Quantitative Trait Locus Mapping of High Photosynthetic Efficiency and Biomass in Oryza longistaminata
2022-10-25SiFengfengFanFengfengWeiXiaoHeShihaoLiXianlongPengXiaojueLiShaoqing
Si Fengfeng, Fan Fengfeng, Wei Xiao, He Shihao, Li Xianlong, Peng Xiaojue, Li Shaoqing
Research Paper
Quantitative Trait Locus Mapping of High Photosynthetic Efficiency and Biomass in
Si Fengfeng1, 2, Fan Fengfeng1, Wei Xiao1, He Shihao1, Li Xianlong1, Peng Xiaojue2, Li Shaoqing1
(State Key Laboratory of Hybrid Rice / Hongshan Laboratory of Hubei Province / Key Laboratory for Research and Utilization of Heterosis in indica Rice of Ministry of Agriculture and Rural Affairs / Engineering Research Center for Plant Biotechnology and GermplasmUtilization of Ministry of Education / College of Life Science, Wuhan University, Wuhan 430072, China; Key Laboratory of Molecular Biology and Gene Engineering of Jiangxi Province / College of Life Science, Nanchang University, Nanchang 330031, China)
Photosynthetic efficiency, a key trait that determines yield potential in rice, is quantitatively regulated by multiple genes. Utilization of valuable genetic resources hidden in wild rice is an effective way to improve rice photosynthesis and yield potential. In this study, 152 backcross inbred lines derived from wild ricewere explored for QTL mapping of photosynthetic rate (Pn) and biomass (BM) in natural fields. Five novel QTLs for Pn and seven QTLs for BM or daily biomass (DBM) derived fromwere identified. One of these QTLs,, could significantly improve Pn and was located in a 68-kb region containing only 11 candidate genes. Meanwhile,andforBM and DBM on chromosome 1 were overlapped withfor Pn from 9311, and could affect both Pn and BM in natural fields. These QTLs identified inmay provide a novel alternative to explore new genes and resources for yield potentiality, highlighting the important role of wild rice in rice breeding programs.
QTL;; photosynthetic rate; biomass; wild rice
Rice (L.) is a staple food that feeds over half of the world’s population, and increasing its production is crucial to solve global food crises in the future (Halder et al, 2016). Over the past decade, rice production in the world has more than doubled owing to the exploitation of a series of favorable genes including semi-dwarf geneand hybrid rice-related genes such as,and–, which confer the plant stronger lodging-resistance, higher harvest index and higher grain yield (Monnaet al, 2002; Kazama et al, 2016; Omukai et al, 2021). The popularization of semi-dwarf rice and hybrid rice results ‘Green Revolution’ in the 1960s‒1980s and promotes the explosion of world’s population, which is predicted to reach to 9 billion by 2050 at current increasing trend, and will require 70% to 100% furtherincrease in rice planting area (Godfray et al, 2010a, b; Ray et al, 2013). Thus, increasing rice grain yield of unit land area may become the sole efficient and practical way to meet the severe and urgent food challenge. Gain in grain yield is the product of the plantbiomass (BM) and harvest index (Kemanian et al, 2007). However, improvement of harvest index is becoming increasingly harder as it has been optimized to a biological maximum in present major cereal crops such as rice and wheat (Foulkes et al, 2011; Driever et al, 2014; Furbank et al, 2020). Therefore, yield improvements will have to depend more on efficient BM accumulation and high photosynthetic efficiency in contemporary rice research (Zhu et al, 2008, 2010; South et al, 2018; Mathan et al, 2021).
BM accumulation is the biological amassing capacity of plants to use light for growth, which reflects the photosynthetic capacity to some extent (Mann, 1999; Kartika et al, 2020). Leaf photosynthesis is the main source of carbon assimilation. The flag leaf, as the rice simulation models indicate, play a primary role in supplying carbon to the developing inflorescence, and it has long been recognized as a major contributor of photo assimilates to the grain (Zhu et al, 2008, 2010). However, agronomically important traits, including plant height and grain yield of rice, typically has more complicated genetic architecture and are often governed by quantitative trait loci (QTLs) with minor effects. Particularly, in the case of QTLs for physiological traits, such as photosynthetic rate (Pn), are hard to be effectively detected and efficiently employed in traditional rice breeding programs (Teng et al, 2004; Reynolds and Langridge, 2016; Haritha et al, 2017). From this point, identification and utility of genetic variations with high efficiency in photosynthesis at the leaf level using modern biotechnology will become a major target in rice breeding research (Long et al, 2015; Furbank et al, 2019).
Wild rice, especially AA genomespecies including,the ancestor of Asian cultivated rice (Huang et al, 2012), is suggested as an extremely important genetic pool for rice improvement, as they live over hundreds of thousands of years in different harsh natural environments and accumulate substantial favorable genetic variations (Sweeney and McCouch, 2007; Kartika et al, 2020; Mathan et al, 2021). However, it is estimated that at least one third of the functional genes from the wild ancestor have been lost in the cultivars, such asforgrain size (Takano-Kai et al, 2009; Mao et al, 2010),for grain production (Huo et al, 2017; Yu et al, 2017), andfor seed dormancy (Sugimoto et al, 2010). It means that re- exploitation of the valuable genetic variations hidden in wild rice will effectively promote the sustainable development of rice industry (Adachi et al, 2020).
, an old ancestor living in the Africa of the AA genome species, is attracting great attention of the rice breeders because of its specific characters with large panicle, brawny and strong stature, and giant BM accumulation (Kovach et al, 2007; Sweeney and McCouch, 2007; Thalapati et al, 2012). In order to uncover the latent enigma at genetic level, here we performed a QTL analysis of BM and Pn in flag leaves using a backcross inbred line (BIL) population derived fromand 9311. There were seven QTLs for BM and five QTLs for Pn identified fromin two different sites. Among them, one QTL for Pn and one QTL for BM were repeatedly identified, which may provide an alternative strategy to increase rice productivity in the future high-yield breeding programs.
Results
Photosynthesis and BM of BIL population in fields
In August 2020 and April 2021, the Pn and BM of BIL population (152 BILs) were investigated in Ezhou, Hubei Province and Lingshui, Hainan Province, China respectively. Pn, stomatal conductance (Gs), BM and daily biomass (DBM) showed continuous and normal distribution patterns in the BIL population (Table 1; Fig. 1), meaning that the photosynthesis and BM are complicated traits and controlled by multiple genes. However, it was apparent that the distribution patterns of Pn, Gs, BM and DBM were highly similar in the two sites (Fig. 1), reflecting that these four traits had high positive relationships.
