Characterization of a Novel Weak Allele of RGA1/D1 and Its Potential Application in Rice Breeding
2022-10-25LiuYantongLiTingJiangZhishuZengChuihaiHeRongQiuJiaoLinXiaoliPengLimeiSongYongpingZhouDahuCaiYicongZhuChanglanFuJunruHeHaohuaXuJie
Liu Yantong, Li Ting, Jiang Zhishu, Zeng Chuihai, He Rong, Qiu Jiao, Lin Xiaoli, Peng Limei, Song Yongping, Zhou Dahu, Cai Yicong, Zhu Changlan, Fu Junru, He Haohua, Xu Jie
Research Paper
Characterization of a Novel Weak Allele ofand Its Potential Application in Rice Breeding
Liu Yantong#, Li Ting#, Jiang Zhishu#, Zeng Chuihai, He Rong, Qiu Jiao, Lin Xiaoli, Peng Limei, Song Yongping, Zhou Dahu, Cai Yicong, Zhu Changlan, Fu Junru, He Haohua, Xu Jie
(These authors contributed equally to this work)
Semi-dwarfing improves the lodging resistance and yield of rice, and the vast majority of modern rice varieties harbor theallele to decrease plant height, resulting in reduced genetic diversity and negative agronomic traits. Thus, exploring alternative sources of dwarfism is imperative for rice breeding. Here, we identified a novelallele,, from a localvariety Xiaolixiang (XLX) using a map-based cloning approach. Compared with other rice varieties,in XLX contained a unique single nucleotide polymorphism that resulted in an additional transcript and reduced functionaltranscript level. Thefrom Nipponbare was introduced into XLX to estimate the value ofin rice breeding. Compared with transgenic XLX plants (XLX), XLX exhibited reduced plant height, increased stem strength, lower reactive oxygen species accumulation, delayed senescence, stronger photosynthesis, higher grain yield and quality (including external, milling and nutritional qualities), and enhanced resistance to drought and. Therefore, we proposed that theallele has potential as an excellent dwarfism resource for rice breeding.
rice;weak allele;; dwarf germplasm resource; yield; grain quality; drought resistance; sheath blight
As a highly important cereal crop, rice () is a staple food for more than half of the world’s population (Yuan, 2014). The last century witnessed two major breakthroughs in rice breeding, the green revolution in the 1960s and the application of hybrid rice in the 1970s. The green revolution was triggered by the introduction of() gene and the creation of semi-dwarf varieties (Khush, 2001). Dwarfism is a very important agronomic trait in rice breeding programs, because it can not only enhance lodging resistance and harvest index, but also has better responses to fertilizer (Monna et al, 2002; Hedden, 2003).
Since then, rice breeding for dwarf varieties has attracted intensive interest from breeders, and more than 80 dwarf mutants have been characterized according to the China Rice Data Center (https://www.ricedata. cn/gene/). There are various reasons for plant dwarfing,including phytohormone-related and hormone-independentpathways. Several phytohormones including gibberellins (GAs), brassinosteroids (BRs), strigolactones (SLs), 3-indoleacetic acid, abscisic acid and ethylene influence crop height (Ashikari et al, 1999; Hong et al, 2002; Sazuka et al, 2009; Yaish et al, 2010; Qi et al, 2011; Alder et al, 2012). Most other dwarf genes that act in non-hormone manner are involved in cell wall development, cytosolic glutamine synthetic pathway, RNA editing, cell division, ubiquitin-proteasome pathway and fatty acid metabolism (Liu et al, 2018). Dwarfing-related genes involved in GAs, BRs and SLs were reviewed by Liu et al (2018) and Ferrero- Serrano et al (2019), including,,(),,(),(),,,() andin GA biosynthesis and homeostasis;,,,(),,,andin GA signaling;,(),(/D11),CYP90D3,(),(D2),(),,andin BR biosynthesis and degradation;(),(),,,,,(),(),(),,,(),() and() in BR signaling;,and() in SL biosynthesis; and,,(),andin SL signaling.
The plant height has been decreased in most modern rice varieties. Unexpectedly,seems to be the sole dwarf source in rice breeding, and 90% of rice varieties still harbor different alleles of the samegene that are loss-of-function or weak-functional types (Sha et al, 2022).encodes a GA20-oxidase, and theallele reduces bioactive GA abundance and increases the accumulation of DELLA protein (Asano et al, 2011). Despite the mutants ofeffectively generating semi-dwarfism and improving lodging resistance and yield (Hedden, 2003), the widespread use of theallele reduces the genetic diversity of rice varieties, andit has some negative effects. Several disadvantageous traits have reportedly been linked to the introduction of theallele, such as reduced spikelet fertility in response to low temperature (Murai et al, 1991), drought sensitivity (Vikram et al, 2015) and low nitrogen use-efficiency (NUE) (Li et al, 2018). Therefore, using alternative dwarf sources in breeding programs would help to correct such secondary negative traits (Wang et al, 2021). Of the dwarf sources previously identified, mutations in/may have potential applications to rice breeding improvement.encodes the α subunit of hetero- trimeric G proteins, which function as messengers in numerous signal transduction pathways (Gilman, 1987). Null mutations ofin rice reduce plant response to both GA and BR (Ueguchi-Tanaka et al, 2000; Oki et al, 2009). The identifiedmutants exhibit abnormal morphologies, such as dwarfism (Ashikari et al, 1999), increased reactive oxygen species (ROS) scavenging ability and stress tolerance (Suharsono et al, 2002; Peng et al, 2019; Cui et al, 2020). For instance, the survival rates ofplants are much higher than those of wild type after drought, chilling and salinity treatments (Urano et al, 2014; Ferrero-Serrano and Assmann, 2016; Peng et al, 2019; Cui et al, 2020).mutants exhibit a highly reduced hypersensitive response to several rice blast fungi and ROS productions, and the expression of defense genes induced by sphingolipid elicitors is reduced or delayed (Suharsono et al, 2002; Assmann, 2005).
Althoughand some other dwarf mutants exhibit beneficial characteristics that are perfect complementation for, theallele still seems to be irreplaceable. The reason is that most dwarf alleles are obtained from mutants which often display pleiotropic phenotypes, such as malformed leaves and panicles, poor sterility, small grains, and low biomasses and yields (Hong et al, 2002; Sazuka et al, 2009; Asano et al, 2010; Hu et al, 2010). Therefore, it is important to continue search for novel semi-dwarf genes. Exploring weak alleles of dwarfing-related genes (especially) from rice varieties, rather than mutants, is the most effective way to obtain valuable dwarf resources and increase genetic diversity in breeding programs. In this study, we isolated a novelallele () from a localrice variety Xiaolixiang (XLX). This weak allele causes a lower expression level of the functionaltranscript. Compared with XLX transformed with normal, wild type XLX showed improved yield and stress tolerance in addition to dwarfism. Therefore,could be a perfect germplasm resource for rice breeding.
RESULTS
Phenotype of XLX
Among 500 local rice varieties, XLX initially garnered particular attention because of its dense and erect panicles and small round seeds (dep-srs)phenotype (Fig. 1). Compared with othervarieties (Table 1), XLX had more effective panicle number per plant, shorter panicle length and grain length, and lower 1000-grain weight (TGW). The grain length-to-width ratio of XLX was less than 2.0, and its TGW was only 16.3 g. There were significantly more secondary branch and grain numbers per spikelet in XLX than in Taichung Native 1 (TN1) and Huazhan (HZ), but moderately fewer than Changhui T025 (CHT025). Interestingly, the plant height of XLX seemed to be relatively normal, even a little higher than other varieties, but the F1plant heights of XLX with other varieties (including TN1, HZ and CHT025) were astronomically high (over 160 cm). These results suggested that XLX harbors a specific recessive semi- dwarf gene that is dominant in other varieties. Meanwhile, XLX might have another positive regulator of plant height that is nonfunctional or weak-functional in other varieties.

Fig. 1. Phenotypic characterization of XLX, HZ, CHT025 and their F1plants.
A and B, Phenotypes of XLX, HZ, CHT025 and their F1plants at the filling stage. Scale bars, 20 cm.
C, Grain shapes of XLX, HZ, CHT025 and their F1s. Scale bar, 5 mm.
XLX, Xiaolixiang; HZ, Huazhan; CHT025, Changhui T025.
Fine mapping of DEP-SRS gene
XLX was initially used to isolate the gene that causes thedep-srsphenotype. A map-based cloning approach was utilized to identify thegene. Totally, 372 plants with the dep-srs phenotype were selected from the F2population (a total of 1 581 plants), which was generated by crossing XLX with TN1. The ratio of individuals with a normal phenotype to those with a dep-srs phenotype fitted well with the expected value of 3:1 (χ2= 1.824,= 0.18). The F2populations from the crosses of XLX with twovarieties (CHT025 and HZ) were used to confirm the segregation ratio. The segregation ratio in the both populations showed a good fit to 3:1 (Table S1). The results indicated that the dep-srs phenotype is controlled by a single recessive gene. Seventy-three simple sequence repeat markers were selected by screening polymorphisms between XLX and TN1, and were then used for linkage analysis. Subsequently, thegene was mapped to a region between markers RM18410 and RM18448 on chromosome 5 (Fig. 2-A). A series of PCR-based markers were developed and detected for fine mapping ofgene (Table S2). Thegene was further narrowed to an interval of about 125 kb between markers M4 and M5, which contains 14 predicted open reading fragments (ORFs) (Fig. 2-A).
