗糖在幼穗积累从而对小麦生长造成影响。张献国等[13]研究表明,水稻在孕穗期遭受低温胁迫后,水稻倒二叶SOD、CAT、POD的活性有较大变化。任红茹[14]在水稻孕穗期时施加低温胁迫(13,15,17 ℃)发现,剑叶叶片中SOD、POD、CAT的活性随着低温处理时间或温度的降低不断升高。
本试验以2个冬季寒冷抗性不同的甘蓝型油菜品系为材料,研究花期低温胁迫后叶片和生殖器官对低温的响应,探究花期低温对油菜生长发育的影响,旨在为今后深入研究油菜花期耐低温机理奠定理论基础。
1 材料和方法
1.1 试验材料
选取2份甘蓝型油菜材料:GZ恢抗冬季(苗期)寒冷强、10B抗冬季(苗期)寒冷弱,材料均来自西北农林科技大学农学院油菜资源课题组。
1.2 试验方法
GZ恢和10B于2020年9月种植于西北农林科技大学试验农场,田间生长通过春化(10~12叶期),于当年12月10日挖取长势一致的植株盆栽,置于温室(24 ℃/14 h,16 ℃/10 h)继续生长,每个材料25盆(株),其中5盆(株)作对照,20盆(株)作低温处理。当植株生长至有3~5朵花开放时,对每个单株主要花序的花蕾依据大小进行标记,同时取不同大小花蕾进行细胞学观察确定花蕾发育时期,之后将要处理的植株(每个材料各20株)放入人工气候室(12 ℃/14 h,2 ℃/10 h),设置5个不同时长的低迫处理:1,2,3,4,5 d,每处理4株。各处理完成后,立即在处理完的4个单株上各取上部苔茎叶1 g和下部短柄叶1 g,苔茎叶和短柄叶分别混样,立即放入液氮速冻,后存放-80 ℃冰箱用以测定胁迫后生理指标,每个处理取样3次重复。未胁迫的对照单株同样方法取样测定生理指标。取样后的植株移回温室(24 ℃/14 h,16 ℃/10 h)恢复生长,观察花蕾的生长发育,待标记的各级花蕾开放后对其花粉活力和雌蕊柱头接受花粉的能力(可授性)进行测定。对照植株一直在温室(24 ℃/14 h,16 ℃/10 h)生长,同样在各级花蕾开放后对其花粉活力和雌蕊柱头的可授性进行测定。
1.2.1 生理指标的测定 可溶性蛋白(SP)、可溶性糖(SS)、丙二醛(MDA)、超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性的测定方法参照植物生理学实验指导[15]。
1.2.2 花粉活力及柱头可授性测定 低温不同处理后植株移至与对照相同的正常环境生长,待不同大小花蕾生长至开花,根据处理前标注的花蕾大小,从3个单株上各取3个当天开放(9:00—10:00)花朵的花粉和相应柱头,花粉取样后立即用醋酸洋红法[16]观察不同大小(发育阶段)花蕾受低温胁迫后的花粉活力,在荧光显微镜(ZEISSImager M2)下观察花粉染色情况,若花粉被染成深红色则具有活性,若被染成浅红色或未被染色则为无活性,每种长度的花蕾设3个重复,每个重复观察3个视野,最后计算平均值。花粉活力=(被染成深红色的花粉粒数目/总花粉粒数)×100%;柱头采用联苯胺-过氧化氢法[17]测定不同大小(发育阶段)花蕾受低温胁迫后的柱头可授性,将采集的柱头放在载玻片上,在柱头周围滴落联苯胺-过氧化氢反应液,若柱头周围有大量气泡产生且变为蓝色,则具有极强可授性(+++),若有气泡且变为蓝色则具有中等可授性(++),若有少量气泡产生且不变色则具有可授性(+)。对照为始终处于正常环境的同一级别花蕾的花粉和柱头。