Moreover, the Pn of BILs ranged from 10.30 to 36.20 µmol/(m2·s) with a mean of 22.00 µmol/(m2·s)in Ezhou andranged from 17.20 to 32.60 µmol/(m2·s) with a mean of 26.20µmol/(m2·s) in Lingshui. The two means of Pn of the BILs were larger than those of the recurrent parent 9311, indicating that the wild rice had stronger Pn than 9311 in Ezhou and Lingshui. Gs is another important index of photosynthesis which indicates the capacity of a plant to assimilate CO2to some extent. In Ezhou, the Gs ranged from 0.30 to 1.21 mol/(m2·s) with a mean of 0.68 mol/(m2·s), and in Lingshui, the Gs ranged from 0.31 to 1.32 mol/(m2·s) with a mean of 0.80 mol/(m2·s) (Table 1). Although both of the means of Gs of the BILs were less than Gs of 9311, the largest Gs of the BILs surpassed over 50% of that of 9311, highly consistent with Pn, meaning that the Gs in part of BILs was stronger than that in 9311.
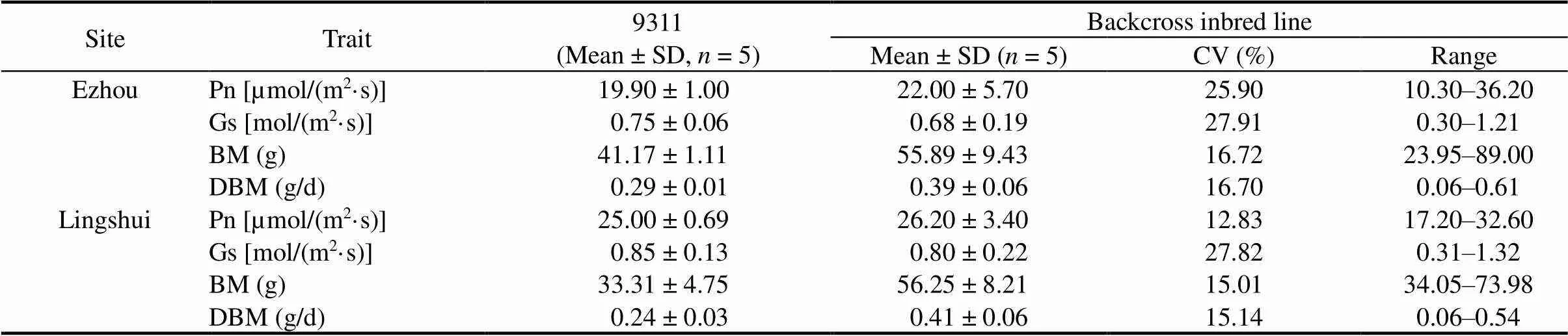
Table 1. Pn and BM of backcross inbred lines in different sites.
Pn, Photosynthetic rate; Gs, Stomatal conductance; BM, Biomass; DBM, Daily biomass; CV, Coefficient of variation.
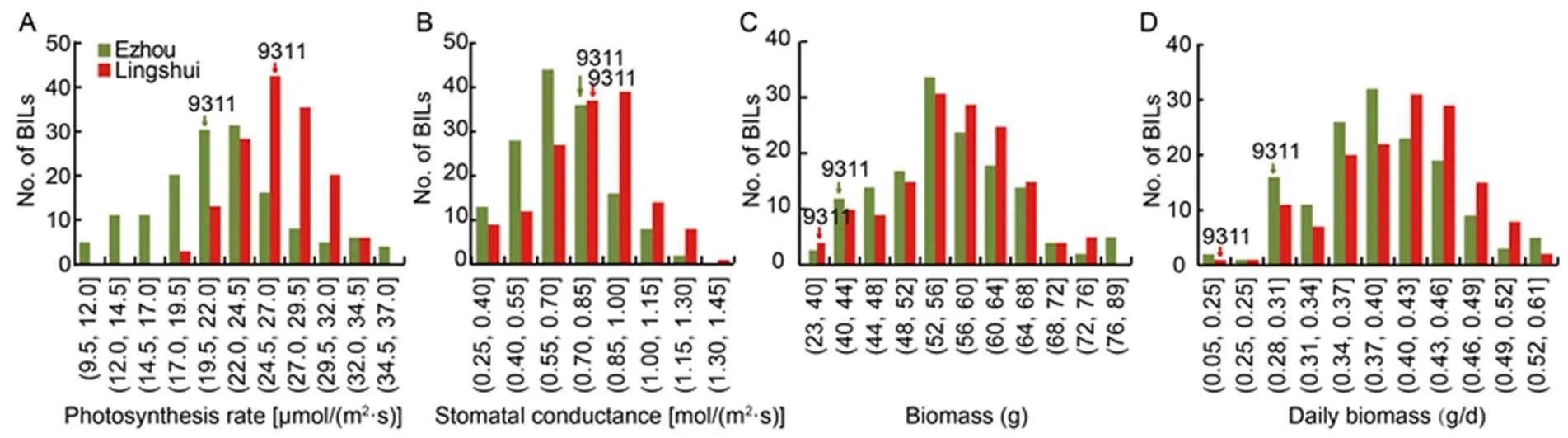
Fig. 1.Frequency distributions of photosynthetic rate (A), stomatal conductance (B), biomass (C) and daily biomass (D) of backcross inbred lines (BILs) in different sites.
BM is the dry weight of a plant above ground, whichcomes from the accumulation of photosynthesis. Similar to Pn, BM and DBM exhibited great variations among BILs (Table 1). Investigation showed that BM ranged from 23.95 to 89.00 g with a mean of 55.89 g in Ezhou, and from 34.05 to 73.98 g with a mean of 56.25 g in Lingshui, respectively. Similarly, the DBM ranged from 0.06 to 0.61 g/d with a mean of 0.39 g/d in Ezhou, and from 0.06 to 0.54 g/d with a mean of 0.41 g/d in Lingshui, respectively. The giant variations of BM and DBM among the BIL population indicated that these traits were controlled by a highly complicated genetic network. Apart from Gs and Pn, BM and DBM also showed a trend that the means of the BIL population were higher than those of 9311 in the two experimental sites (Table 1).