We found thatwas located in this region. As allmutants have a dep-srsphenotype that is similar to XLX,was speculated to be the candidate forin XLX. Then,from XLX was sequenced. To avoid the interference by sequence diversity that was unconnected with the dep-srs phenotype, thegene of two othervarieties, TN1 and Nanjing 06 (NJ06), and twovarieties, Nipponbare (NPB) and Wuyunjing 7 (WYJ7), was also sequenced. Sequence alignment indicated thatthesequences in the twovarieties were exactly the same, and those in TN1 and NJ06 were also the same. Moreover, there were 36 nucleotide diversities in(Table S3), almost all of which resided in the promoters or introns, except four synonymous single nucleotide polymorphisms (SNPs) in exons. Compared with other varieties, thesequence in XLX was very close to those in the tworice varieties, except for two unique divergences, both of which were in introns, and the divergence (A to G) in the first intron produced a new splice site (Fig. 2-B and Table S3). The transcripts ofin XLX, NPB and TN1 were sequenced, and the divergence did cause alternative splicing. There were twotranscripts in XLX: one was the same with that in NPB and TN1, and the other added 15 bp at the 5′-teminal of the second exon. The additional sequence contained a stop code that led to a null RGA1 (Fig. 2-C). Unsurprisingly, the levels of thetranscript in XLX were much lower than those in NPB and TN1 (Fig. 2-D). These results suggested that the unique SNP (A/G) incould be responsible for dep-srs phenotype in XLX.
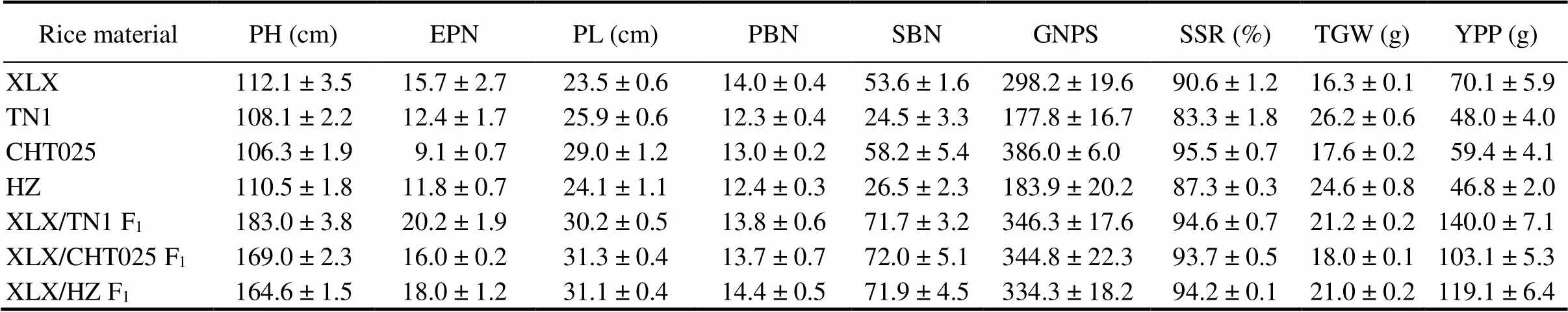
Table 1. Agronomic characteristics of XLX and three common varieties as well as their F1 plants.
Data are Mean ± SD (= 8). XLX, Xiaolixiang; TN1, Taichung Native 1; CHT025, Changhui T025; HZ, Huazhan; PH, Plant height; EPN, Effective panicle number per plant; PL, Panicle length; PBN, Primary branch number per plant; SBN, Secondary branch number per plant; GNPS, Grain number per spikelet; SSR, Seed-setting rate; TGW, 1000-grain weight; YPP, Yield per plant.
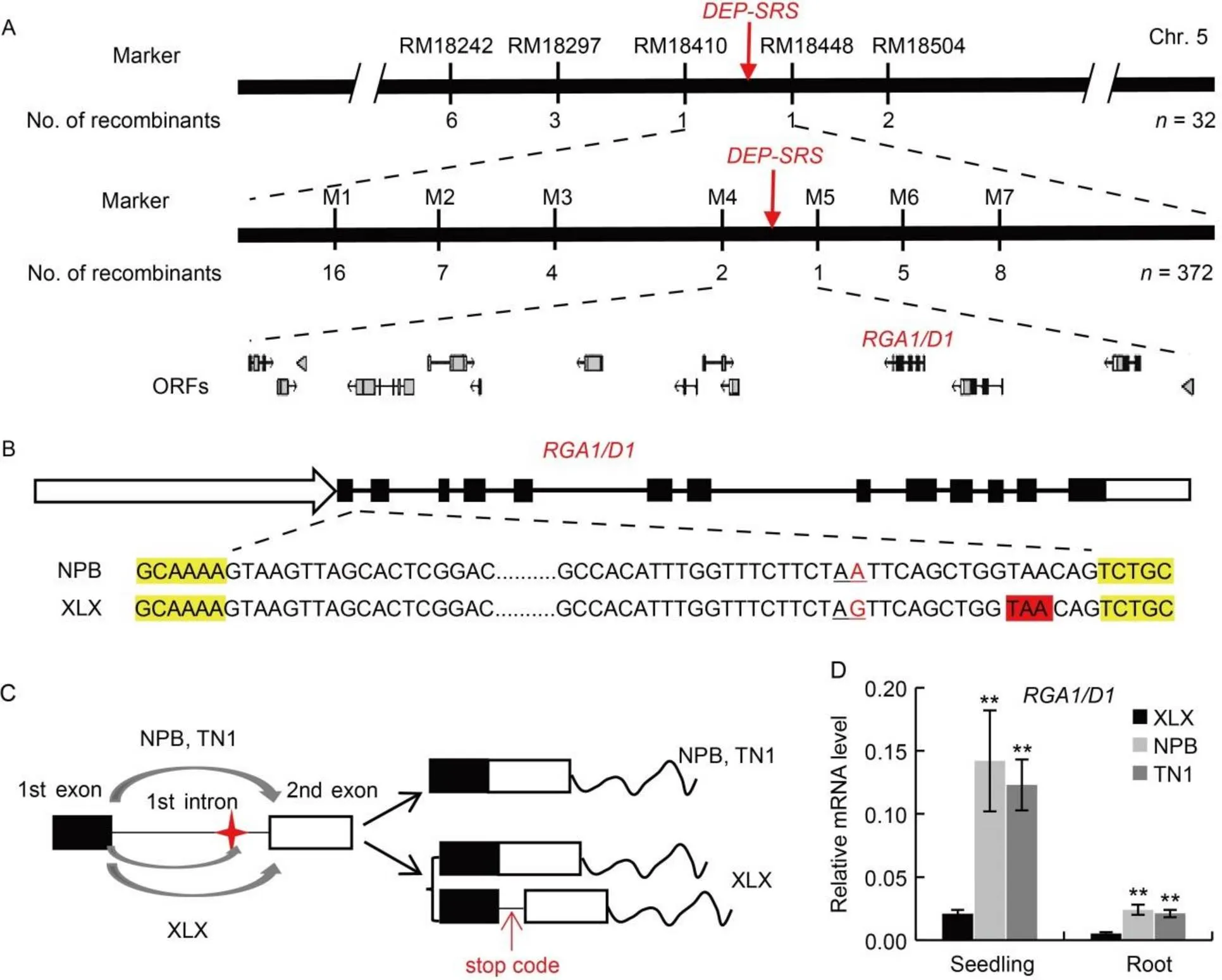
Fig. 2. Map-based cloning ofgene.
A, Fine mapping ofgeneThe gene was mapped to the interval between molecular markers RM18410 and RM18448 on chromosome (Chr.) 5 and further delimited to a 125-kb genomic region between markers M4 and M5 containing 14 open reading fragments (ORFs). Numbers below the markers indicate the number of recombinants.
B, The first intron sequences of XLX and NPB. Black boxes indicate exons, black lines indicate introns, white box with arrow indicates promoter, and white box without arrow indicate 3′-UTR. Letters with yellow background indicate exon sequences, while those with red background indicate a stop code in the new transcript. Red letter with underline indicates the divergent sequence and black letter with underline indicates the splice site.
C, The divergence (A to G) in the first intron produced a new splice site in XLX. Red star indicates the SNP, and red arrow indicates a stop code in the new transcript.
D, Relative expression levels of functional/transcripts. The two-week-old seedlings of XLX, NPB and TN1 were collected for qRT-PCR, and thegene was used as a control. Mean and SD values were obtained from three biological replicates. **,< 0.01 (the Student’s-test).
XLX, Xiaolixiang; NPB, Nipponbare; TN1, Taichung Native 1; SNP, Single nucleotide polymorphism.
