文王浓香型白酒不同深度新老窖泥理化性质与微生物演替分析
2022-10-27徐相辉许姗姗许博阳穆冬冬吴学凤李兴江
徐相辉,常 强,孙 伟,许姗姗,许博阳,穆冬冬,吴学凤,李兴江,
(1.合肥工业大学食品与生物工程学院,安徽合肥 230601;
2.安徽文王酿酒股份有限公司,安徽临泉 236400)
白酒根据风味特点的不同可分为12种香型,其中浓香型白酒的产量占酒类行业的70%以上,是中国主要的白酒消费品。浓香型白酒采用传统的固态发酵工艺,以高粱、小麦、玉米、糯米和大米为原料,稻壳为疏松剂,以中温大曲为发酵剂在泥窖中发酵60~90 d后蒸馏制得。酒窖底部和侧面均覆盖有窖泥,独特的泥窖发酵工艺赋予了浓香型白酒窖香浓郁,绵甜爽净的特点。大量研究表明,窖泥的稳定和成熟度与浓香型白酒的品质和特征紧密相关,实际生产中也发现“老窖产好酒”。窖泥的成熟是一个长期驯化的过程,新窖泥到成熟稳定状态的老窖泥要经历初驯期、过渡期和成熟期三个阶段,需要较长的时间才能完成成熟转换。
浓香型白酒的发酵过程涉及大曲、环境和窖泥多种微生物,也是微生物群落的演替以及各种微生物共生、共酵的过程。通过对糟醅微生物进行溯源分析发现,糟醅中的原核微生物主要来自窖泥。窖泥被认为是原核微生物的持续释放源,同时糟醅中的原核微生物在窖泥成熟和稳定中也扮演了重要角色。郑佳等研究发现,靠近窖泥的糟醅蒸馏得到的原酒挥发性风味物质含量更加丰富,窖池边缘的糟醅中己酸、丁酸以及它们对应的己酸乙酯、丁酸乙酯含量也高于同期窖池中间的糟醅。乳酸菌、芽孢杆菌等原核微生物产生己酸、丁酸、乙酸和乳酸等各种有机酸,作为酯类化合物的底物,参与了糟醅中风味化合物的形成。
相比于池壁,池底窖泥所处环境氧气更少,并长期浸泡在酸性的黄水中,窖泥微生物所处环境更为恶劣,因此对于窖泥理化与微生物的关联分析有助于进一步指导新老窖泥的养护。同时,窖泥的成熟是一个连续动态的过程,伴随着各类微生物的此消彼长。当前,连续的发酵以及不合理的养护容易造成窖泥板结和窖泥微生物衰亡,最终导致窖泥退化,生产出的白酒品质不佳。目前已有关于大曲和糟醅群落演替的研究,也有对新窖泥前四轮次发酵的窖泥微生物演替分析,而对于窖池垂直维度的微生物群落结构的解析和演替鲜有报道。本研究通过对安徽文王贡酒新老窖池不同深度的窖泥细菌微生物进行分析,理清窖泥深度与其理化性质、有机酸含量和窖泥微生物菌群结构的相关关系,为窖泥的稳定成熟及窖池的养护提供参考。
1 材料与方法
1.1 材料与仪器
本实验所用窖泥样品来自正常生产的安徽文王酿酒股份有限公司3年(new)和30年(old)窖池,采样点经纬度 33°04′N;115°15 ′E,采样时间为 2020年12月。取样示意图如图1所示,从池底3个位置取样混合在一起进行分析,每个采样点分为0~2、2~4、4~6、6~8、8~10 cm 5 层,新窖泥标注为 new_02~new_810;老窖泥标注为 old_02~old_810。将采集样品于均质袋中充分混匀后置于4 ℃和-80 ℃冰箱,分别用于理化测定和高通量测序分析。