Pn is positively correlated with BM in rice
In order to explore the relationship between Pn and BM of the BIL population, the correlation coefficients of the four traits were analyzed. Results showed that there was a significant positive correlation between Pn and BM/DBM in Ezhou, similar results were also observed in Lingshui. Correlation analysis showed thatPn of these two sites was positively correlated with Gs, and the correlation coefficients reached 0.437 and 0.588 in Ezhou and Lingshui, respectively. Furthermore,an extremely significant correlation was observed betweenBM and DBM, and the correlation coefficients reached 0.988 and 0.958 in Ezhou and Lingshui, respectively (Table 2), reflecting that both BM and DBM were highly controlled by Pn.
QTL mapping for photosynthesis in BIL population
A total of five QTLs for Pn and one QTL for Gs were identified in the two sites. Among them, five novel QTLs, come from, were the firstly identified (Fig. 2). Five QTLs (,,,and) for Pn were located on chromosomes 1, 2, 5, 8 and 12, explaining 11.9%, 9.2%, 7.9%, 9.2%/8.4% and 7.0% of the phenotypic variations, respectively. One QTL for Gs,was mapped on chromosome 2, and explained about 11.7% of the phenotypic variations (Table 3).
Two BM-related QTLs and five DBM-related QTLs were detected in the two sites (Fig. 2). Among them,andwere located on chromosomes 1 and 6, and explained 8.2%, 15.0% and 8.1% of the phenotypic variations, respectively. For DBM,,,,andwere mapped on chromosomes 1, 3, 4, 6 and 12, and explained 8.1%/13.0%, 4.5%, 7.7%, 9.3% and 5.0% of the phenotypic variations, respectively (Table 3).Importantly,and, as well asandhad the greatest effects on the BM and DBM phenotypic variations, respectively, all of which were come from(Table 3). It is indicated that the QTLs fromplayed a critical role on the BM accumulation. Interestingly, in all of the 13 QTLs, the,andwere repeatedly detected in the experimental field of Ezhou and Lingshui. More importantly,was co-located withandon chromosome 1 and formed a QTL cluster as/. Importantly, according to the genotype of the BILs, this QTL was delimited by L/Bin 1-143 and R/Bin 1-144 to a region with a physical distance about 111.4 kb referenced to the Minghui 63 (MH63) genome, and the BIL lines with this QTL all showed apparently higher photosynthetic rate (Fig. 3), reflecting great potentiality of thelocus fromin the rice improvement.

Table 2. Correlation coefficients among photosynthesis and biomass of backcross inbred lines in Ezhou (upper right) and Lingshui (lower left), China.
Pn, Photosynthetic rate; Gs, Stomatal conductance; BM, Biomass; DBM, Daily biomass. * and ** mean significant differences at the 0.05 and 0.01 levels, respectively.
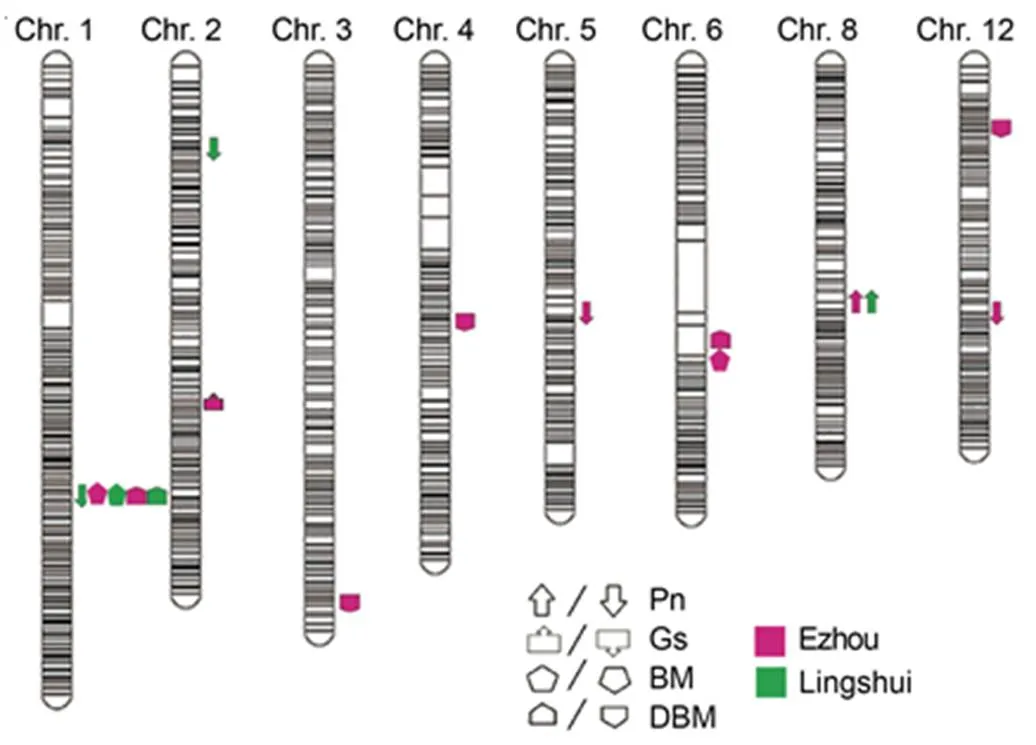
Fig. 2. QTL analysis of photosynthesis ofbackcross inbred line population in different sites.
Upward direction indicates the allele showing positive effects, while downward direction indicates the allele showing negative effects.
Pn, Photosynthetic rate; Gs, Stomatal conductance; BM, Biomass; DBM, Daily biomass.