Functional complementation of DEP-SRS
A genetic complementation experiment was used to confirm that the weakallele in XLX was responsible for the dep-srsphenotype. The full length ofDNA fragment (6 640 bp, including the 2-kb upstream and 1-kb downstream sequences) was amplified from NPB and inserted into a binary vector pCAMBIA1300 (pRGA1-COM). The constructed vector was introduced into XLX, and 32 transgenic plants were obtained. Compared with the wild type XLX, the transformed plants were higher (both at the seedling and maturity stages) (Fig. 3-A, -B and -F), and had longer panicles and grains (Fig. 3-C, -D, -G and -H). The increased rates for plant height, panicle length and grain length were 41.8% (Fig. 3-F), 29.2% (Fig. 3-G) and 32.8% (Fig. 3-H), respectively. Due to the longer grain length and similar grain width (Fig. 3-D, -E, -H and -I), the grain length-to-width ratio was larger in the transformed plants (Fig. 3-J). In addition, the erect panicle of XLX was converted to the curved panicle in the XLX transformed plants with functional(XLX)(Fig. 3-A). These results demonstrated thatwas indeed responsible for the dep-srs phenotype. In the remainder of this paper,orRGA1was used as the gene name, and XLXwas used to represent transgenic XLX plants.
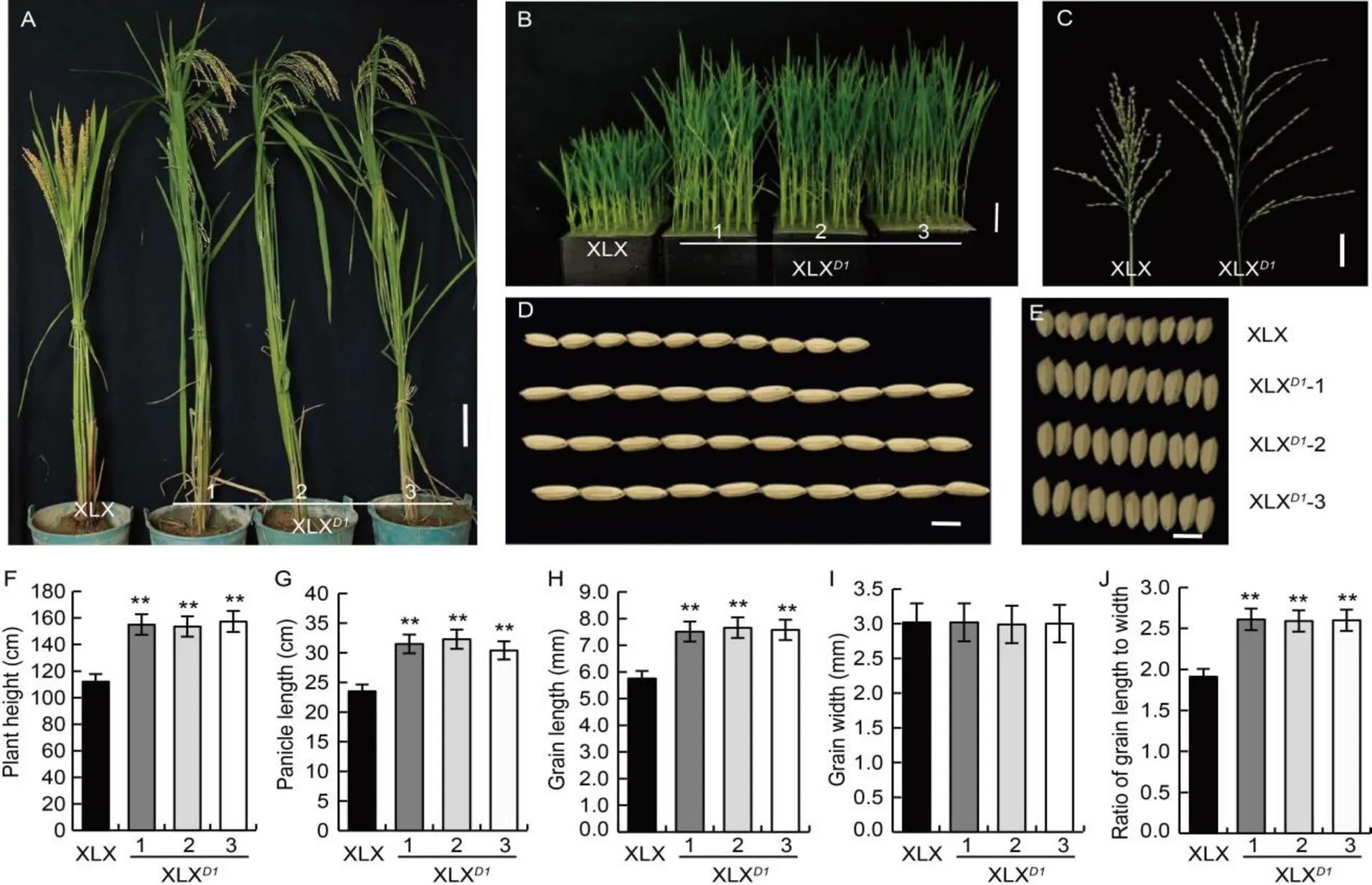
Fig. 3. Phenotypes of complementation transgenic lines of Xiaolixiang (XLX).
A, Morphology at the maturity stage. Scale bar, 10 cm. B, Morphology at the seedling stage. Scale bar, 5 cm. C, Panicle type. Scale bar, 5 cm. D and E, Grain shape. Scale bars, 5 mm. F, Plant height. G, Panicle length. H, Grain length. I, Grain width. J, Ratio of grain length to width.
Mean and SD values were obtained from three biological replicates. **,< 0.01 (the Student’s-test).
d1-w strengthens lodging resistance
In comparison with XLX plants, the lengths of the different internodes of XLXwere all increased (Fig. 4-A and -B). As the culm traits and lodging resistance are closely linked (Liu and Li, 2016), the culm strength was tested at the early- and late-filling stages (Fig. 4-C and Table S4). The strength of each internode of XLX was stronger than that of XLX. At the early-filling stage, the strengths of the upper four internodes in XLX were 6.8, 16.7, 25.6 and 35.1 N, respectively, while those of XLXwere less than or equal to 0.0, 10.7, 19.0 and 25.8 N, respectively. At the late-filling stage, these changed to 7.1, 18.9, 29.8 and 37.5 N in XLX and 0.0, 9.7, 13.8 and 23.5 N in XLX, respectively. Culm strength gradually declined during filling in the transgenic plants, possibly as a result of senescence at the later growth stage. Interestingly, XLX showed the opposite trend, with greater strength at the late-filling stage, which might due to delayed aging caused by.
d1-w enhances photosynthesis in rice
Leaf development was strongly affected by. The leaves of XLX were very short and wide, with dark green at the filling stage, while XLXleaves were long, narrow and pale green (Fig. 5-A). Both the flag leaves and secondary leaves of XLX were 25%‒30% shorter, and about 30% wider than those of XLX(Fig. 5-B and -C). The chlorophyll and carotenoid contents in the flag leaves of XLX were higher at the filling stage by 25%‒30% than those in the XLXplants (Fig. 5-D). The net photosynthetic rate and stomatal conductance were higher at both the heading and filling stages in XLX compared with XLX(Fig. 5-E and -F). Moreover, the net photosynthetic rate in XLX remained at a high level, even at the filling stage, at about 10.0 μmol/(m2∙s), which was nearly double over that in XLX. The optimal/maximal quantum yield of PSII (v/m) was also larger in the wild type XLX than that in XLX(Fig. 5-G), indicating thatenhanced the plants’ maximum potential photosynthetic ability. The transcript levels of photosynthesis-associatedgenes (i.e. light-harvesting chlorophyll-binding protein genesand) were also detected in the XLX and XLXplants, andandwere strongly up-regulated in XLX (Fig. 5-H and -I). In summary, theallele resulted in higher levels of photosynthesis in XLX than in XLXplants.