图1 窖泥样品取样位置示意图Fig.1 Schematic diagram of sampling location of pit mud samples
氟化铵、盐酸、磷酸二氢钾、氯化铵、重铬酸钾、硫酸亚铁、酒石酸钾钠、抗坏血酸(均为分析纯)国药集团化学试剂有限公司;纳氏试剂 上海源叶生物有限公司;氯化亚锡、钼酸铵 上海麦克林生化科技有限公司;2-乙基丁酸 梯希爱(上海)化成发展有限公司;Fast DNASPIN试剂盒 美国MP Biomedicals。
PHS-25 pH计 海力辰邦西仪器科技有限公司;UV-500 PC紫外分光光度计 上海元析仪器有限公司;7890 A气相色谱仪 安捷伦科技(中国)有限公司;1525型液相色谱仪 沃特世科技(上海)有限公司。
1.2 实验方法
1.2.1 窖泥理化性质分析 窖泥水分参考国标GB 5009.3-2016采用烘干法;pH是将新鲜窖泥与去离子水按照1:5 g/mL混匀静置后用pH计检测;有效磷参考地标DB 34T 2266-2014,采用氟化铵-盐酸比色法测定;铵态氮根据纳氏试剂比色原理,采用紫外分光光度法;腐殖质参考地标DB 34T 2265-2014,采用重铬酸钾氧化法测定。
有机酸的测定均采用相同的前处理方法,将新鲜窖泥与15%甲醇按照1:9 g/mL混匀,超声处理20 min 再次混匀,8000 r/min 离心10 min,用0.22 μm有机滤膜过滤后测定有机酸。丁酸与己酸采用气相色谱内标法定量,将前处理样品加2-乙基丁酸作为内标,色谱柱采用 DB-Wax(Agilent 30 m×0.25 mm),进样量1 μL,柱流速1 mL/min,进样口温度250 ℃,分流比30:1。升温条件:初始温度35 ℃,2 ℃/min升温至 60 ℃,保持 4 min;6 ℃/min升温至 195 ℃,保持20 min。氢火焰离子检测器温度250 ℃。乙酸与乳酸的测定采用高效液相色谱法,色谱柱采用Ultimate LP-C(Welch 4.6 mm×250 mm,5 μm),检测器采用Waters 2996二极管阵列检测器,进样量15 μL,检测波长210 nm。流动相:磷酸二氢钾缓冲液(0.01 mol/L,pH2.50):甲醇=95:5,流速 0.6 mL/min。
1.2.2 窖泥微生物基因组的提取与高通量测序 将采集到的样品用自封袋分装后采用干冰转运,委托上海美吉生物医药科技有限公司进行 Illumina MiSeq高通量测序。窖泥微生物总基因组DNA采用Fast-DNASPIN 试剂盒进行提取,338F_(5’-ACTCCTAC GGGAGGCAGCAG-3’)与 806R_(5’-GGACTACH VGGGTWTCTAAT-3’),扩增微生物 16S rRNA 基因的 V3~V4区。
