南海中部海盆自由生活线虫三个新纪录种描述
2022-10-29郝映东贾素素
陆 洋,郝映东,贾素素,黄 勇,3
(1.聊城大学 生命科学学院,山东 聊城 252059; 2.中国海洋大学 海洋生命学院,山东 青岛 266003;3.生态环境保育与生物资源创新利用山东高校重点实验室,山东 聊城 252059)
0 引言
自由生活海洋线虫(以下简称海洋线虫)是海洋沉积环境中数量最为丰富,分布较为广泛,物种多样性较高的一类小型底栖无脊椎动物类群,其丰度占小型底栖生物总量的70%以上,生物量占60%以上[1,2]。在有机质较丰富的软底沉积物中每平方米丰度可达上百万条。在底栖食物网方面,海洋线虫能够占据不同的营养级,起着承上启下的作用,不仅可以作为大型底栖生物以及一些经济鱼虾类的优质饵料,也可以刺激微生物的生产,加速有机质的降解;在海洋环境监测方面,其种群分布与底栖环境密切相关,其多样性指数和群落分布格局的变化可以作为环境监测的有效工具[3]。并且,海洋线虫有着极强的环境耐受力,其特有的生理生态特点与生殖对策可以较好的适应极端环境,从极易受到扰动的海岸浅滩到极寒缺氧条件下的海底沉积环境(潮下带)都有分布,成为国际底栖生态学家重点研究的对象之一。分类与生物多样性研究是对海洋线虫深入研究的基础,但由于我国相关研究起步晚,人才缺乏等原因,对于该类群的分类和多样性研究还比较薄弱,缺乏四大海区完整的海洋线虫种类组成和多样性数据。据估计,我国海洋线虫常见种约1 000种[4],目前已鉴定到种的约为500种[5,6]。总体上看,我国海洋线虫分类学研究还处在初级阶段,我国海洋线虫的分类和多样性研究亟待加强。
南海是中国近海中面积最大、最深的海区,位于中国的最南端。南海东接太平洋,西南通印度洋,具有海区广阔、水深跨度大等特点,根据不同水深可划分为河口近岸区(30 m以内水域)、陆架区(30~200 m水域)、大陆坡(500 m以内水域)和深海盆区(大于500 m以上的水域)[7];气候方面,南海地处热带与亚热带区域,热带海洋性气候显著,尤其是中部和南部海区,终年高温高湿,气候变化很小,与其他海区相比拥有丰富的动植物资源与多种热带生态系统。随着我国海洋发展战略的推动,小型底栖生物研究已经逐步从中国近海扩展到深海[8],在国家自然科学基金项目的资助下,前人已对南海东北部及北部湾等区域进行了沉积物样品的采集,并对其小型底栖生物进行分析,对海洋线虫进行分类研究,但关于南海中部深海区域小型底栖生物的研究较为薄弱,对于该区域海洋线虫的分类与生物多样性研究缺乏基础资料。
本文通过对南海中部海盆沉积物的采集,调查研究分析了该海区小型底栖生物和海洋线虫的种类组成和多样性特征,描述了深海双单宫线虫AmphimonhysteramolloyensisTchesunov & Mokievsky,2005,莫帕多毛线虫GreeffiellamoppaSchrage,1972和四腺拟薄咽线虫PararaeolaimustetradenusLeduc,2017三个我国海洋线虫新纪录种,丰富了南海中部区域海洋线虫的生物多样性信息。
1 材料与方法