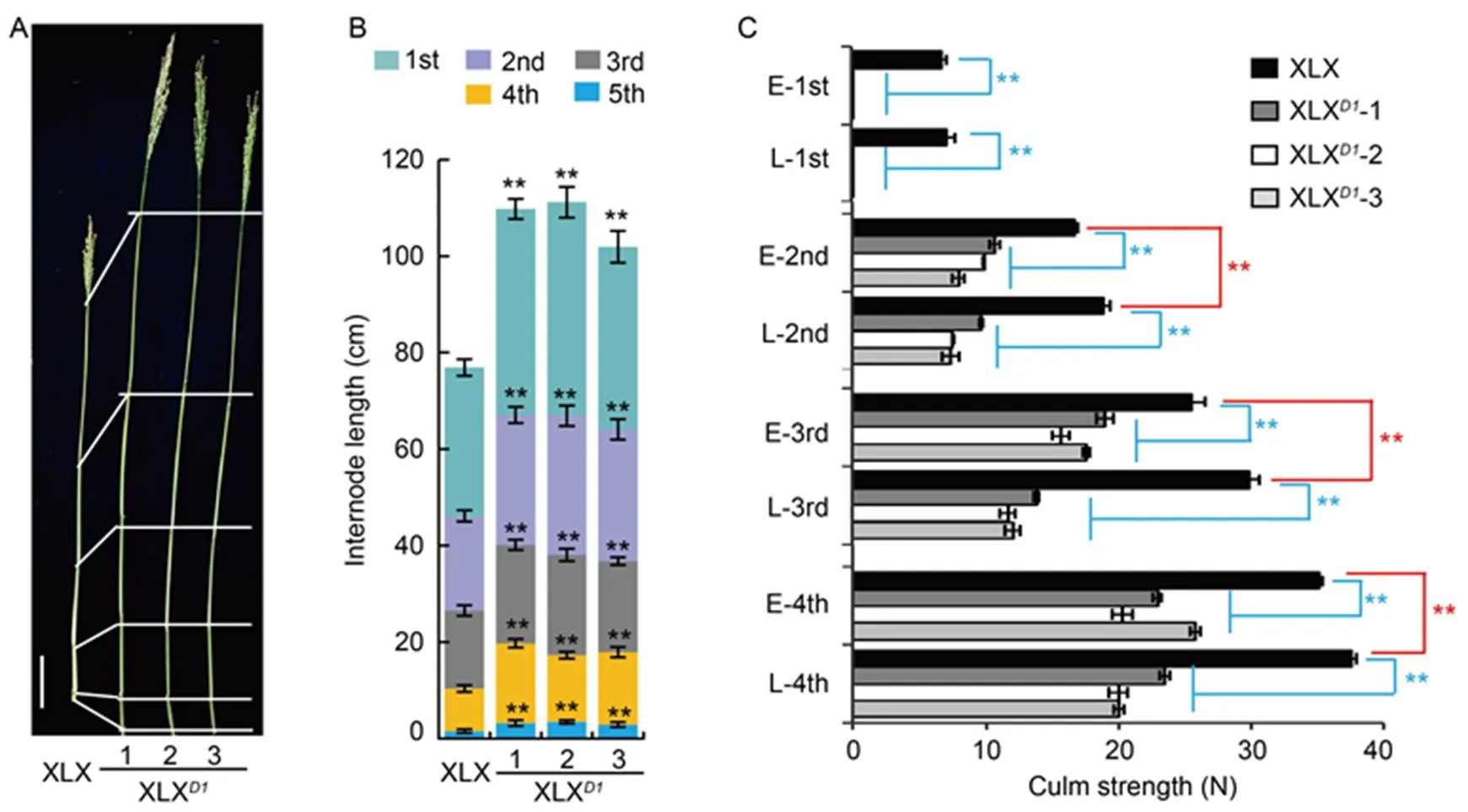
Fig. 4. Lodging resistance of Xiaolixiang (XLX) and its transgenic XLXplants.
A, Morphologies of culm and panicles. Scale bar, 10 cm. B, Internode length. C, Culm strengths of internodes at the early (E)- and late (L)-filling stages.
1st to 5th indicate the first internode to the last internode. Mean and SD values were obtained from three biological replicates. **,< 0.01 (the Student’s-test).
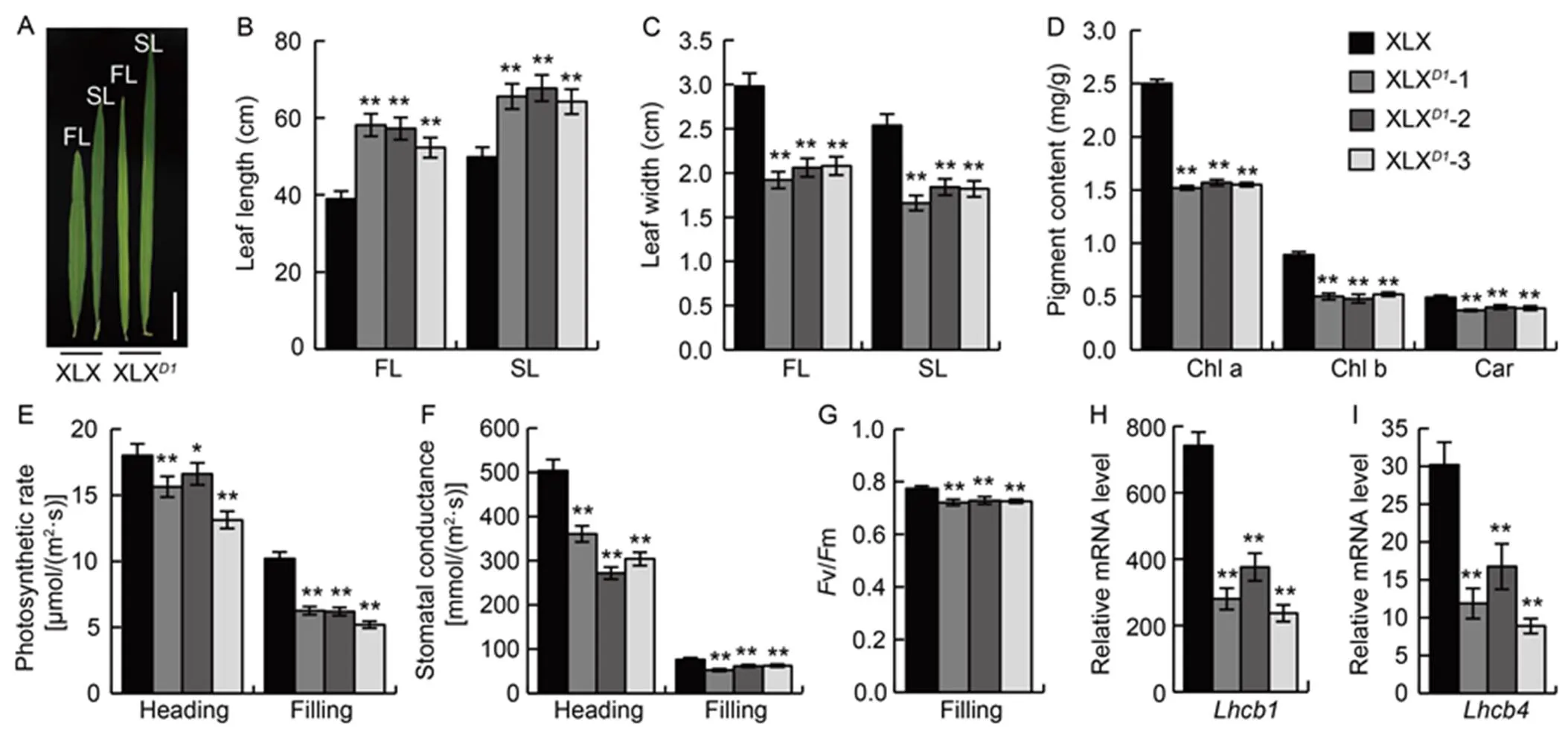
Fig. 5. Photosynthesis of Xiaolixiang (XLX) and its transgenic XLXplants.
A, Morphologies of flag leaf (FL) and secondary leaf (SL) at the filling stage. Scale bar, 10 cm. B, Leaf lengths of FL and SL at the filling stage. C, Leaf widths of FL and SL at the filling stage. D, Pigment content of FL at the filling stage. Chl a, Chlorophyll a; Chl b, Chlorophyll b; Car, Carotenoid. E and F, Photosynthetic rate (E) and stomatal conductance (F) of FL at the heading and filling stages. G, Optimal/maximal quantum yield of PS II (v/m) of FL at the filling stage. H and I, Relative expression levels ofandin FL at the filling stage.
Mean and SD values were obtained from three biological replicates.*,< 0.05; **,< 0.01 (the Student’s-test).
d1-w represses ROS accumulation and delays ageing
ROS accumulation is a key factor resulting in leaf senescence, and RGA1 plays an essential role in ROS accumulation. The H2O2and O2·̄ in flag leaves at the heading and filling stages were detected by diamino- benzidine (DAB) and nitroblue tetrazolium (NBT) staining (Fig. 6). More DAB and NBT signals appeared in the leaves of XLXplants at the two stages, and there were greater accumulations of both signals at the filling stage, while the leaves from XLX showed only few staining spots at the two stages (Fig. 6-A). The transcript levels of ROS-scavenging related genes were tested, and the expression levels of,,andin XLXwere similar to those in XLX, while theexpressionwas obviouslyincreased in XLX(Fig. 6-B). These results suggested that the lower ROS accumulation in XLX was a result of lower ROS production rather than stronger ROS- scavenging activity. Two marker genes of leaf senescence,and, were significantly up- regulated in the XLXplants (Fig. 6-B), indicating thatdelayed leaf senescence of XLX at the filling stage, which resulted in stronger photosynthesis.
d1-w improves grain yield and quality
Grain yield and quality are the most valued traits in rice breeding, and the photosynthetic rate during the grain-filling stage is an important physiological factor that affects grain yield and quality (Feng et al, 2013). A comparative analysis of the agronomic traits of XLX and functionaltransgenic plants was performed to evaluate the application values of(Table 2). The results showed thatincreased the panicle number by about five effective panicles per plant. Both the numbers of primary and secondary branches were increased in theplants, leading to more grain number per spikelet (about 298.2, compared with no more than 200.0 in the XLXplants). The seed-setting rate was mildly improved by theallele. However,reduced the grain length, as well as TGW (only 16.3 g, the reduction reached 23.4%). Combining all yield components, the theoretical yield per plant of XLX (70.1 g) was much higher than that of XLX(about 37.7 g).