1.3 数据处理
利用IBM SPSS Statistics 25进行数据处理,利用GraphPad Prism 7进行绘图。
2 结果与分析
2.1 不同时长低温处理对油菜生长发育的影响
2.1.1 植株形态和花蕾发育 图1是GZ恢和10B在花期遭受不同天数低温胁迫后的表型变化,由图1可见,不同时长低温胁迫后,植株叶片表现有萎蔫,随着处理时间的延长,叶片萎蔫的程度有所加重,但总体来看,即便是处理最长时间5 d,GZ恢和10B叶片萎蔫均不严重;低温胁迫后在正常环境中恢复生长一段时间后,与没有处理的CK相比,各处理后植株的生长发育受到一些影响,突出表现为处理3 d以上,花序顶端小花蕾(小于3.0 mm)不同程度死亡脱落,处理时间越长,死亡花蕾数越多,处理4,5 d 小于3.0 mm发育阶段的花蕾全部死亡。同时观察到处理5 d的部分大花蕾(大于3.0 mm)尽管开花,但随后花柄脱落,低温处理1,2 d花蕾生长发育从外观看不出明显差异,也无死蕾现象。由此可见,12 ℃/14 h,2 ℃/10 h胁迫超过4 d会对油菜小于3.0 mm的花蕾造成致死性伤害。会使油菜植株部分大于3.0 mm的花蕾开花后花柄脱落。

图1 低温处理后植株形态变化Fig.1 Plant morphology changes after cold stress
2.1.2 叶片生理指标的变化 GZ恢和10B各处理的SOD活性(以鲜质量计)如图2所示,GZ恢苔茎叶和下部叶SOD活性各处理与CK无显著差异。10B苔茎叶在不同时间低温胁迫处理后,其SOD活性均比对照高,其中处理4 d后比CK增高了41.76%,而下部叶各处理与CK并无显著差异;从两部位叶片来看,除GZ恢处理1 d外,两材料苔茎叶SOD活性CK与不同处理均高于下部叶。从两材料对比来看,GZ恢SOD活性不论是CK还是处理均比10B要高。
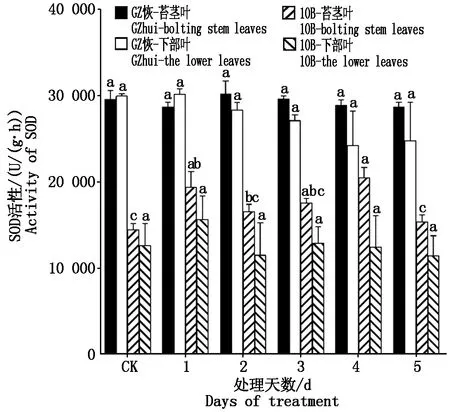
不同字母表示同一材料间同一部位不同处理下差异显著(P< 0.05)。图3—7同。Different letters indicate significant differences between same materials under the different treatment(P<0.05).The same asFig.3—7.
GZ恢和10B各处理的POD活性(以鲜质量计)见图3。GZ恢苔茎叶POD活性在处理后均比CK高,其中处理第1天与第3天涨幅较大,较CK相比分别达到了85.53%,113.24%。
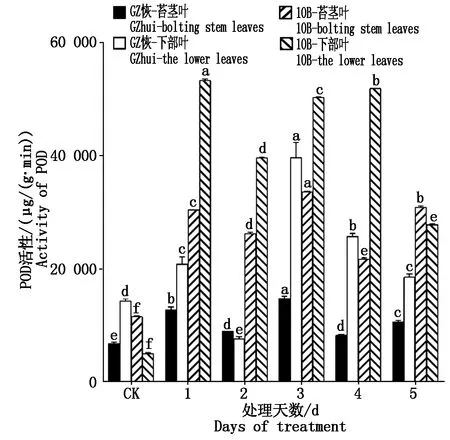
图3 低温胁迫后POD活性的变化Fig.3 Changes of peroxidase activity after low-temperature stress