2020年6月,搭载国家自然科学基金委KK2002共享航次“嘉庚号”考察船对南海中部区域(东经110°45′2″~118°53′54″,北纬10°0′45″~21°32′57″)共计13个站位进行了沉积物样品采集和小型底栖生物的调查研究。使用XCYQ-2.5-2深海沉积物箱式采样器采集未受扰动的海底表层沉积物,将采集好的沉积物样品用由医用注射器改造的内径为2.6 cm的采样管对表层进行插管取样,本航次同一个站位取四个平行样,每个平行样取表层0~8 cm的沉积物,并按照0~2 cm、2~5 cm、5~8 cm分层,将分层的样品分别装入125 mL的塑料样品瓶中,用与样品等体积的10%海水福尔马林溶液进行固定,用作小型底栖生物的研究,另外刮取部分表层的沉积物作为站位样品的定性补充。
实验室内样品的分选、制片等方法参照作者相关文献介绍,分选下限为31 μm孔径网筛。将制备好的永久装片利用微分干涉显微镜(Olympus BX53)对线虫进行形态学的观察鉴定,利用Olympus DP73与CellSens standard软件进行相关数据的测量与拍照绘图工作,对线虫的结构进行测量时,弯曲结构沿弧线或身体中线测量,测量3次取其平均值,单位为微米。分类鉴定时主要依据一系列形态学特征以及德曼比值(a、b、c)等指标,对照Warwick 等分属图示检索表[9],按照De Ley等人于2004年修订的分类系统对海洋线虫进行分类鉴定[10],参考Bezerra等国际线虫数据库[11]以及国内外线虫分类学家的大量文献资料进行种或分类实体的鉴定。凭证标本存放于聊城大学生命科学学院小型底栖生物研究室。
2 结果与讨论
2.1 自由生活海洋线虫的丰度和多样性
本航次样品共分选出小型底栖生物类群14个,其中包括线虫、桡足类、多毛类、寡毛类、涡虫、端足类、介形类、动吻类、海螨类、等足类、异足类、轮虫、缓步动物等,平均丰度为(204.4±55.6)ind./10 cm2,其中线虫占小型底栖生物总丰度的76.2%,平均丰度为(155.8±27.6)ind./10 cm2。共鉴定出海洋线虫275种或分类实体,隶属于142个属,39个科,10个目,2个纲。优势属为AcantholaimusAllgén,1933,Halalaimusde Man,1888,CervonemaWieser,1954和Microlaimusde Man,1880等。本文描述了海洋线虫3个中国新纪录种,即AmphimonhysteramolloyensisTchesunov & Mokievsky,2005,GreeffiellamoppaSchrage,1972和PararaeolaimustetradenusLeduc,2017。
2.2 新纪录种描述
2.2.1 深海双单宫线虫AmphimonhysteramolloyensisTchesunov & Mokievsky,2005(表1,图1~2)。该种隶属于双单宫线虫属Amphimonhystera,隆唇线虫科Xyalidae,单宫目Monhysterida。该属由Allgén于1929年建立[12],目前全世界已知8个有效种。该属的主要特征是虫体呈棕色,外唇刚毛与头刚毛排成一圈,化感器较大,圆形或椭圆形,或有一个内部角质化开口,角皮环纹明显,尾锥柱状。