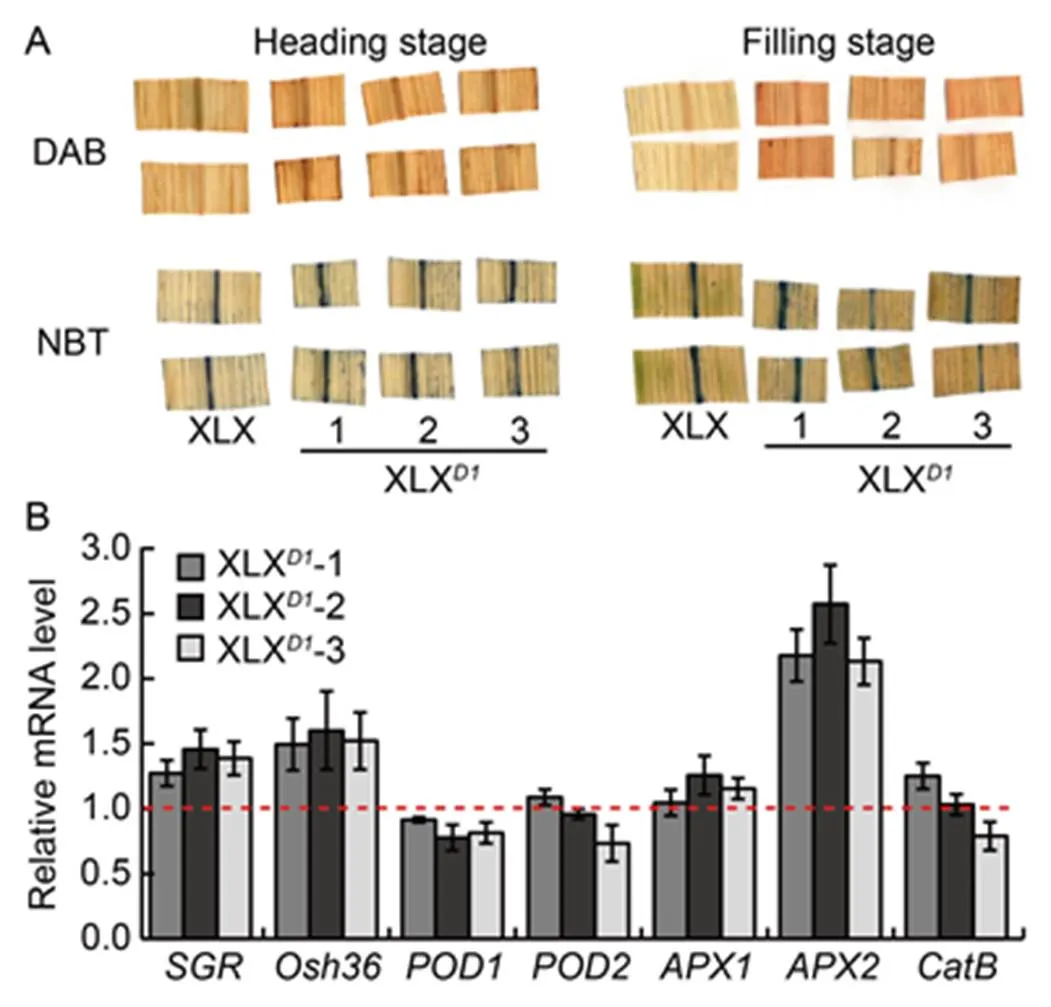
Fig. 6. Delayed leaf senescence in Xiaolixiang (XLX).
A, Diaminobenzidine (DAB) staining and nitroblue tetrazolium (NBT) staining of leaves at the heading and filling stages.
B, Changes in transcript levels of senescence-associated genes and ROS-scavenge enzyme genes in the leaves of XLX and XLXplants at the filling stage. The flag leaves of XLX and XLXplants at the filling stage were used for qRT-PCR. Two senescence-induced genes,() and(); catalase gene(); two peroxidase genes,() and(); two ascorbate peroxidase genes,() and() were detected, and thegene was used as a control.Mean and SD values were obtained from three biological replicates. The red line indicates the transcript level of these genes in XLX.
The grain qualities of XLX and XLXplants were also investigated. Both chalky rice rate and chalkiness degree were much lower in XLX, which were only 8.76% and 0.87%, while those were more than 26.99% and 2.91% in XLX, respectively (Fig. 7-A to -D). The average head rice rate was about 56.41% in XLX, whereas it reached up to 66.33% in XLX (Fig. 7-E). However, the milled rice rate was not significantly different between XLX and XLX, and the brown rice rate was slightly lower in XLX (Fig. 7-E). Moreover, the protein and nitrogen contents of the XLX grains were about 20% and 17% higher than those of XLX(Fig. 7-F and -G). These results indicated that the weak allele ofin XLX greatly improved both grain yield and grain quality, including external quality, milling quality and nutritional quality.
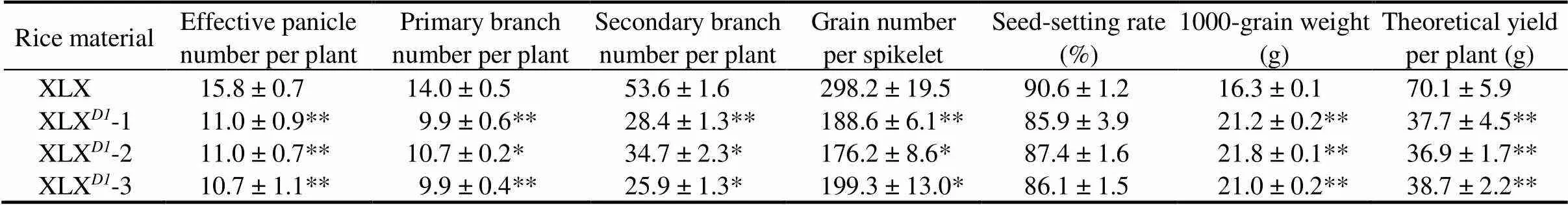
Table 2. Agronomic characteristics of Xiaolixiang (XLX) and its transgenic XLXD1 plants.
Data are Mean ± SD (= 8). *,< 0.05; **,< 0.01 (the Student’s-test).
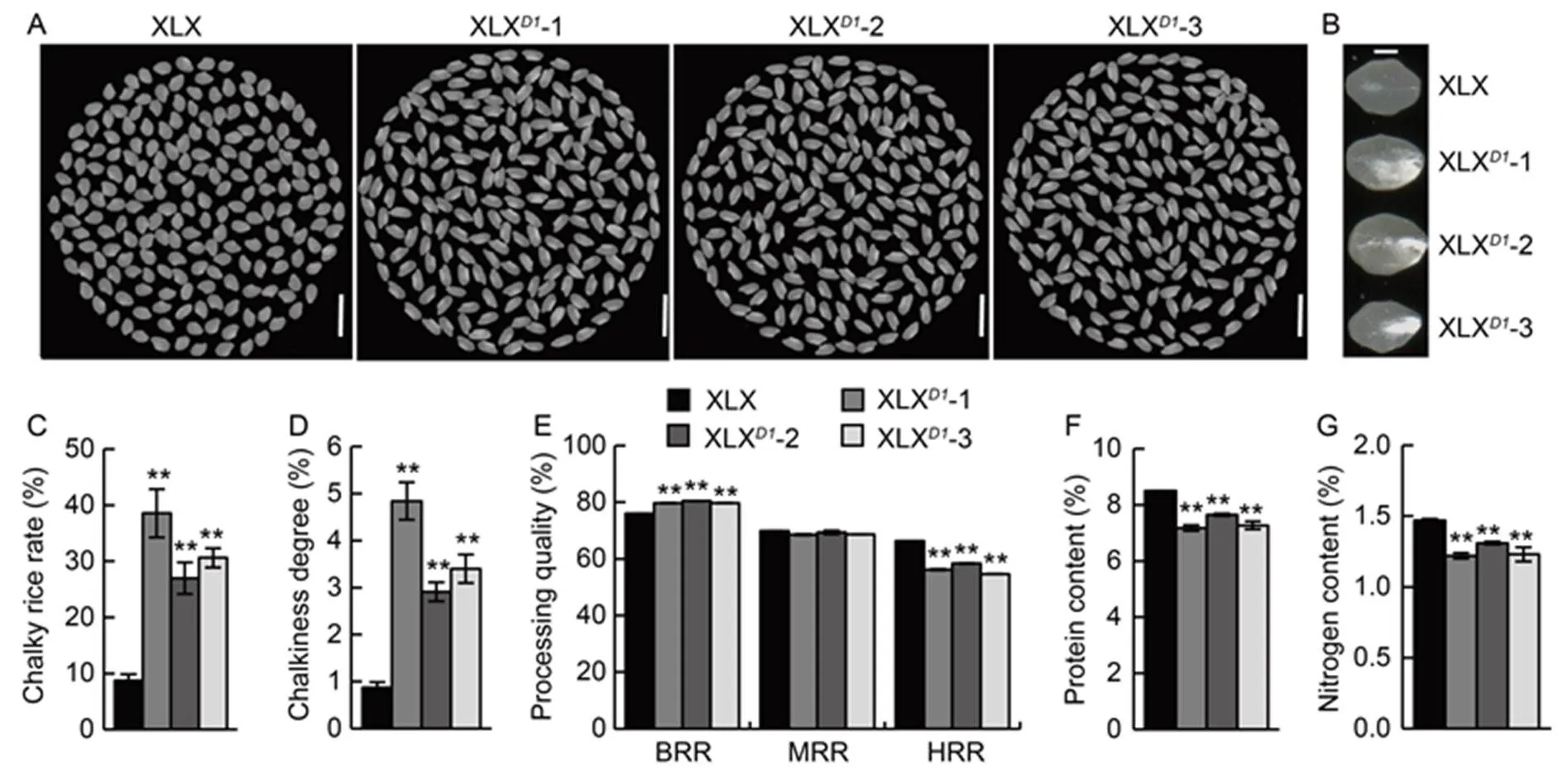
Fig. 7. Grain quality of Xiaolixiang (XLX) and its transgenic XLXplants.