GZ恢下部叶处理第2天的POD活性比CK低,降低了45.60%,其余处理均比CK高,处理第3天POD活性最高,较CK增加了1.74倍;从两部位叶片来看,除处理2 d GZ恢下部叶POD活性低于苔茎叶,其余处理与CK均是下部叶POD活性高于苔茎叶。10B两部位叶POD活性所有处理都显著高于CK(P<0.05),其中苔茎叶POD活性最高值出现在处理第3天,而下部叶最高值出现在处理第1天,较CK分别增加了1.86,9.35倍;从两部位叶片来看,除CK与处理5 d后,其余处理10B下部叶POD活性均高于苔茎叶。从两材料来看,GZ恢苔茎叶CK与处理POD含量低于10B苔茎叶,而GZ恢下部叶CK为POD含量高于10B,处理后却表现为10B下部叶POD含量高于GZ恢。
GZ恢和10B各处理的CAT活性如图4所示,GZ恢苔茎叶低温胁迫后除处理2 d的CAT活性高于CK、处理5 d后活性显著低于CK(P<0.05),其余处理与CK比并无显著差异,其中处理第5天较CK显著降低约29.16%;GZ恢下部叶处理3 d较CK下降了30.38%,其余处理较CK来说均无显著差异;从两部位叶片来看,GZ恢在处理2,3 d后苔茎叶CAT活性高于下部叶,CK与其余处理则呈现下部叶高于苔茎叶。10B苔茎叶处理1,2,3 d后CAT活性均低于CK,且处理3 d后与CK比显著降低(P<0.05),降低幅度达86.22%,在处理4 d后与对照相比有显著升高(P<0.05),升高的幅度为4.20%,处理5 d后与CK无显著差异;而其下部叶处理后CAT活性均低于CK,处理2 d后降低幅度最大,为85.97%,但处理5 d后与CK相比并无显著差异;从两部位叶片来看,10B在处理2,4 d后CAT活性下部叶低于苔茎叶,CK与其余处理均为下部叶高于苔茎叶。从2个材料来看,不论CK还是处理,GZ恢两部位叶片的CAT活性都远远高于10B。

图4 低温胁迫后CAT活性的变化Fig.4 Changes of catalase activity after low-temperature stress
GZ恢和10B各处理的MDA含量如图5所示,GZ恢苔茎叶在不同时长低温处理后,MDA含量均与CK无显著差异;而GZ恢下部叶低温处理后其MDA含量均比CK高,其中处理4 d达到最高,较CK比升高了2.53倍;且GZ恢下部叶MDA含量高于苔茎叶。10B苔茎叶处理第1,2天后MDA含量与CK无显著差异,在处理3,4,5 d后较CK显著升高(P<0.05),在处理第5天达到最高峰,较CK相比升高了2.04倍;下部叶在处理后MDA含量较CK相比升高,在第2天达到最高,较CK增长了13.87倍;从两部位叶片来看,除处理1,2 d后下部叶MDA含量高于苔茎叶,CK与其余处理均为苔茎叶高于下部叶。从两材料来看,GZ恢两部位叶片的MDA含量远远低于10B。
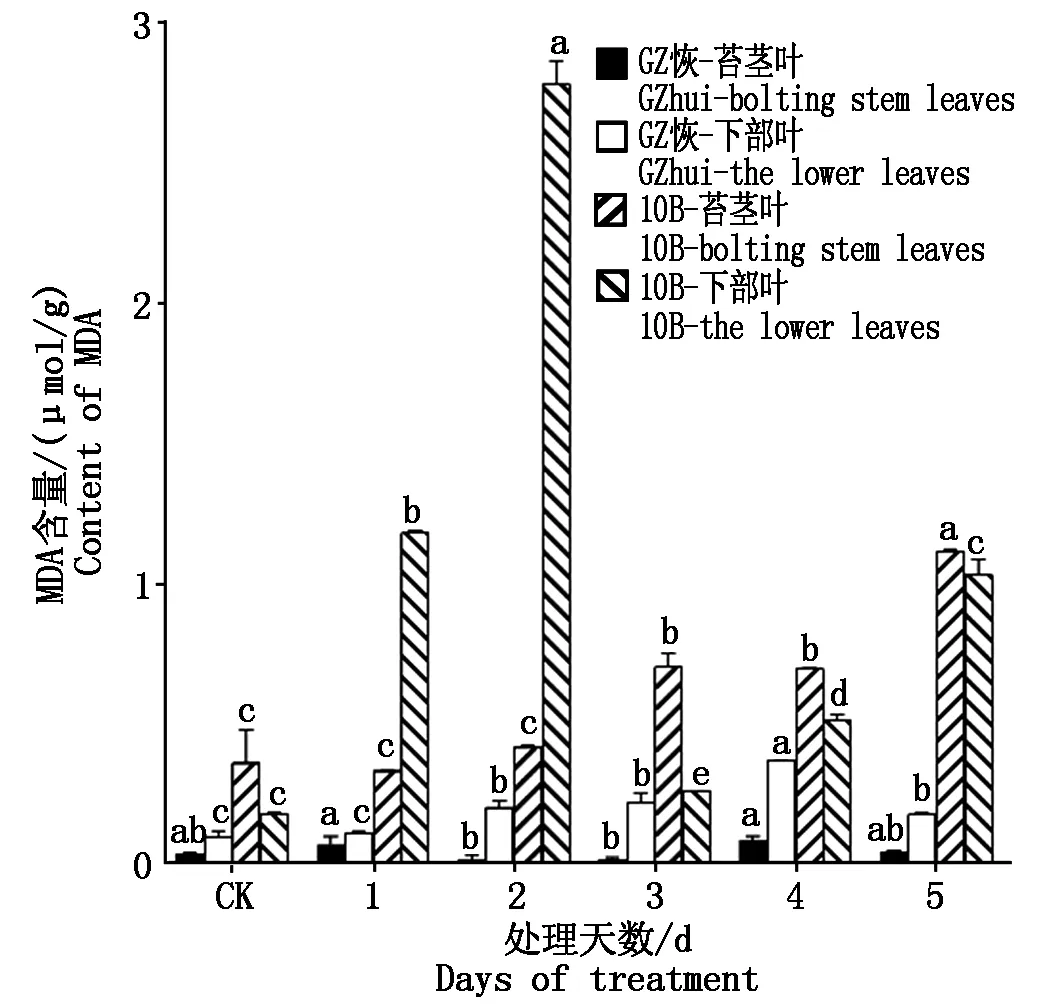
图5 低温胁迫后MDA含量的变化Fig.5 Changes of malondialdehyde content after low-temperature stress