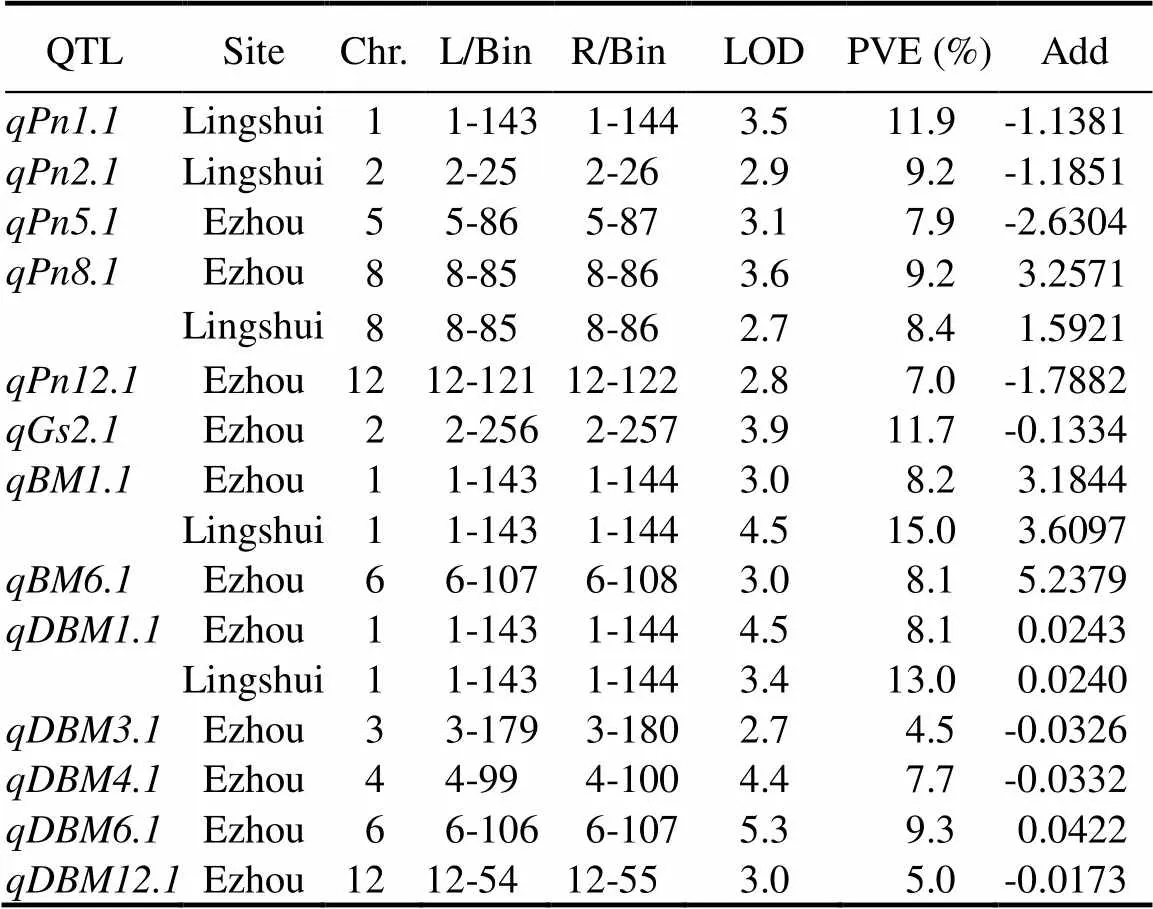
Table 3.QTL information for photosynthic rate and biomass of backcross inbred lines in fields.
L/Bin, Left molecular marker; R/Bin, Right molecular marker; LOD, Logarithm of odds; PVE, Phenotypic variation explained. Add, Additive effect with positive number representing the positive effect fragment from, and negative number representing the positive effect fragment from 9311.
Validation of qPn8.1 from O. longistaminata for Pn
In order to verify the genetic function of the newly identified QTLs for photosynthesis from, we further analyzed the genetic effects of the novel. Results showed thatwas delimited by L/Bin 8-85 and R/Bin 8-86 in a region with a physical distance about 68 kb in MH63 genome (Fig. 4-A). The Pn of the lines harboringwas about 12.71% higher than those without(Fig. 4-B and -C), meaning that thefromhad strongly positive effect on the improvement of rice photosynthesis. Genomic structural analysis showed that there were only 11 genes at the locus of(Table S1), of which, 3 genes showed structural and sequence variations between 9311 and(Fig. S1). Expression analysis showed that theexpression level in the lines harboringwere about four-fold higher than those without(Fig. 4-D). Thus, it is inferred thatmight be the candidate gene for the functionof, which encodes a highly conserved chloroplast- localized protein. These results indicated thatderived fromsignificantly increased the photosynthetic efficiency in the BIL population, meaningharbors some importantly favorable alleles for improving rice photosynthesis.
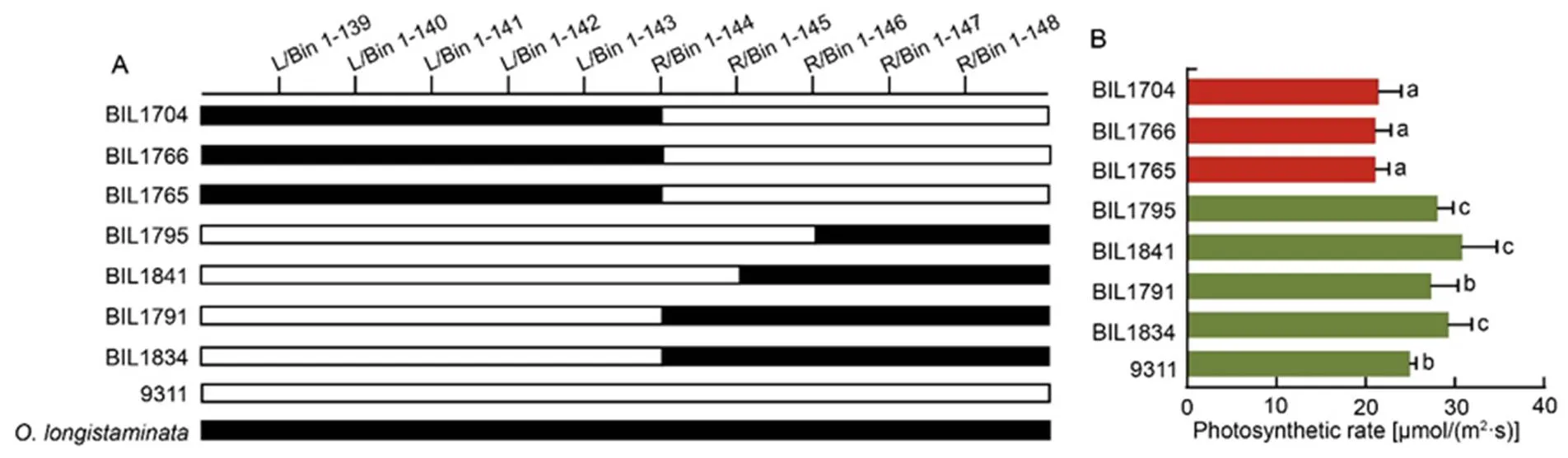
Fig. 3. Validation of function of.