A, Head rice of XLX and its transgenic XLXlines (XLX-1, -2 and -3). Scale bars, 1 cm. B, Chalk sizes of XLX and XLX. Scale bar, 1 mm. C, Chalky rice rates of XLX and XLX. D, Chalkiness degrees of XLX and XLX. E, Processing qualities of XLX and XLX, including brown rice rate (BRR), milled rice rate (MRR) and head rice rate (HRR). F and G, Protein contents (F) and nitrogen contents (G) of XLX and XLXhead rice.
Mean and SD values were obtained from three biological replicates. **,< 0.01 (the Student’s-test).
d1-w raises stress tolerance
As mostmutants show high tolerance to several biotic and abiotic stresses (Urano et al, 2014; Ferrero- Serrano and Assmann, 2016; Peng et al, 2019; Cui et al,2020), two XLXlines (XLX-2 and XLX-3) were used to investigate the effects ofon drought and sheath blight fungal resistances. In the paddy field, XLX had a leaf temperature of 33.6 ºC, while that of XLXplants was about 34.9 ºC (Fig. 8-A to -E). The lower leaf temperature also confirmed the stronger drought tolerance of XLX. Three-leaf seedlings of XLX and XLXplants were treated with 20% polyethylene glycol (PEG) for 13 d. At this point, most XLX plants still had green tissues, while all XLXplants were withered (Fig. 8-F and -G). The survival rates of XLX, XLX-2 and XLX-3 were 85.8%, 6.3% and 2.1%, respectively (Fig. 8-H). When the seedlings were inoculated with, XLXplants exhibited increased susceptibility to infection compared with XLX, which was indicated by much higher hyphal growth rate in XLXplants (Fig. 8-I and -J). The average sheath blight score of XLX was 2.5, while those of XLXlines were significantly higher, with XLX-2 and XLX-3 scoring 4.3 and 5.5, respectively. More than half of XLXseedlings scored over 4.0, whereas most XLX seedlings scored less than 2.0, and a significant portion of XLX had no lesions (Table S5). The leaves harvested from XLX and XLXplants at the tillering stage were also inoculated with, and XLX-2 and XLX-3 showed more severe symptoms than XLX, which had very few disease spots (Fig. 8-K). These results demonstrated thatincreased rice plant resistance to drought and sheath blight.
DISCUSSION
Most rice dwarf mutants are severely dwarfing, resulting in abnormal development and unfavorable phenotypes. For example,/mutants show various degrees of dwarfism, curled leaves, small and wrinkled seeds, and low biomasses (Hu et al, 2010). Themutant has very low fertility (Chen et al, 2019), and() is sensitive to cold, salinity and drought (Liu et al, 2020). It is extremely difficult to identify applicable semi-dwarf resources in mutants that are likely to be workable in rice varieties. In this study, we obtained a newallele () from a local variety XLX.
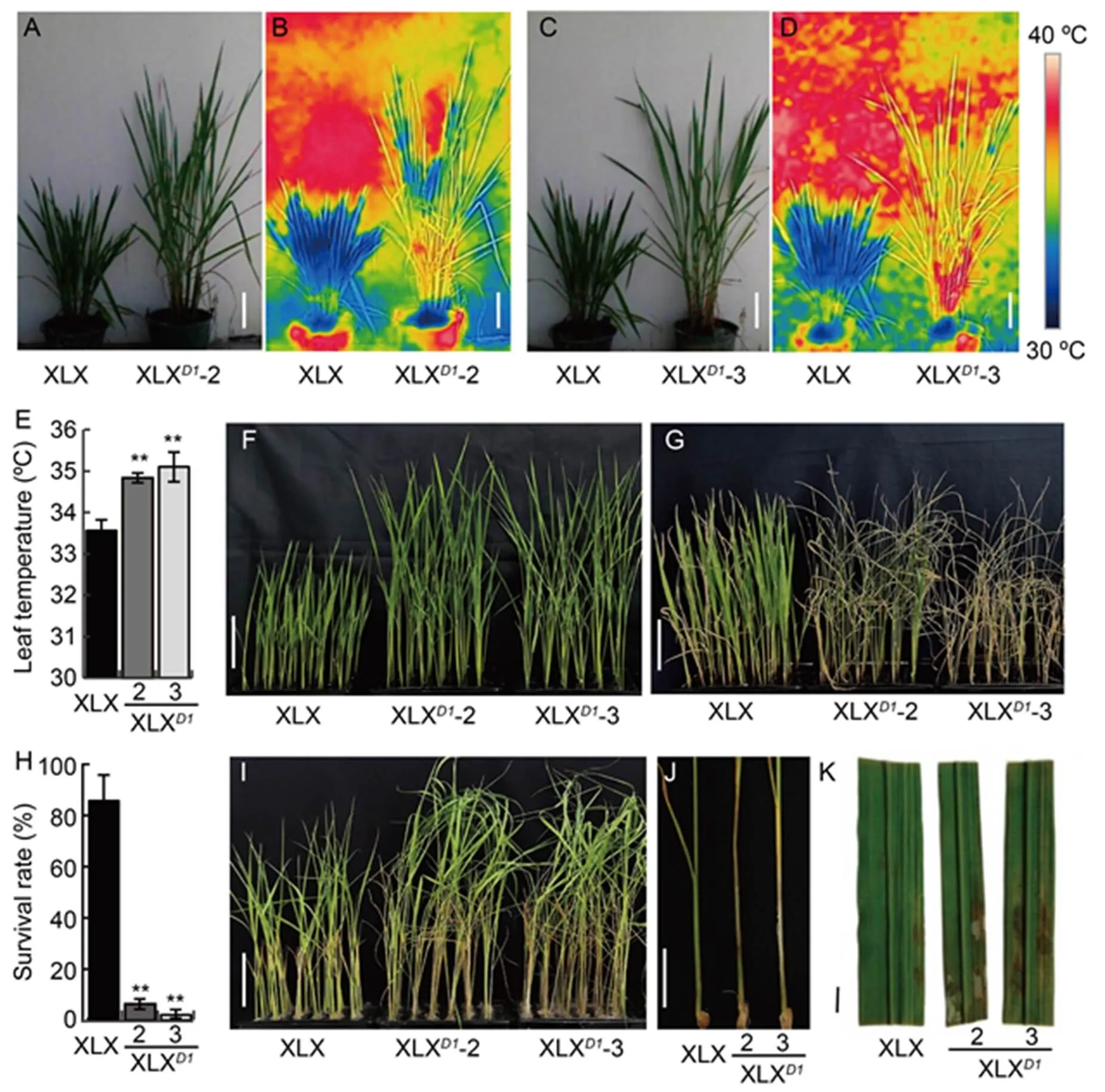
Fig. 8. Xiaolixiang (XLX) exhibits stronger drought and sheath blight resistances.
A and C, Representative images of XLX, XLX-2 and XLX-3. Scale bars, 20 cm.
B and D, Thermal images of XLX, XLX-2 and XLX-3. Scale bars, 20 cm.
E, Leaf temperatures of XLX, XLX-2 and XLX-3 in paddy field.
F‒H, Seedlings of XLX, XLX-2 and XLX-3 before 20% polyethylene glycol (PEG) treatment (F), PEG treatment for 13 d (G) and survival rate after PEG treatment (H). Scale bars, 5 cm.
I, Seedlings infected with. Scale bar, 5 cm.
J, Lesion positions on seedlings after inoculation with. Scale bar, 2 cm.
K, Adult leavesinoculated with. The leaves were collected at the tillering stage. Scale bar, 1 cm.
Mean and SD values were obtained from three biological replicates. **,< 0.01 (the Student’s-test).
RGA1 is a component of heterotrimeric G proteins involved in GA and BR signal pathways (Ashikari et al, 1999; Wang et al, 2006).mutants usually show dwarfism, short and wide leaves, erect and dense panicles, and small round seeds (Ashikari et al, 1999; Fujisawa et al, 1999; Zhu et al, 2020). Compared with other varieties, XLX had a typical RGA1-null phenotype, except without dwarfism. Conversely, XLX was a bit higher than many varieties such as TN1, CHT025 and HZ (Fig. 1 and Table 1). Moreover, the crosses of XLX with other varieties used were truly higher than their parents (Fig. 1 and Table 1), suggesting that there is at least one another dominant and functional plant height related gene in XLX that can restrain dwarfism in XLX and increase the plant height of the crosses. Interestingly, we found a novel allele of the ‘Green Revolution Gene’ () in XLX (data not shown). Two non-synonymous SNPs at residues 100 [glutamic acid (E) and glycine residues (G)] and 340 [glutamine (Q) and arginine (R)] inwere identified as functional nucleotide poly- morphisms responsible for a shorter culm length (Asano et al, 2011). Theallele in XLX was not found in eitheror(Asano et al, 2011), and this allele might have strong effects in improving culm length. Therefore, both XLX with the genotype of/and varieties with/or/have normal plant heights, while F1s with heterozygousandloci were higher than their parents.