该研究标本分别发现于南海中部S1站位(东经118°52′42″,北纬21°32′56″,水深2 694 m)、S57站位(东经112°0′26″,北纬17°59′51″,水深2 448 m)和S58站位(东经110°45′1″,北纬15°42′45″,水深733 m),沉积物分别为粘土含少量细砂、极细腻灰色砂泥以及表层灰黄、质地软的粘土。发现并测量了2个雄体和1个雌体,凭证标本号分别是NH-S58-193-6、NH-S1-321-1和NH-S57-178-1。测量数据见表1。
雄体:身体细长,逐渐向两端变细,虫体中部最宽。表皮薄且有明显的横纹。体刚毛稀疏不规则排列。内唇感受器未观察到,6根外唇刚毛稍长于头刚毛,并与4根头刚毛排列成一圈。化感器大,椭圆状,近端窄,远端宽,大小为15.3~17.6×9.7~11.3 μm,距头端8.7 μm。口腔锥状,深4.2~4.7 μm,宽2.8~3.3 μm,口腔壁轻微角质化。咽柱状,细长,基部稍膨大。贲门圆锥形,被肠组织包围。神经环位于咽的中部,距前端71.4 μm。排泄系统未观察到。尾锥柱状,长约121~143 μm。尾端具2根刚毛,长2~3 μm,具3个尾腺细胞。
生殖系统具2个相对排列的精巢。交接刺2次弯曲呈倒S形,近端头状,远端逐渐变尖,长约1.6倍泄殖孔相应体径。具小而简单的引带,无引带突。
雌体:个体稍大,尾稍短。生殖系统单子宫,具1个向前伸展的卵巢。雌孔位于虫体后部,约体长的2/3处,距前端594 μm。具1个充满精子的长袋状的储精囊。
本研究发现于南海的标本特征与Tchesunov和Mokievsky最初描述的发现于格陵兰岛与斯瓦尔巴群岛之间北冰洋海域5 416 m深海的该种模式标本的特征基本一致[13],只是南海的标本雄体具有简单的引带,表皮上没有明显的色素颗粒,其他特征完全一致。

表1 深海双单宫线虫Amphimonhystera molloyensis个体特征测量值(μm,除a、b、c、c′和V%)

注:(a) 雄体前端,示头刚毛、口腔、化感器;(b) 雄体尾端;(c) 雌体全貌;(d)交接刺和引带;标尺:(a、d)10 μm;(b) 20 μm ;(c) 100 μm。图1 深海双单宫线虫Amphimonhystera molloyensis手绘图

注:(a) 雄体前端,示头刚毛、口腔、化感器;(b) 雌体前端,示头刚毛、口腔、化感器;(c) 雄体尾端;(d) 雄体泄殖腔区,示交接刺、引带,标尺;(a、d) 10 μm;(b) 20 μm ;(c) 100 μm。图2 深海双单宫线虫Amphimonhystera molloyensis显微照片
2.2.2 莫帕多毛线虫GreeffiellamoppaSchrage,1972 (表2,图3~4)。该种隶属于多毛线虫属Greeffiella,带线虫科Desmoscolecidae,带线虫目Desmoscolecida。该属由Cobb于1922年建立[14],目前全世界已知13个有效种。该属的特征是具有明显体环,体环中部具有横向排列的体刚毛,且遍布全身,较长。头部圆形,基部小于第一个体环宽度,个体较小,长200~400 μm。
该研究标本发现于南海中部S69站位(东经116°27′21″,北纬11°52′40″)和S74站位(东经115°8′50″,北纬10°0′45″),水深分别为2 670 m和1 951 m,沉积物为黄灰色钙质粘土。发现并测量了2个雄体,凭证标本号分别是NH-S69-250-1和NH-S74-252-1。测量数据见表2。