A, Comparison of genotypic structure oflocus in different backcross inbred lines (BILs). Black bar and blank bar represent fragments fromand 9311, respectively.
B, Photosynthetic rate of corresponding BILs.Different lowercase letters at the top of the bar indicate significant differences among different BILs by the Duncan’s multiple range test at< 0.05.
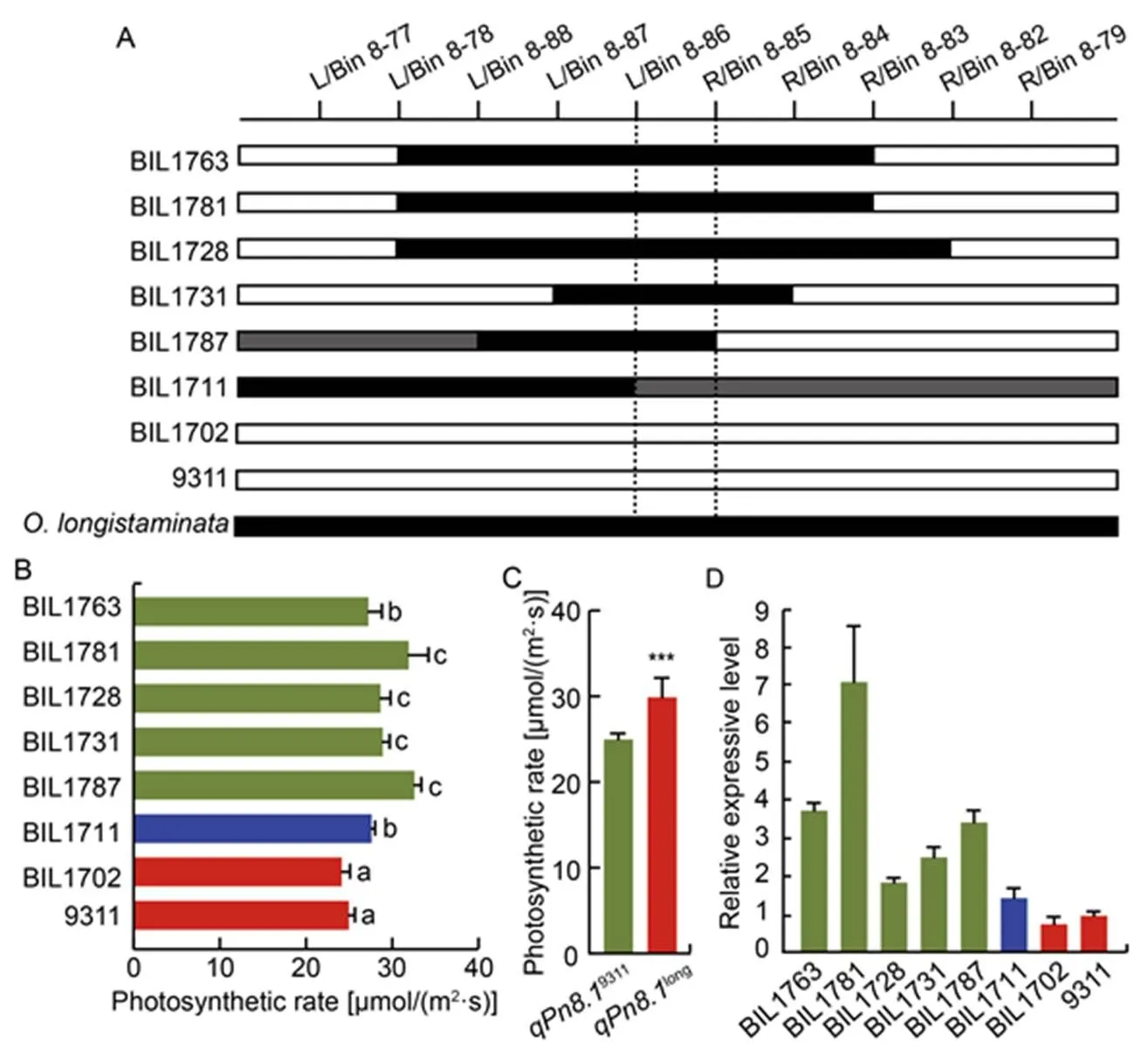
Fig. 4. Validation of function of.
A, Comparison of genotypic structure oflocus in different backcross inbred lines (BILs). Black bar and blank bar represent fragments fromand 9311, respectively. Gray bar representsheterozygote genotype.
B,Photosynthetic rate of corresponding BILs.Different lowercase letters at the top of the bar indicate significant differences among different BILs at< 0.05 by the Duncan’s multiple range test.
C, Means of Pn of BILs with. ***, Significant difference at< 0.001 by the Duncan’s multiple range test.
D, Relative expression levels of candidate genein different BILs and 9311.
In A, B and D, BIL1763, BIL1781, BIL1728, BIL1731 and BIL1787 harborfrom;BIL1702 does not harboras 9311; and BIL1711 is the heterozygote genotype.
Discussion
High Pn and BM accumulation are the basis of high grain yield of rice (Furbank et al, 2020). As the improvement of rice grain yield keeps stagnant in recent years, it has been suggested that pursuing high grain yield by increasing Pn and BM accumulationmay be another effective way to achieve this goal in rice (Zhu et al, 2010; Long et al, 2015). To this end, a few QTLs related to Pn have been identified from cultivated rice and wild rice species (Masumoto et al, 2004; Adachi et al, 2011; Haritha et al, 2017), and some QTLs from common wild rice species,and, have been proven and employed in increasing Pn of cultivated rice (Yeo et al, 1994; Masumoto et al, 2004; Kiran et al, 2013). However, less QTLs and genes are detected in other wild rice species, meaning that it is needed to identify more novel QTLs or genes related to photosynthesis and BM from wild rice apart fromandso as to broaden the genetic diversity of cultivated rice to break the barrier of high grain yield at present status.