GZ恢和10B各处理的SS含量如图6所示,GZ恢苔茎叶低温胁迫后所有处理SS含量均显著高于CK,其中处理5 d后含量最高,较CK增加了2.77倍,下部叶也同样所有处理均显著高于CK(P<0.05),但含量最高值处于处理2 d后,较CK增加了3.62倍;从两部位叶片来看,下部叶的SS含量高于苔茎叶。10B苔茎叶处理1,2,3 d后SS含量显著低于CK(P<0.05),处理2 d后最低,较CK降低了36.65%,处理4 d后较CK显著升高了38.12%(P<0.05),处理5 d后与CK比较并无显著差异;其下部叶与苔茎叶变化基本相同;从两部位叶片来看,10B下部叶的SS含量高于苔茎叶。从两材料对比来看,GZ恢的SS含量CK与处理均高于10B。

图6 低温胁迫后SS含量的变化Fig.6 Changes of soluble sugar content after low-temperature stress
GZ恢和10B各处理的SP含量如图7所示,GZ恢低温处理后苔茎叶SP含量与CK均无显著差异,下部叶与苔茎叶表现相同;从两部位叶片来看,其SP含量差异不显著。10B苔茎叶低温处理后同样所有处理SP含量与CK比无显著差异;10B下部叶处理后SP含量也同样与CK无显著差异;从两部位来看,10B苔茎叶SP含量不论CK还是处理,均高于下部叶。从两材料来看,10B CK和处理的SP含量均高于GZ恢。

图7 低温胁迫后SP含量的变化Fig.7 Changes of soluble protein content after low-temperature stress
2.2 不同时长低温胁迫对花蕾发育及花粉活力的影响
对分别经过12 ℃/14 h,2 ℃/10 h 处理1,2,3,4,5 d的GZ恢和10B的不同发育阶段的花蕾的发育及花粉活力进行测定。从表1可以看出,GZ恢和10B在没有低温胁迫时,各级大小花蕾开花后的花粉活力均在92%以上,平均分别为95.64%,93.58%;处理1 d后,GZ恢小于1.5 mm的花蕾花粉活力为80.82%,与CK(95.31%)比较花粉活力显著下降(P<0.05),大于1.5 mm的花蕾开花后花粉活力均与CK无显著差异。处理2,3 d后,GZ恢小于3.0 mm的花蕾花粉活力与CK比较显著下降(P<0.05),活力在72.83%~90.51%,均值为79.22%,而大于3.0 mm的花蕾花粉活力几乎不受影响。处理4,5 d后,大于3.0 mm的花蕾仍然表现花粉活力几乎不受影响,但小于3.0 mm的花蕾均表现死亡,不能正常开花。10B受低温胁迫花蕾受冻和花粉活力与GZ恢具有同样的趋势:花蕾越小受害越严重,花粉活力越低。具体表现与GZ恢相同的是:小于3.0 mm的花蕾处理4,5 d后均表现死亡;不同的是GZ恢大于3 mm花蕾在经过各级时长低温处理后,其花粉活力较CK均无显著差异,但10B仅大于6.0 mm的花蕾经过1,2,3,4 d的处理后花粉活力无显著降低,且10B 3.0~6.0 mm的花蕾所有处理花粉活力均有显著降低(P<0.05)。总体来看,10B受不同时长的低温胁迫后,相同大小花蕾的花粉活力多低于GZ恢。纵观2个材料可以看出,花期低温(12 ℃/14 h,2 ℃/10 h)超过4 d,小于3.0 mm的花蕾发育停止、花瓣不能开张直至死亡;大于6.0 mm的花蕾,花期低温(12 ℃/14 h,2 ℃/10 h)4 d以内,其花蕾发育和花粉活力基本不受影响。3.0~6.0 mm的花蕾,花期低温处理后花粉活力差异可很好地反映不同材料耐低温的能力。