1.3 数据处理
原始测序数据采用 Flash(1.2.11)和 Fastp(0.19.6)进行拼接和质控,采用Mothur(1.30.2)进行多样性分析,采用QIIME软件(V1.9.1)生成各分类学水平丰度表,多样性距离计算,窖泥理化与样本和微生物之间的冗余分析(RDA)分析借助美吉生信云平台进行分析,采用SPSS 25进行单因素方差显著性分析。
2 结果与分析
2.1 新老窖池窖泥理化性质分析
窖泥的理化如表1所示,新老窖泥的含水量均随窖泥深度增加而降低,pH随窖泥深度增加而升高,除new_02窖泥含水量高于old_02外,同一深度的老窖泥中水分含量和pH高于新窖泥,该结果与前人研究相一致。表层窖泥含水量高、pH低的原因可能是此部分窖泥与呈酸性的黄水直接接触,含水量高的窖泥致密性更强,厌氧效果也更好。老窖泥中的有效磷含量高于新窖泥,老窖池中有效磷含量随窖泥深度增加先上升后下降,新窖池中的有效磷含量随窖泥深度增加而增加。有效磷是微生物生长繁殖和代谢的必要元素,但含量控制在一定范围(1.5~3.0 g/kg),过高的浓度可能会导致窖泥板结,影响窖泥微生物活动致使窖泥退化,本次检测的老窖池中有效磷含量适宜,新窖池中含量偏低,在新窖泥养护中应适量添加含磷介质。氨态氮的含量与窖泥质量呈明显正相关,在本文研究中发现老窖池中氨态氮含量远高于新窖泥,老窖池不同深度的窖泥中含量差异不明显,在新窖池中随窖泥深度增加而增加,新窖泥在深层质量更好。腐殖质是由微生物分解有机质产生的,其含量可以用于判断窖泥质量,有学者对不同腐殖质含量的窖泥在实际生产中进行了验证,发现窖泥腐殖质含量高低与所产的原酒中己酸乙酯含量有相关性,但在本研究中发现新老窖泥中腐殖质含量均随窖泥深度的增加而降低,并且各层之间差异显著(<0.05)。这可能是由于表层窖泥微生物与有机质含量高糟醅直接接触,代谢旺盛产生的,而深层窖泥有机质含量较低。

表1 新老窖池池底窖泥理化性质Table 1 Physicochemical properties of bottom pit mud from old and new pit cellar
窖泥中原核微生物可以产生各种有机酸。有机酸作为酯类物质的底物,参与了糟醅中风味化合物的形成。己酸、丁酸、乳酸和乙酸是浓香型白酒典型风味物质己酸乙酯、丁酸乙酯、乳酸乙酯和乙酸乙酯的前体物质。随窖泥深度的增加,有机酸的含量均降低。相比于乳酸和乙酸,新老窖池中不同深度窖泥中己酸和丁酸含量差异不明显。根据Zhou等的研究,老窖池中有机酸代谢的途径更为活跃,但新窖池中乳酸和乙酸含量明显高于老窖池,这可能与新窖池中占据主导相关,乳酸含量的高低可以作为窖泥成熟的判断标准。
2.2 窖泥中原核微生物α多样性指数分析