To date, several alleles ofhave been identified, including() with a 2-bp deletion,with a 19-bp insertion,with an ochre mutation,with an amber mutation,with a 48-bp deletion (Fujisawa et al, 1999),with a 2-bp deletion (Ueguchi-Tanaka et al, 2000),with a 833-bp deletion (Ashikari et al, 1999), and(Peng et al, 2019) and(Zhu et al, 2020) both with an SNP at the splice site. All these alleles result in null Gα subunits. Although most of thesenull allele mutants have high stress tolerance and NUE, they are not available for breeding because of their low biomass. Therefore, isolating low-expression or weak function ofalleles is necessary to avoid the reduced grain yield. Although anmutant was identified, the transcript ofis repressed by epigenetic silencing to various degrees (Miura et al, 2009). Plants with theallele are unstable and cannot be accurately controlled. However,isolated from XLX was a stably function-weakenedallele, and it contained a unique SNP in the first intron, leading to alternative splicing (Fig. 2-B and -C; Table S3). The pre-mRNA ofin XLX produced both functional and non-functional transcripts, and the normal transcript level was significantly reduced in XLX (Fig. 2-D).
Unlike othermutants, plants withhad more tiller number per plant, more grain number per panicle, and higher seed-setting rate, leading to increased grain yield (Table 2). Grain yield and quality are closely related to photosynthesis at the filling stage (Murchie et al, 2002). The wide, erect and dark green leaves of XLX ensured adequate photosynthesis at the filling stage (Fig. 5-A to -E). Leaf angle is important for light interception, and an upright leaf is a requisite for ideal rice architecture breeding (Wang et al, 2020). Lhcbs form the light-harvesting complex II for light harvesting and are essential for photosynthesis (Caffarri et al, 2004). Bothandwere up- regulated in XLX compared with XLX. The increased photoavoidance and photoprotection inare interested agronomic traits (Ferrero-Serrano et al, 2018), and erect leaf disposition reduces the incidence of irradiance peaks at midday to escape the damage of photoinhibition (Ferrero-Serrano et al, 2018). In this study, we showed that XLX plants were less sensitive to photoinhibition relative to the wild type. Chlorophyll fluorescence confirmed there was an increase in variable fluorescence (v/m) (Fig. 5-G), indicating a higher potential quantum yield and a lower photo- inhibitory damage degree. Furthermore,improved drought and sheath blight resistances (Fig. 8). We speculated that the decreased ROS accumulation in XLX contributed to these phenomena (Fig. 6-A). The over-production of ROS disrupts cell membranes, promotes protein and chloroplast degradation, and finally results in cell apoptosis and early leaf senescence (Yang et al, 2016; Hong et al, 2018). Several research efforts have demonstrated thatmutants usually display less ROS accumulation than the wild type (Peng et al, 2019; Pathak et al, 2021).
Furthermore, the protein and nitrogen contents were higher in XLX grains than in the wild type grains (Fig. 7-F and -G). These results suggested that plants with theallele might have high NUE. Several studies have reported that the G protein complex regulates nitrogen signaling and nitrogen uptake in rice (Sun et al, 2014; Ferrero-Serrano and Assmann, 2016; Zhu et al, 2020).encodes the γ subunit of G proteins in rice, and the differentalleles confer different nitrogen responses.increases nitrogen uptake and assimilation, improving the harvest index and grain yield. The Gβ subunit has a similar effect to DEP1 on NUE, while RGA1 has the opposite effect, and the reduced RGA1 activity inhibits nitrogen responses (Sun et al, 2014)., amutant, shows increased nitrogen uptake (Zhu et al, 2020). To explore the molecular mechanisms offunction in nitrogen-use processes, Pathak et al (2021) performed microarray analyses and identified differentially expressed genes in a classicmutant. They found that loss of function ofperturbs the expression of N-regulated transcription factors (i.e. Nin-like, WRKY and bZIP) and their target genes (i.e.,,and), indicating that RGA1 plays important roles in nitrogen uptake, translocation, signaling and NUE (Pathak et al, 2021).
In conclusion, thegene isolated from XLX was a weak allele of. Plants withallele kept many good characteristics ofnull mutants, such as lodging resistance, and biotic and abiotic stress tolerance.Furthermore,avoided the disadvantageous traits which are always caused by otheralleles, and both grain yield and grain quality (including external quality, milling quality and nutritional quality) were greatly increased in XLX. Therefore,seemed to be a perfectallele and a potential resource for rice breeding.
METHODS
Rice materials and growth conditions
Five hundred local varieties were screened for usable germplasm resources. XLX was selected because of the dep-srs phenotype. An F2mapping population was generated from a cross between XLX and TN1 (ssp.). All plants were grown in a paddy field at Jiangxi Agricultural University in Nanchang (28.77º E, 115.84º N), Jiangxi Province, China, and Sanya (109.51º E, 18.25º N), Hainan Province, China. For drought treatment, three-leaf seedlings were cultured in nutrient solution containing 20% PEG in a growth chamber at 30 ºC/25 ºC with a day/night light cycle for 13 d. The three-leaf seedlings in greenhouse were used for rapid identification of rice sheath blight resistance (Wang et al, 2009). In brief, agar blocks withhyphae were put at the base of each rice seedling, then grown under greenhouse conditions (28 ºC, 90% relative humidity) for 3 d. The lesion heights (LH) and the first leaf sheaths (FLSH) were measured to calculate the relative disease grade scores which were equal to 9 × LH / FLSH. At least 15 individual plants were used for each treatment, and all experiments were repeated three times. Sheath blight resistance was confirmed using adult leavesinoculated with(Sun et al, 2021).
Fine mapping of DEP-SRS
A mapping population was derived from a cross between XLX and TN1. A total of 372 individuals with a typical dep-srs phenotype were used for mapping. Bulked segregant analysis was performed with a mix of DNA from 30 individuals. For linkage analysis and primary mapping, 73 polymorphic simple sequence repeat markers distributed among all the 12 rice chromosomes were selected and detected the polymorphisms between normal plants and dep-srs plants. For fine mapping, new markers between the two flanking markers were designed based on the differences between the published genomic sequences of Nipponbare and 93-11 (www.gramene.org/ resources). Polymorphisms between XLX and TN1 were detected by PCR, and the polymorphic markers were used to screen for recombinants. All primers used for mapping are shown in Table S2. The PCR products were separated on 8% polyacrylamide gel.
Plasmid construction and plant transformation
For the complementation assay, a 6640-bp genomic fragment comprising the 2-kbpromoter region, coding sequence and 3′-UTR was amplified from the genomic DNA of NPB plants using COM-F and COM-R primers (Table S2). The amplified fragment was inserted into pCAMBIA1300 at theRI site using a recombinase (Vazyme, ClonExpress II One Step Cloning Kit, China), and the RGA1-COM construct was generated. The RGA1-COM plasmid was transformed intostrain EHA105. The transformation of XLX plants was performed as previously described (Li and Li, 2003).
Measurements of pigment content and ROS accumulation
The total pigment in the leaves was extracted with 80% acetone, and the contents of chlorophyll and carotenoid were estimated according to Porra et al (1994) using a spectrophotometer (Shimadzu UV2400, Japan). The accumulations of O2·̄ and H2O2were measured via NBT and DAB staining, respectively. Staining and bleaching of the leaves were conducted as previously described (Yang et al, 2016).
Determination of mechanical strength, photosynthetic rate and leaf temperature
The main culm of 10 plants were collected at the early-filling and late-filling stages, and the leaf sheaths on every node were removed. The above four rice internodes were selected to measure the breaking force with a force testing device (YF-1200, Henan YunfeiTechnique & Development Co. Ltd, China). The photosynthetic rate was measured at the heading and filling stages from 10:00 am to 12:00 am on a sunny day with a portable photosynthesis system (LI-6400, LICOR Biosciences, Lincoln, NE, USA). Leaf temperatures were determined with thermal images obtained using a FLIR C3 thermal imaging camera (FLIR Systems, USA). Infrared thermal images were taken between 12:00 am and 13:00 pm with plants set at a 1-m distance from the camera. Three points in the middle of the flag leaf of the primary tiller at the peak tillering stage were used to calculate the leaf temperature. All experiments were performed with more than five independent individuals as biological replicates.
RNA extraction and qRT-PCR
Total RNA was extracted from rice tissues using a TaKaRa Plant MiniBEST RNA Extraction Kit (TaKaRa, Dalian, China), and reverse transcription was performed using a PrimeScriptTMII reverse transcriptase (TaKaRa, Dalian, China) as described by the manufacturer. qRT-PCR was performed using 2× SYBR Green PCR Master Mix (Applied Biosystems, Massachusetts, USA) in a 7500 Real-time PCR System (Applied Biosystems,Massachusetts, USA) with at least three PCR replicates per sample. The mRNA expression levels of these genes were quantified, including normal and nulltranscripts in XLX, NPB and TN1, and,,,,,,,andin XLX and RGANPBtransformed plants.() gene was used as a control. The specific primers used for quantification are listed in Table S2. The 2-ΔCtand 2-ΔΔCtmethods were used for quantification. All of the experiments were performed in triplicate.