2.3 花期低温对油菜柱头可授性的影响
对分别经过12 ℃/14 h,2 ℃/10 h 处理1,2,3,4,5 d的GZ恢和10B的不同发育阶段的花蕾开花后柱头可授性进行测定。从表2可以看出,GZ恢和10B没有低温胁迫时,各级大小花蕾开花后的柱头可授性均具有极强可授性;处理1 d后,GZ恢小于1.5 mm的花蕾柱头可授性为具有可授性,大于1.5 mm小于3.0 mm的花蕾柱头可授性为中等可授性,而大于3.0 mm的花蕾可授性与CK比较并无差异。处理2 d后,GZ恢小于2.0 mm的花蕾为仅具有可授性,2.0~3.0 mm的花蕾为具有中等可授性,大于3.0 mm的花蕾柱头可授性与CK并无差异。处理3 d后,GZ恢小于2.0 mm的花蕾柱头可授性都为仅具有可授性,2.0~6.0 mm的花蕾为中等可授性,大于6.0 mm的花蕾与CK无差异。处理4,5 d后,大于3.0 mm的花蕾均为中等可授性,而小于3.0 mm的花蕾均表现死亡,不能开花。10B受低温胁迫后柱头可授性与GZ恢具有同样的趋势:花蕾越小受害越严重,且10B在处理1 d后,1.5~2.0 mm的花蕾为仅具有可授性。纵观2个材料发现,小于3.0 mm的花蕾,处理1 d后柱头可授性就会有所降低;大于3.0 mm的花蕾,花期低温(12 ℃/14 h,2 ℃/10 h)3 d以内,其柱头可授性基本不受影响,在处理4~5 d后,其柱头可授性有所降低。

表1 低温胁迫对GZ恢、10B花粉活力的影响Tab.1 The changes of pollen vitality in GZhui and 10B after low-temperature stress %

表2 低温处理对GZ恢、10B柱头可授性的影响Tab.2 The changes of stigma receptivity in GZhui and 10B after low-temperature stress
3 结论与讨论

MDA的含量高低则与植物抵御逆境的能力呈现负相关,过量的MDA会使细胞代谢失调,从而导致植物更易受到逆境伤害[19],GZ恢在低温胁迫后MDA的含量低于10B,说明GZ恢细胞受到的伤害低于10B。SS可以提高细胞的保水能力,降低水势,从而保护植株,SP具有很强的亲水性,使得植株可以锁住更多的水分,以此减少植株遭受低温胁迫的伤害。本研究在低温胁迫后,GZ恢的SS含量高于10B,而SP含量则低于10B,说明在遭受低温胁迫后,GZ恢和10B可能通过不同的渗透胁迫物质来抵抗低温造成的伤害。由此可见,植株在遭受低温胁迫后的抵抗机制是非常复杂的,很难由单一指标来判断低温胁迫对植株造成的影响。