对所有测序序列进行过滤,30个窖泥样品的平均长度为416 bp。每个样本的读取次数从33172到68678。在97%的水平,所有优质序列分成4838 OTUs。样本中优质序列的覆盖率均大于99%,说明细菌微生物群落具有较好的代表性。
如表2所示,Shannon指数随窖泥深度增加而增加,Simpson指数随窖泥深度降低,表明越深层窖泥微生物多样性越高;对比新老窖池发现,老窖池的微生物多样性更高。老窖池物种丰富度(Chao指数、Ace指数和Sobs指数)随窖泥深度增加,表层0~4 cm与深层4~10 cm存在显著差异(<0.05);而新窖池表层0~2 cm物种丰富度最大,总体呈现先下降后上升的趋势,但各层间无显著性差异(>0.05)。对比新老窖池的微生物丰富度可以看出,老窖池的层次较为分明,老窖池微生物丰度低于新窖池,这可能是超过30年的窖泥存在退化的原因。
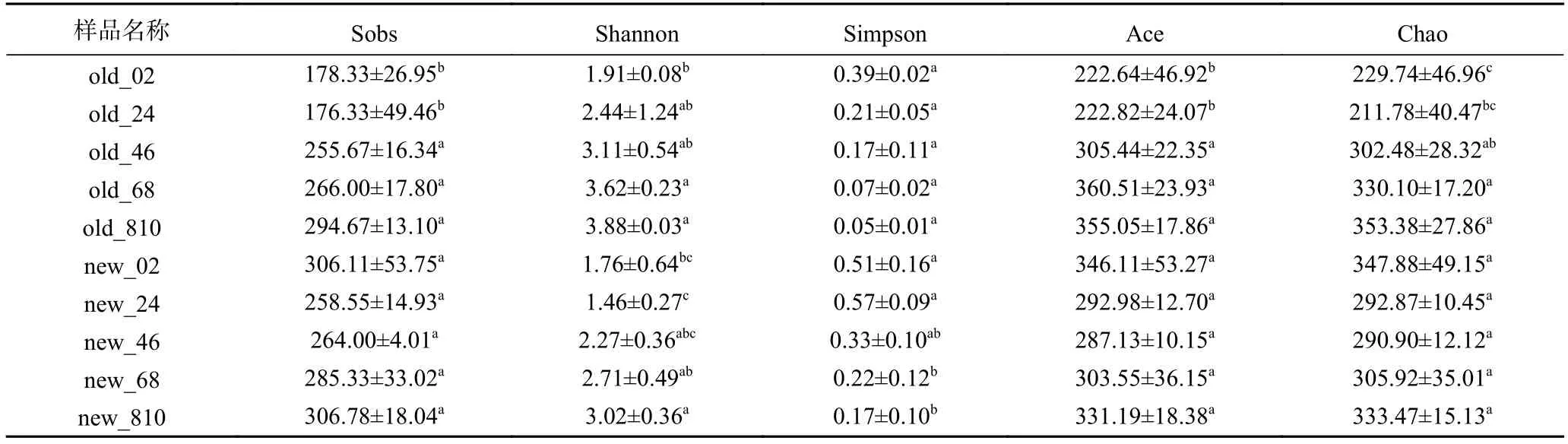
表2 新老窖池池底窖泥原核微生物群落α多样性指数Table 2 α-Diversity index of prokaryotic community in pit mud from old and new pit cellar
从多样性可以看出,窖泥微生物的多样性和丰度呈现出层次差异。如图2所示,对新老窖池不同深度窖泥微生物的物种多样性进行组间非度量多维排列(NMDS)比较分析表明,老窖池0~4 cm和4~10 cm深度的窖泥微生物更紧密地聚集在一起;新窖池呈现相似的规律,但新窖池4~6 cm更趋向于过渡阶段,在两个分组中均有出现。
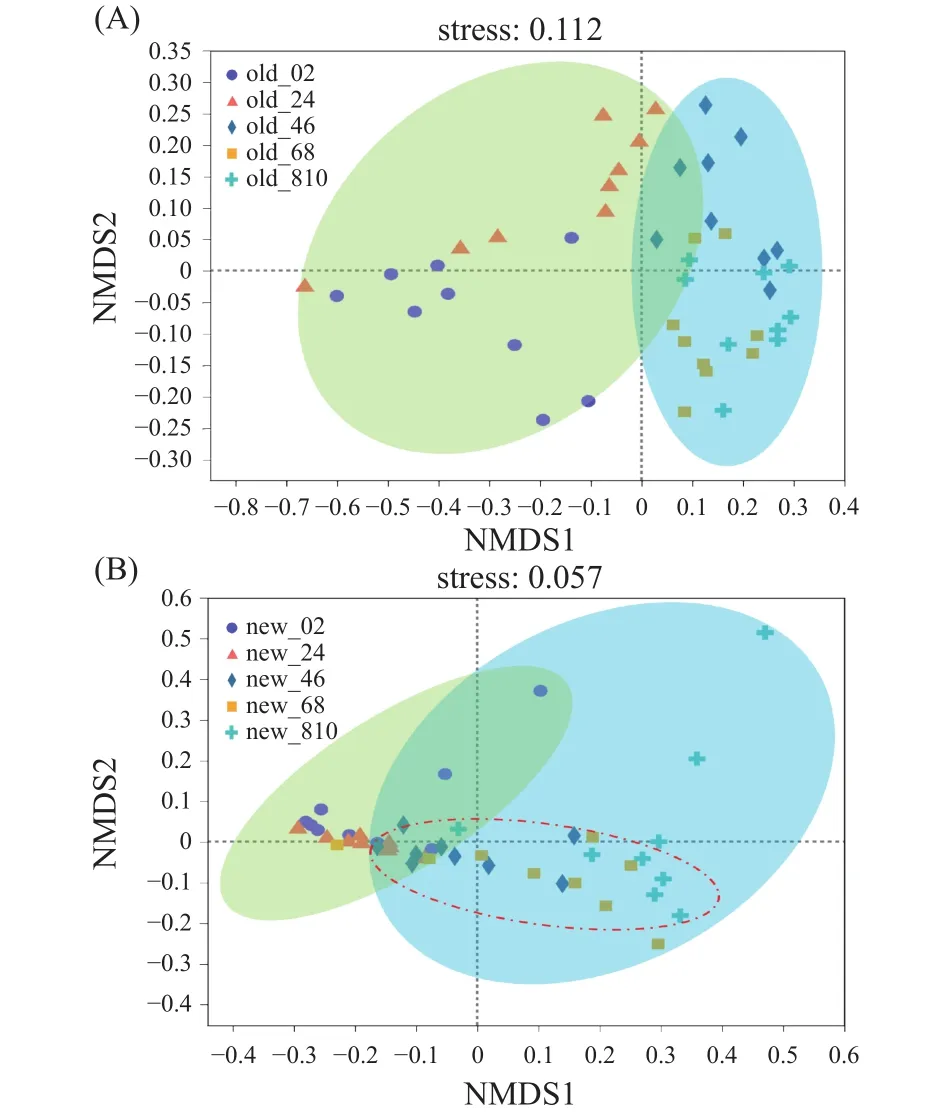
图2 基于新老窖泥细菌群落Bray-Curtis相似性的NMDS排序Fig.2 NMDS ordination based on Bray-Curtis similarities of bacterial communities at old and new PM
2.3 窖泥中原核微生物门层面分析
如图3A所示,在老窖泥样品中微生物共检出35个门,有18个门在所有深度中均存在。如图3B所示,在新窖池中共检测到43个门,有21个门在所有深度中均存在。如图4所示,老窖泥中占比最高的是厚壁菌门(Firmicutes,78.25%),其次是拟杆菌门(Bacteroidota,10.06%),占比超过1%的还有互养菌门(Synergistota,4.02%)、Patescibacteria(2.4%)、Caldatribacteriota(1.45%)和放线菌门(Actinobacteriota,1.31%)。新窖泥中占比最高的也是厚壁菌门(Firmicutes,95.22%),其次是变形菌门(Proteobacteria,2.43%)和放线菌门(Actinobacteriota,1.75%)。随窖泥深度的增加,微生物的门类也相应增加。无论是老窖泥还是新窖泥,厚壁菌门始终在窖泥微生物群落中占据主导地位,该发现与前人研究相似。