In this study, we focused on a population of 152 BILs from a distant cross ofand 9311, and detected 12 novel QTLs related to Pn and BM in, indicating thatcontains abundant genetic resources for Pn improvement. Among them, the,largely contributing to the high Pn, was detected in both Ezhou and Lingshui (Fig. 2 and Table 3), reflecting the great potentiality ofin the improvement of rice for high Pn. Importantly, genomic analysis showed thatwas delimited by L/Bin 8-85 and R/Bin 8-86 in a region with a physical distance of about 68 kb containing only 11 genes referenced to the MH63 genome (Table S1). This region was overlapped with, which encodes a highly conserved chloroplast- localized protein and has positive effects on Pn and BM (Wang et al, 2018). Interestingly, theelite allele substantially improves the nitrogen-use-efficiencyand yield performance under nitrogen-limiting conditions (Wang et al, 2021), and showed significantly high expression in BILs harboringin this study, which was in accordance with the opinion that the expression of genes related to carbon assimilation and nitrogen-use-efficiency are usually coordinated in high- yielding crop plants (Li et al, 2018). If we introgress theinto elite cultivars rice, it may greatly enhance both the Pn and nitrogen-use-efficiency of rice to break the yield barrier in rice (Zhao et al, 2010).
BM is a critical indicator reflecting the intensity of plant Pn to some extent (Yamori et al, 2016). A number of studieshave confirmed that Pnmay providea novel way to increase BM (Richards, 2000; Kim et al, 2018). In this study,for BM andfor DBM fromwere also repeatedly detected in the BIL population in the two sites, indicating these loci play stable effects in different environments. Moreover,andwere overlapped withfrom 9311 to form a cluster/(Fig. 2), which meant that the alleles for high photosynthesis from 9311 and the alleles for BM from wild rice were un-allelic.At present, it has been well-documented that Pn and BM have a significantly positive correlation (Teng et al, 2004; Kiran et al, 2013). If we further pyramid/andtogether, they may show larger effects on the rice yield improvement by simultaneously increasing rice BM accumulation and Pn (Furbank et al, 2019).
detected in this study does not overlap with any other previously identified QTLs (Zhang et al, 2017), and it was delimited to a region between L/Bin 1-143 and R/Bin 1-144 with a physical distance of about 111.4 kb in the MH63 genome (Fig. 3), which contained only 15 functional genes (Table S2). These functions will greatly establish the basis to clone the candidate genes for high Pn and BM accumulation from wild rice, although few valuable genes inhave been cloned and explored in the rice genetic improvement (Fan et al, 2019).
Methods
Plant materials and growth conditions
A set of rice BC2F20BIL population derived from wild riceand the recurrent parent 9311, an elitecultivar, were employed for BM and Pn QTLs analysis (Jin et al,2018). All the parent and 152 BILs were planted in the experimental field of Ezhou (114º35′ E, 30º10′ N, Hubei Province) and Lingshui (110º02′E, 18º48′ N, Hainan Province) in 2020 and 2021, respectively. The average temperature and relative humidity in Lingshui were 27.5ºC and 79.9%, respectively during March and April 2021, and those in Ezhou was 26.5ºC, and (80.46%), respectively during August andSeptember 2020. Fifty plants of each BIL and 9311 were transplanted with a row spacing of 18 cm × 20 cm, and with normal management under natural conditions.
Measurements of Pn, BM and DBM
At the heading stage, field-grown plants were used for detailed quantification of DBM and BM. Pn of fully expanded flag leaves was measured using an infrared gas analyzer (Li-6800, LI-COR, Lincoln, NE, USA) with a leaf chamber fluorescent. For all measurements, the leaf chamber was maintained at the reference CO2concentration of 400 μmol/mol, natural light irradiance of 1500 µmol/(m2·s), relative humidity of 70%–80%, leaf temperature in the gas exchange chamber of 30 ºC, and leaf-to-air vapour pressure deficit of 1.0 kPa. Gas exchange measurements were performed between 9.0 and 11.3 h on clear sunny days. Meanwhile, leaves were held in the leaf chamber until the photosynthetic values were stabile (Chang et al, 2017; Du et al, 2020). To ensure the reliability of the data, all the measurements were conducted under saturated sun light, and at least five individual plants were measured for each line.
After the Pn was measured, the above ground of each plant was harvested and dried immediately in an oven at 85 ºCfor measurement of BM, and five randomly-selected plants were collected for each BIL. The duration from seeding to 20% panicle emerged (start heading) was recorded as heading time. DBM is the dry weight of above ground divided by the heading time.
Data analysis
Analysis of the data of Pn and BM were performed using SPSS software (version25.0, SPSS,Chicago, USA), and the 95% confidence intervals (≤ 0.05) was considered statistically significant. Correlation among four traits was computed at< 0.05 and< 0.01. The graphs were performed by GraphPad Prism 8 (GraphPad Software, San Diego, USA). The images were typesetting by Adobe Illustrator software (version2021, AI, San Jose, USA).
QTL analysis
The methods of DNA extraction, SNP screening, genotyping, bin map and genetic linkage map construction were applied according to Fan et al (2019). Genetic linkage maps of the parents and 152 BIL populations were constructed using QTL IciMapping V4.1 software. Significance levels for thelogarithm of odds (LOD) support intervals were determined by 1000-permutation test corresponding to= 0.05. QTL mapping for Pn and BM were analyzed, and the threshold LOD of adjacent markers was 2.5. The estimated additive effect and phenotypic variation explained by each putative QTL were also calculated in IciMapping V4.1 software based on the population variance. The candidate genes in the QTL region were identified through the database of mh63rs1 (RIGW, http://rice.hzau.edu.cn/rice_rs3).
RNA isolation and qRT-PCR analyses
To validate the detected QTLs for high Pn in the localization interval, RNA was extracted from the flag leaves of the parent and the selected BILs at the heading stage. Total RNA was extracted using TRIzol reagent(Invitrogen, ThermoFisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol and then reverse transcribed using ABScript II cDNA First Strand Synthesis Kit (ABclonal, Wuhan, China) with Oligo d(T)23 primer. qRT-PCR analyses were performed in a CFX connect Real-time PCR detection system (Bio-Rad, California, USA) with 2× Universal SYBR Green Fast qPCR Mix (ABclonal, Wuhan, China). The rice() gene was used as an internal control, and primers used in this assay were provided in Table S3.
ACKNOWLEDGEMENTS
This study was partly granted from the National Natural Science Foundation of China (Grant Nos. U20A2023 and 32172074), the Creative Research Groups of the Natural Science Foundation of Hubei Province, China (Grant No. 2020CFA009), and the earmarked fund for China Agriculture Research System (Grant No. CARS-01-08).