开花植物的花粉粒是通过减数分裂产生的,在大多数研究中指出,小配子形成过程中遭受胁迫会导致小孢子无法正常发育至成熟或雄性不育[20-22]。张自阳等[23]利用抗倒春寒材料和不抗倒春寒材料,通过在小麦雌雄穗原基分化期、药隔分化期以及四分体时期0 ℃低温处理72 h后,发现在雌雄穗原基分化期抗倒春寒材料(矮抗58)和不抗倒春寒材料(郑麦366)的穗位数减少幅度最大,分别较CK降低了22.37%,64.71%,因此,证明雌雄蕊原基分化期对低温最为敏感。Satake[24]通过形态学和组织学观察发现,水稻花粉发育处于四分体向单核阶段这一时间对低温胁迫非常敏感。Yu等[25]利用甘蓝型油菜温敏不育系材料进行研究,发现花药的败育发生在四分体至单核小孢子阶段,并且液泡绒毡层的提前破裂与单核小孢子的原生质体降解有关。本研究从GZ恢和10B低温胁迫后花粉活力的测定结果可以看出,小于3.0 mm的花蕾,花期低温(12 ℃/14 h,2 ℃/10 h)超过4 d,花蕾发育停止、花瓣不能张开直至死亡,这一大小的花蕾雄性生殖细胞处于双核期之前,说明双核期前较双核期之后对低温更为敏感;大于6.0 mm的花蕾,花期低温(12 ℃/14 h,2 ℃/10 h)4 d以内,其花蕾发育和花粉活力基本不受影响。
侯国佐[26]研究发现,柱头可授性在日均温10~20 ℃时比日均温10 ℃以下强。Larden等[9]用受低温处理的花粉授未受胁迫柱头,发现对产量有较小影响,并且在受精后20 d是种子发育过程中对低温敏感的时间,而低温胁迫发生在受精后35 d,种子的发育不会受到影响。Kovaleski等[27]发现,通过-6 ℃的低温胁迫会导致未经低温驯化的油菜植株死亡,并且低温会造成降低油菜植株的总干物质、每株角果数以及胚胎活力,并且不论植株是否经过驯化,都不会减小低温对于胚珠的伤害。本研究表明,小于3 mm的花蕾,处理1 d后柱头可授性就会有所降低;大于3.0 mm的花蕾,花期低温(12 ℃/14 h,2 ℃/10 h)3 d以内,其柱头可授性基本不受影响,在处理4~5 d后,其柱头可授性有所降低,这点与前人所研究的低温会造成可授性下降相同。结合上述结果,由于花粉活力降低幅度并不大,可能油菜在花期遭受低温后不结实是由于柱头可授性降低和花粉活力降低共同造成的。常志坤等[28]发现,越冬期日最低气温<-5 ℃的总天数与蕾薹期连续日最低气温<0 ℃天气的最长持续时间,对冬油菜产量影响较为显著。因此,后续会测定花期低温对结实率的影响,以进一步确定花期低温胁迫对甘蓝型油菜的影响。
本研究通过对2个冬季抗寒性不同的甘蓝型油菜材料在花期进行低温胁迫(白天14 h/12 ℃,夜晚10 h/2 ℃;处理时间1,2,3,4,5 d),发现两材料叶片在低温处理后叶片均表现为轻微萎蔫,且植株形态无明显受损,低温胁迫超过3 d会对小于3.0 mm的花蕾造成致死性伤害。低温处理后叶片的生理指标变化较为复杂,其中两材料POD活性处理后敏感,除GZ恢下部叶处理2 d外,其余处理较CK均显著增多,且多数处理呈现下部叶增加幅度比苔茎叶更大,10B较GZ恢升高更多。MDA含量GZ恢远低于10B。渗透调节物质SS含量变化明显,GZ恢远高于10B,因此,生理指标一定程度上反映了GZ恢比10B更抗花期低温。两材料低温胁迫后,大于6.0 mm的花蕾花粉活力几乎不受影响,而小于3.0 mm的花蕾在处理1~3 d后花粉活力显著下降,处理4~5 d则死亡,GZ恢3.0~6.0 mm的花蕾在低温胁迫后花粉活力大多高于10B。且两材料柱头可授性表现与花粉活力基本一致。