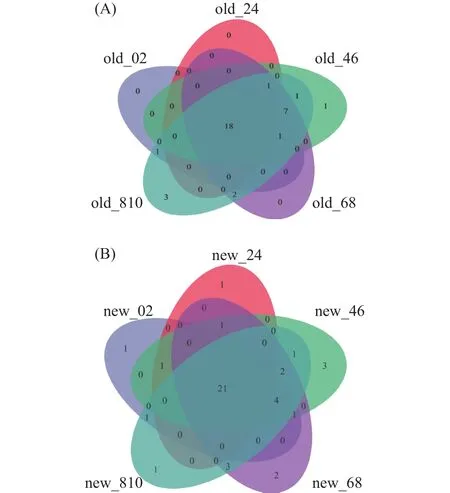
图3 新老窖池不同深度窖泥微生物门水平Veen图Fig.3 Veen diagram of different depths new and old PM at phylum level

图4 新老窖泥门水平物种占比情况Fig.4 The proportion of microbial in old and new PM at phylum level
2.4 窖泥中原核微生物属层面分析
对老窖池的多样性检测发现,相比于新窖池,老窖池的微生物多样性更高。进一步对窖泥微生物群落属层面的分析也进一步验证了这一结论,丰度占比超过0.01%的微生物有37种,而新窖池相对丰度占比0.01%只有16种。如图5A所示,通过对比老窖泥不同深度的微生物占比发现,在浅层窖泥中占据主导地位,深层窖泥中和占据主导。和是重要的酿造功能菌。占比随窖泥深度的增加而降低,在表面0~2 cm的窖泥中占比最高为41.29%,而深层窖泥中占比却低于1%,与前人研究一致。属的微生物占比随窖泥深度增加先上升后下降,在4~6 cm的中层占比最高为23.9%。在6~8 cm窖泥中占比最高,为18.71%。根据前人的研究,和在窖泥成熟稳定中扮演重要的角色,与乳酸呈负相关。新窖池占比超过0.01%的微生物有16个属,如图5B所示,乳酸菌()在窖泥各个深度中所占比例最高,在表层2~4 cm窖泥中所占比例最高(74.7%),但其占比随着窖泥深度的增加而下降。芽孢杆菌()、梭菌(12)和的随着深度的增加占比增加,成为深层窖泥微生物的优势菌群,这可能与深层窖泥的厌氧环境有关,有利于芽孢杆菌、梭状芽胞杆菌等专性厌氧微生物的生存。在属水平上,不同深度的微生物群落演替存在显著差异,说明微生物群落在垂直方向上演替显著。

图5 新老窖池窖泥样本与微生物菌群属水平Circos图Fig.5 Circos diagram for microflora structure in PM samples of old and new pit cellar at genus level
2.5 窖泥理化性质与微生物样本冗余分析
如图6所示,RDA的结果分别解释了老窖池54.00%和新窖池57.21%样本与微生物理化的关系,新老窖池不同深度样品对应的理化具有相似的分布情况。新老窖池中浅层窖泥(0~4 cm)与理化因子水分、己酸、丁酸、乳酸和乙酸和腐殖质正相关,深层窖泥(4~10 cm)与理化因子氨态氮、有效磷和pH正相关。这一发现与陶勇对2~50年不同窖龄窖泥与理化因子的相关性中有效磷、pH和氨态氮对微生物群落结构影响相同。不同的是,老窖池浅层窖泥与微生物正相关,能够促进己酸的产生。有研究表明,随窖龄增加其相对丰度增加。深层与微生物和正相关。新窖池浅层窖泥与微生物正相关,深层窖泥与微生物和12正相关。根据对窖泥微生物的研究,这两类微生物参与窖泥的老化成熟。