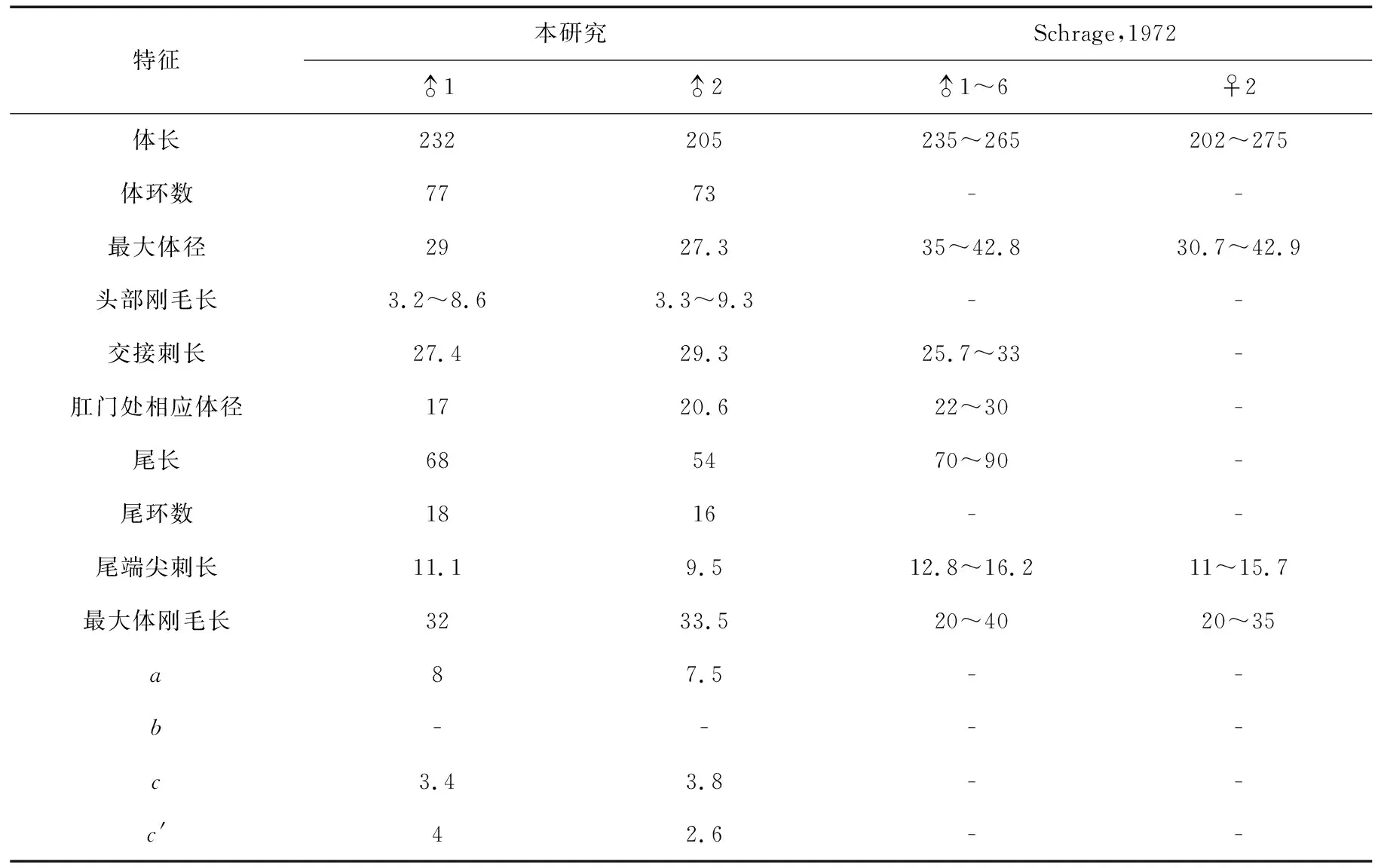
表2 莫帕多毛线虫Greeffiella moppa个体特征测量值(μm,除a、b、c和c′)
雄体:虫体较小,长205~232 μm,呈长纺锤形,由中部向两端逐渐变细,由73~77个体环组成,每个体环中央都有一排较细的体刚毛,由身体前部向后部逐渐变长,最长的体刚毛长31.9~33.4 μm,位于虫体中部。头端半圆形,两侧各具1个大的椭圆形化感器,头最宽处10.2~13.1 μm,基部宽7.9~8.4 μm。4条头刚毛粗短,长3.2~3.3 μm。口腔小。咽基部膨大。尾长锥状,长53.9~68.1 μm,为泄殖孔相应体宽的2.6~4倍,尾部具16~18个体环,末端具1~2根长的尾刺,刺长9.5~11.1 μm。
生殖系统具1个较短的伸展的精巢。一对交接刺等长,较直,近端膨大为明显的头状,由远端向近端逐渐变尖,长27.4~29.3 μm,为泄殖孔相应体径的1.4~1.6倍。无引带,无肛前辅器。
雌体未发现。
本研究发现于南海中部海盆的雄性标本特征与最初由Schrage于1972年描述的发现于北海斯卡格拉克海峡的该种模式标本特征基本一致[15],只是南海标本个体稍纤细(最大体宽27.3~29 μm :30.7~42.9 μm),尾稍短(54~68 μm :70~90 μm),其他特征均相符。

注:(a)雄体前半部分,示头、化感器、咽和体刚毛;(b)雄体后半部分,示交接刺和尾端;标尺:(a),(b) 10 μm。图3 莫帕多毛线虫Greeffiella moppa手绘图

注:(a)雄体全貌,示头、体纹和体刚毛分布;(b)雄体全貌,示头、尾和交接刺;(c)雄体前端,示头部刚毛和化感器;(d)雄体尾部,示交接刺近端;标尺:(a、b) 20 μm;(c、d) 10 μm。图4 莫帕多毛线虫Greeffiella moppa显微照片
2.2.3 四腺拟薄咽线虫PararaeolaimustetradenusLeduc,2017 (表3,图5~6)。该种隶属于拟薄咽线虫属Pararaeolaimus,双盾线虫科Diplopeltidae,薄咽线虫目Araeolaimida。该属由Timm于1961年建立[16],目前全世界已知4个有效种。在该属的原始描述中,虫体无口腔,但在之后发现的种类以及本标本的观察中发现,该属个体拥有较小的口腔,无齿。该属另一个显著特征是雌性个体与雄性个体的化感器异形,雌体的化感器较雄性小,为单圈的螺旋形或1.5圈的螺旋形化感器,且口腔较雄性大,双卵巢。尾锥状,具有较为粗钝的尾尖。
该研究标本发现于南海中部S74站位(东经115°8′50″,北纬10°0′45″)和S79站位(东经114°58′57″,北纬10°45′2″),水深分别为1 951 m和2 390 m,沉积物为黄灰色钙质粘土。发现并测量了4个雄体和1个雌体,凭证标本号分别是NH-S74-274-4、NH-S74-276-4、NH-S79-289-6、NH-S79-293-1、NH-S79-281-1。测量数据见表3。