SUPPLEMENTAL DATA
The following materials are available in the online version of this article at http://www.sciencedirect.com/journal/rice-science; http://www.ricescience.org.
Fig. S1. Sequence alignment of candidate genes of.
Table S1. Predicted functional genes atlocus
Table S2. Predicted functional genes at QTL cluster
Table S3. Primers used for qRT-PCR analysis.
Adachi S, Tsuru Y, Nito N, Murata K, Yamamoto T, Ebitani T, Ookawa T, Hirasawa T. 2011. Identification and characterization of genomic regions on chromosomes 4 and 8 that control the rate of photosynthesis in rice leaves., 62(6): 1927‒1938.
Adachi S, Ohkubo S, San N S, Yamamoto T. 2020. Genetic determination for source capacity to support breeding of high- yielding rice ()., 40: 20.
Chang T G, Xin C P, Qu M N, Zhao H L, Song Q F, Zhu X G. 2017. Evaluation of protocols for measuring leaf photosynthetic properties of field-grown rice., 24(1): 1‒9.
Driever S M, Lawson T, Andralojc P J, Raines C A, Parry M A J. 2014. Natural variation in photosynthetic capacity, growth, and yield in 64 field-grown wheat genotypes., 65(17): 4959‒4973.
Du T T, Meng P, Huang J L, Peng S B, Xiong D L. 2020. Fast photosynthesis measurements for phenotyping photosynthetic capacity of rice., 16: 6.
Fan F F, Long W X, Liu M M, Yuan H R, Pan G J, Li N W, Li S Q. 2019. Quantitative trait locus mapping of the combining ability for yield-related traits in wild rice., 67(32): 8766‒8772.
Foulkes M J, Slafer G A, Davies W J, Berry P M, Sylvester-Bradley R, Martre P, Calderini D F, Griffiths S, Reynolds M P. 2011. Raising yield potential of wheat: III. Optimizing partitioning to grain while maintaining lodging resistance., 62(2): 469‒486.
Furbank R T, Jimenez-Berni J A, George-Jaeggli B, Potgieter A B, Deery D M. 2019. Field crop phenomics: Enabling breeding for radiation use efficiency and biomass in cereal crops., 223(4): 1714‒1727.
Furbank R T, Sharwood R, Estavillo G M, Silva-Perez V, Condon A G. 2020. Photons to food: Genetic improvement of cereal crop photosynthesis., 71(7): 2226‒2238.
Godfray H C J, Beddington J R, Crute I R, Haddad L, Lawrence D, Muir J F, Pretty J, Robinson S, Thomas S M, Toulmin C. 2010a. Food security: The challenge of feeding 9 billion people., 327: 812‒818.
Godfray H C J, Crute I R, Haddad L, Lawrence D, Muir J F, Nisbett N, Pretty J, Robinson S, Toulmin C, Whiteley R. 2010b. The future of the global food system., 365: 2769‒2777.
Halder T, Hoque M E, Islam M M, Ali L, Chowdhury A K. 2016. Morphomolecular characterization of local Boro rice (L.) germplasm., 29(2): 1‒9.
Haritha G, Vishnukiran T, Yugandhar P, Sarla N, Subrahmanyam D.2017. Introgressions fromincrease photosynthetic efficiency of KMR3 rice lines., 24(2): 85‒96.
Huang X H, Kurata N, Wei X H, Wang Z X, Wang A H, Zhao Q, Zhao Y, Liu K Y, Lu H Y, Li W J, Guo Y L, Lu Y Q, Zhou C C, Fan D L, Weng Q J, Zhu C R, Huang T, Zhang L, Wang Y C, Feng L, Furuumi H, Kubo T, Miyabayashi T, Yuan X P, Xu Q, Dong G J, Zhan Q L, Li C Y, Fujiyama A, Toyoda A, Lu T T, Feng Q, Qian Q, Li J Y, Han B. 2012. A map of rice genome variation reveals the origin of cultivated rice., 490: 497‒501.
Huo X, Wu S, Zhu Z F, Liu F X, Fu Y C, Cai H W, Sun X Y, Gu P, Xie D X, Tan L B, Sun C Q. 2017.increases grain production in rice., 8(1): 1497.
Jin J, Long W X, Wang L T, Liu X D, Pan G J, Xiang W, Li N W, Li S Q. 2018. QTL mapping of seed vigor of backcross inbred lines derived fromunder artificial aging., 9: 1909.
Kartika K, Sakagami J I, Lakitan B, Yabuta S, Wijaya A, Kadir S, Widuri L I, Siaga E, Nakao Y. 2020. Morpho-physiological responseofto gradual soil drying., 27(1): 67‒74.
Kazama T, Itabashi E, Fujii S, Nakamura T, Toriyama K. 2016. Mitochondriallevels determine pollen abortion in cytoplasmic male sterile rice., 85(6): 707‒716.
Kemanian A R, Stöckle C O, Huggins D R, Viega L M. 2007. A simple method to estimate harvest index in grain crops., 103(3): 208‒216.
Kim Y S, Kim J J, Park S I, Diamond S, Boyd J S, Taton A, Kim I S, Golden J W, Yoon H S. 2018. Expression ofgene improves cellular redox homeostasis and photosynthesis efficiency inPCC 7942., 9: 1848.
Kiran T V, Rao Y V, Subrahmanyam D, Rani N S, Bhadana V P, Rao P R, Voleti S R. 2013. Variation in leaf photosynthetic characteristics in wild rice species., 51(3): 350‒358.
Kovach M J, Sweeney M T, McCouch S R. 2007. New insights intothe history of rice domestication., 23(11): 578‒587.
Li S, Tian Y H, Wu K, Ye Y F, Yu J P, Zhang J Q, Liu Q, Hu M Y, Li H, Tong Y P, Harberd N P, Fu X D. 2018. Modulating plant growth-metabolism coordination for sustainable agriculture., 560: 595‒600.
Long S P, Marshall-Colon A, Zhu X G. 2015. Meeting the global food demand of the future by engineering crop photosynthesis and yield potential., 161(1): 56‒66.
Mann C C. 1999. Genetic engineers aim to soup up crop photosynthesis., 283: 314‒316.
Mao H L, Sun S Y, Yao J L, Wang C R, Yu S B, Xu C G, Li X H, Zhang Q F. 2010. Linking differential domain functions of theGS3 protein to natural variation of grain size in rice., 107(45): 19579‒19584.