图6 窖泥理化指标和微生物群落样本冗余分析Fig.6 Redundancy analysis of physicochemical indexes and microbial community in pit mud
2.6 有机酸与窖泥微生物相关性分析
浓香型白酒有机酸中,己酸、丁酸、乳酸和乙酸构成总酸的90%以上。窖池中产生有机酸的微生物主要来自窖泥,为了揭示窖泥中产有机酸微生物,本文用热图展现了有机酸与相对丰度排名前20的微生物属间的Spearman相关系数。从图7A可以看出,老窖池中微生物聚为2类,其中、与四种有机酸呈正相关,为老窖池中有机酸合成关键微生物。与己酸合成正相关的微生物有4种,分别是、、和。如图7B所示,新窖池中与有机酸相关的微生物同样聚为 2类,其中、和与除丁酸外的有机酸呈正相关,是新窖池中主要有机酸合成的关键微生物。主要代谢产物为己酸、丁酸、乙酸和氢气,在新老窖池中均参与有机酸的生成。
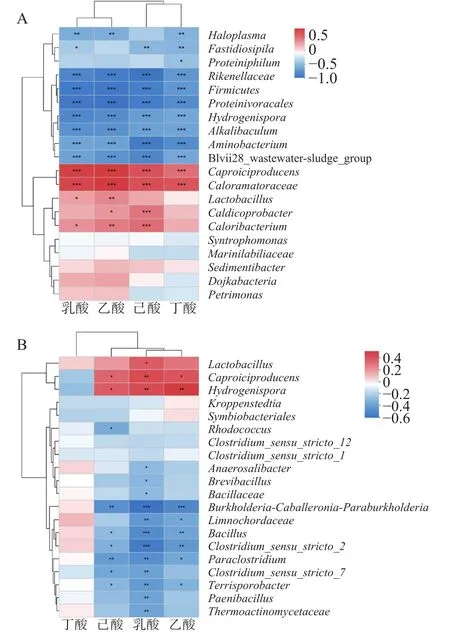
图7 窖泥有机酸与微生物相关性热图Fig.7 Heatmap of correlation between pit mud organic acid and microorganism
3 结论
本研究采用Illumina MiSeq高通量测序分析了3、30年新老窖池不同深度窖泥的原核微生物群落演替,结合理化相关性分析,得到结论:新老窖池理化存在显著差异,老窖池中水分、pH、有效磷、氨态氮含量普遍高于新窖池;新窖泥中乳酸和乙酸含量明显高于老窖泥,乳酸含量的高低可以成为成熟窖泥的评价指标。新老窖池微生物存在显著差异,老窖池的微生物多样性更高而丰富度低,和是老窖泥中的优势微生物 ,、和是新窖泥的优势微生物。垂直维度存在明显群落演替。深度4 cm是区分表层深层的分界线;随窖泥深度的增加,和成为老窖泥中优势微生物,和12成为新窖泥的优势微生物。新老窖泥理化关联分析具有相似性,新老窖池深层窖泥与理化因子氨态氮、有效磷和pH均呈正相关;新老窖泥有机酸关联分析具有差异性,、是老窖池中有机酸合成关键微生物,、和是新窖池中主要有机酸合成的关键微生物。
综上,本研究系统测定了垂直维度不同深度新老窖池池底窖泥的理化与微生物菌群结构,首次提出4 cm是区分窖泥深浅的分界线,并通过冗余分析阐明了不同深度样本与理化因子的相关性,并与有机酸进行关联,找出了与有机酸生成相关的微生物,丰富了新老窖泥在垂直维度上的研究,对于指导窖泥养护和科学研究采样具有一定现实意义。