雄体:虫体呈圆柱状,由中部向两端逐渐变细,头端较为圆钝,尾端呈短锥状。雄体长526~601 μm,最大体宽26.7~33.9 μm,角皮环纹不明显,无体刚毛。内唇感受器与外唇感受器均不明显,四根头刚毛较短,长1.2 μm左右。化感器较大,其顶部距前端16.9~19.8 μm,即0.77~0.96倍相应体径,呈向下开口的圆环状,长11.1~13.5 μm,宽10~11.7 μm,宽占相应体径的48.6%~59.7%。口腔较小,呈杯状,无齿,深3.1~4.1 μm,口腔壁轻微角质化。咽基部膨大成咽球。神经环位于咽的前部,距前端55.6~63.1 μm,占咽长的39.2%~47.7%。排泄系统具两对明显的腹腺细胞,大小26.9×16.5 μm~36.2×20.7 μm,位于咽的下面,肠的前端。贲门锥状,长12.2 μm。尾锥状,长72.3~80 μm,为泄殖孔相应体宽的3.1~4.1倍,三个尾腺细胞明显,末端具明显的黏液管。
生殖系统具有两个反向排列直伸的精巢,前精巢位于肠的左侧,后精巢位于肠的右侧。一对交接刺等长,交接刺稍弯曲,呈匕首状,长18.6~24.4 μm,为泄殖孔相应体径的0.8~1.2倍。无引带,无肛前辅器。
雌体:个体较小,长496 μm,且具有更低的a值与c值。口腔较雄性大,深7.3 μm,化感器与雄性具有明显的不同,较小,单圈螺旋形,长4.5 μm,宽6.8 μm,占相应体径的25.4%。腹腺细胞较雄性小,大小18.3×14 μm。生殖系统双卵巢,前卵巢位于肠的左侧,后卵巢位于肠的右侧。受精囊不明显。雌孔开口于腹面中部靠后,距离头端为体长的58%。尾较雄性长,85.9 μm,为肛门相应体宽的4.2倍。
本研究发现于南海中部的标本特征与Leduc最初描述的发现于新西兰北岛东南部大陆坡深海1 217 m的该种模式标本的特征基本一致[17],只是南海的标本个体稍粗(28~34 μm :26 μm),尾稍长(70~86 :55~76 μm),其他特征完全一致。

表3 四腺拟薄咽线虫Pararaeolaimus tetradenus个体特征测量值(μm,除a、b、c、c′和V%)
3 结论
本研究通过形态学分类方法对采集自南海中部海盆的海洋线虫进行了分类鉴定,共发现鉴定275种或分类实体,优势属为AcantholaimusAllgén,1933、Halalaimusde Man,1888、CervonemaWieser,1954和Microlaimusde Man,1880等典型的深海类群。与南海东北部海区293 种[18],北部湾120种相比,南海中部深海区海洋线虫丰度较低,但多样性较高。本文描述了AmphimonhysteramolloyensisTchesunov & Mokievsky,2005、GreeffiellamoppaSchrage,1972和PararaeolaimustetradenusLeduc,2017海洋线虫中国三个新纪录种。其中,PararaeolaimustetradenusLeduc,2017在南海的首次发现,是Pararaeolaimus属在中国海域的首次纪录,为中国线虫多样性增加了新的属和种的纪录。并且,本研究丰富了我国南海海洋线虫的物种多样性信息,进一步扩大了这三个物种的地理分布范围。