Masumoto C, Ishii T, Kataoka S, Hatanaka T, Uchida N. 2004. Enhancement of rice leaf photosynthesis by crossing between cultivated rice,and wild rice species,., 7(3): 252‒259.
Mathan J, Singh A, Jathar V, Ranjan A. 2021. High photosynthesis rate in the selected wild rice is driven by leaf anatomy mediating high Rubisco activity and electron transport rate., 72(20): 7119‒7135.
Monna L, Kitazawa N, Yoshino R, Suzuki J, Masuda H, Maehara Y, Tanji M, Sato M, Nasu S, Minobe Y. 2002. Positional cloning of rice semidwarfing gene,: Rice ‘green revolution gene’ encodes a mutant enzyme involved in gibberellin synthesis., 9: 11‒17.
Omukai S, Arimura S I, Toriyama K, Kazama T. 2021. Disruption of mitochondrial open reading frame 352 partially restores pollen development in cytoplasmic male sterile rice., 187(1): 1‒11.
Ray D K, Mueller N D, West P C, Foley J A. 2013. Yield trends are insufficient to double global crop production by 2050., 8(6): e66428.
Reynolds M, Langridge P. 2016. Physiological breeding., 31: 162‒171.
Richards R A. 2000. Selectable traits to increase crop photosynthesis and yield of grain crops., 51: 447‒458.
South P F, Cavanagh A P, Lopez-Calcagno P E, Raines C A, Ort D R. 2018. Optimizing photorespiration for improved crop productivity., 60(12): 1217‒1230.
Sugimoto K, Takeuchi Y, Ebana K, Miyao A, Hirochika H, Hara N, Ishiyama K, Kobayashi M, Ban Y, Hattori T, Yano M. 2010. Molecular cloning of, a regulator involved in seed dormancy and domestication of rice., 107(13): 5792‒5797.
Sweeney M, McCouch S. 2007. The complex history of the domestication of rice., 100(5): 951‒957.
Takano-Kai N, Jiang H, Kubo T, Sweeney M, Matsumoto T, Kanamori H, Padhukasahasram B, Bustamante C, Yoshimura A, Doi K, McCouch S. 2009. Evolutionary history of, a gene conferring grain length in rice., 182(4): 1323‒1334.
Teng S, Qian Q, Zeng D L, Kunihiro Y, Fujimoto K, Huang D N, Zhu L H. 2004. QTL analysis of leaf photosynthetic rate and related physiological traits in rice (L.)., 135: 1‒7.
Thalapati S, Batchu A K, Neelamraju S, Ramanan R. 2012.gene from a wild rice,improves yield in rice., 12(2): 277‒289.
Wang Q, Nian J Q, Xie X Z, Yu H, Zhang J, Bai J T, Dong G J, Hu J, Bai B, Chen L C, Xie Q J, Feng J, Yang X L, Peng J L, Chen F, Qian Q, Li J Y, Zuo J R. 2018. Genetic variations inmediate grain yield by modulating nitrogen utilization in rice., 9(1): 735.
Wang Q, Su Q M, Nian J Q, Zhang J, Guo M, Dong G J, Hu J, Wang R S, Wei C S, Li G W, Wang W, Guo H S, Lin S Y, Qian W F, Xie X Z, Qian Q, Chen F, Zuo J R. 2021. Thetranscription factor repressesexpression to enhance nitrogen utilization and grain yield in rice., 14(6): 1012‒1023.
Yamori W, Kondo E, Sugiura D, Terashima I, Suzuki Y, Makino A. 2016. Enhanced leaf photosynthesis as a target to increase grain yield: Insights from transgenic rice lines with variable Rieske FeS protein content in the cytochromeb/f complex., 39(1): 80‒87.
Yeo M E, Yeo A R, Flowers T J. 1994. Photosynthesis and photorespiration in the genus., 45(5): 553‒560.
Yu J P, Xiong H Y, Zhu X Y, Zhang H L, Li H H, Miao J L, Wang W S, Tang Z S, Zhang Z Y, Yao G X, Zhang Q, Pan Y H, Wang X, Rashid M A R, Li J J, Gao Y M, Li Z K, Yang W C, Fu X D, Li Z C. 2017.contributing to rice grain length and yield was mined by Ho-LAMap., 15(1): 28.
Zhang S H, He X Y, Zhao J L, Cheng Y S, Xie Z M, Chen Y H, Yang T F, Dong J F, Wang X F, Liu Q, Liu W, Mao X X, Fu H, Chen Z M, Liao Y P, Liu B. 2017. Identification and validation of a novel major QTL for harvest index in rice (L.)., 10(1): 44.
Zhao M, Ding Z, Lafitte R, Sacks E, Dimayuga G, Holt D. 2010. Photosynthetic characteristics inspecies., 48(2): 234‒240.
Zhu X G, Long S P, Ort D R. 2008. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass?, 19(2): 153‒159.
Zhu X G, Long S P, Ort D R. 2010. Improving photosynthetic efficiency for greater yield., 61: 235‒261.
22 October 2021;
26 January 2022
Copyright © 2022, China National Rice Research Institute. Hosting by Elsevier B V
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/)
Peer review under responsibility of China National Rice Research Institute
http://dx.doi.org/10.1016/j.rsci.2022.01.011
LiShaoqing (shaoqingli@whu.edu.cn); Peng Xiaojue (xiaojuepeng@ncu.edu.cn)
(Managing Editor: Wang Caihong)
杂志排行

Rice Science的其它文章
- Transcriptome Analysis Provides Novel Insights into Salt Stress Response in Two Egyptian Rice Varieties with Different Tolerance Levels
- QTL Mapping for Plant Height Using Introgression Lines Derived from Zhonghui 8015 and Wild Rice (Oryza rufipogon)
- Genetic Dissection of Quantitative Trait Loci for Panicle Traits and Heat Tolerance by High-Density Bin Map in Rice
- Improvement of Rice Production under Drought Conditions in West Africa:Application of QTLs in Breeding for Drought Resistance
- Characterization of a Novel Weak Allele of RGA1/D1 and Its Potential Application in Rice Breeding
- CRISPR/Cas9-Targeted Knockout of Rice Susceptibility Genes OsDjA2 and OsERF104 Reveals Alternative Sources of Resistance to Pyricularia oryzae