ACKNOWLEDGEMENTS
This study was supported by grants from the National Natural Science Foundation of China (Grant Nos. 31960403 and 31501286), and Jiangxi Natural Science Foundation-Outstanding Youth Science Fund Project, China (Grant No. 20212ACB215003).
SUPPLEMENTAL DATA
The following materials are available in the online version of this article at http://www.sciencedirect.com/journal/rice-science; http://www.ricescience.org.
Table S1. Heredity model of dep-srs phenotype in Xiaolixiang.
Table S2. Sequences of primers in this study.
Table S3.sequence diversities in NPB, WYJ7, TN1, NJ06 and XLX.
Table S4. Culm strength analysis of Xiaolixiang (XLX) and XLXplants.
Table S5. Distributions of sheath blight score of Xiaolixiang (XLX) and XLXseedlings.
Alder A, Jamil M, Marzorati M, Bruno M, Vermathen M, Bigler P, Ghisla S, Bouwmeester H, Beyer P, Al-Babili S. 2012. The path from β-carotene to carlactone, a strigolactone-like plant hormone., 335: 1348–1351.
Asano K, Miyao A, Hirochika H, Kitano H, Matsuoka M, Ashikari M. 2010., which encodes a plant-specific novel protein, controls plant elongation by regulating cell division in rice., 86(3): 265–273.
Asano K, Yamasaki M, Takuno S, Miura K, Katagiri S, Ito T, Doi K, Wu J Z, Ebana K, Matsumoto T, Innan H, Kitano H, Ashikari M, Matsuoka M. 2011. Artificial selection for a green revolution gene duringrice domestication., 108(27): 11034–11039.
Ashikari M, Wu J, Yano M, Sasaki T, Yoshimura A. 1999. Rice gibberellin-insensitive dwarf mutant geneencodes the alpha-subunit of GTP-binding protein., 96(18): 10284–10289.
Assmann S M. 2005. G protein regulation of disease resistance during infection of rice with rice blast fungus., 2005: cm13.
Caffarri S, Croce R, Cattivelli L, Bassi R. 2004. A look within LHCII: Differential analysis of the Lhcb1-3 complexes building the major trimeric antenna complex of higher-plant photosynthesis., 43(29): 9467–9476.
Chen W W, Cheng Z J, Liu L L, Wang M, You X M, Wang J, Zhang F, Zhou C L, Zhang Z, Zhang H, You S M, Wang Y P, Luo S, Zhang J H, Wang J L, Wang J, Zhao Z C, Guo X P, Lei C L, Zhang X, Lin Q B, Ren Y L, Zhu S S, Wan J M. 2019., encoding an HD-Zip II family transcription factor, regulates plant development by modulating gibberellin biosynthesis in rice., 288: 110208.
Cui Y, Jiang N, Xu Z J, Xu Q. 2020. Heterotrimeric G protein are involved in the regulation of multiple agronomic traits and stress tolerance in rice., 20(1): 90.
Feng B H, Yang Y, Shi Y F, Lin L, Chen J, Wei Y L, Leung H, Wu J L. 2013. Genetic analysis and gene mapping of light brown spotted leaf mutant in rice., 20(1): 13–18.
Ferrero-Serrano Á, Assmann S M. 2016. The α-subunit of the rice heterotrimeric G protein, RGA1, regulates drought tolerance during the vegetative phase in the dwarf rice mutant., 67(11): 3433–3443.
Ferrero-Serrano Á, Su Z, Assmann S M. 2018. Illuminating the role of the Gα heterotrimeric G protein subunit, RGA1, in regulating photoprotection and photoavoidance in rice., 41(2): 451–468.
Ferrero-Serrano Á, Cantos C, Assmann S M. 2019. The role of dwarfing traits in historical and modern agriculture with a focus on rice., 11(11): a034645.
Fujisawa Y, Kato T, Ohki S, Ishikawa A, Kitano H, Sasaki T, Asahi T, Iwasaki Y. 1999. Suppression of the heterotrimeric G protein causes abnormal morphology, including dwarfism, in rice., 96(13): 7575–7580.
Gilman A G. 1987. G proteins: Transducers of receptor-generated signals., 56:615–649.
Hedden P. 2003. The genes of the green revolution., 19(1): 5–9.
Hong Y B, Zhang Y X, Sinumporn S, Yu N, Zhan X D, Shen X H, Chen D B, Yu P, Wu W X, Liu Q E, Cao Z L, Zhao C D, Cheng S H, Cao L Y. 2018. Premature leaf senescence 3, encoding a methyltransferase, is required for melatonin biosynthesis in rice., 95: 877–891.
Hong Z, Ueguchi-Tanaka M, Shimizu-Sato S, Inukai Y, Fujioka S, Shimada Y, Takatsuto S, Agetsuma M, Yoshida S, Watanabe Y, Uozu S, Kitano H, Ashikari M, Matsuoka M. 2002. Loss-of- function of a rice brassinosteroid biosynthetic enzyme, C-6 oxidase, prevents the organized arrangement and polar elongation of cells in the leaves and stem., 32(4): 495–508.
Hu J, Zhu L, Zeng D L, Gao Z Y, Guo L B, Fang Y X, Zhang G H, Dong G J, Yan M X, Liu J, Qian Q. 2010. Identification and characterization of, a novel gene regulating leaf morphology and plant architecture in rice., 73(3): 283–292.
Khush G S. 2001. Green revolution: The way forward., 2(10): 815–822.
Li M R, Li H Q. 2003. A simple and highly efficient- mediated rice transformation system., 36(4): 289–294. (in Chinese with English abstract)
Li S, Tian Y H, Wu K, Ye Y F, Yu J P, Zhang J Q, Liu Q, Hu M Y, Li H, Tong Y P, Harberd N P, Fu X D. 2018. Modulating plant growth-metabolism coordination for sustainable agriculture.,560: 595–600.
Liu C, Li L G. 2016. Advances in molecular understanding of rice lodging resistance., 30(2): 216–222. (in Chinese with English abstract)
Liu F, Wang P D, Zhang X B, Li X F, Yan X H, Fu D H, Wu G. 2018. The genetic and molecular basis of crop height based on a rice model., 247(1): 1–26.
Liu X, Hu Q L, Yan J J, Sun K, Liang Y, Jia M R, Meng X B, Fang S, Wang Y Q, Jing Y H, Liu G F, Wu D X, Chu C C, Smith S M, Chu J F, Wang Y H, Li J Y, Wang B. 2020. ζ-carotene isomerase suppresses tillering in rice through the coordinated biosynthesis of strigolactone and abscisic acid., 13(12): 1784–1801.
Miura K, Agetsuma M, Kitano H, Yoshimura A, Matsuoka M, Jacobsen S E, Ashikari M. 2009. A metastableepigenetic mutant affecting plant stature in rice., 106(27): 11218–11223.
Monna L, Kitazawa N, Yoshino R, Suzuki J, Masuda H, Maehara Y, Tanji M S, Sato M, Nasu S, Minobe Y. 2002. Positional cloning of rice semidwarfing gene,: Rice ‘green revolution gene’ encodes a mutant enzyme involved in gibberellin synthesis., 9(1): 11–17.
Murai M, Hirose S, Sato S, Takebe M. 1991. Effects of dwarfing genes from dee-geo-woo-gen and other varieties on cool temperature tolerance at booting stage in rice., 41(2): 241–254.
Murchie E H, Yang J C, Hubbart S, Horton P, Peng S B. 2002. Are there associations between grain-filling rate and photosynthesis in the flag leaves of field-grown rice?, 53: 2217–2224.
Oki K, Inaba N, Kitagawa K, Fujioka S, Kitano H, Fujisawa Y, Kato H, Iwasaki Y. 2009. Function of the α subunit of rice heterotrimeric G protein in brassinosteroid signaling., 50: 161–172.
Pathak R R, Mandal V K, Jangam A P, Sharma N, Madan B, Jaiswal D K, Raghuram N. 2021. Heterotrimeric G-protein α subunit (RGA1) regulates tiller development, yield, cell wall, nitrogen response and biotic stress in rice., 11: 2323.
Peng P, Gao Y D, Li Z, Yu Y W, Qin H, Guo Y, Huang R F, Wang J. 2019. Proteomic analysis of a rice mutantpossessing a novelallele of heterotrimeric G protein alpha subunit (RGA1) in salt stress with a focus on ROS scavenging., 20(1): 167.
Porra R J, Schäfer W, Cmiel E, Katheder I, Scheer H. 1994. The derivation of the formyl-group oxygen of chlorophyll b in higher plants from molecular oxygen., 219(1/2): 671–679.
Qi W W, Sun F, Wang Q J, Chen M L, Huang Y Q, Feng Y Q, Luo X J, Yang J S. 2011. Rice ethylene-response AP2/ERF factorrestricts internode elongation by down-regulating a gibberellin biosynthetic gene., 157(1): 216–228.
Sazuka T, Kamiya N, Nishimura T, Ohmae K, Sato Y, Imamura K, Nagato Y, Koshiba T, Nagamura Y, Ashikari M, Kitano H, Matsuoka M. 2009. A rice tryptophan deficient dwarf mutant,, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos., 60(2): 227–241.
Sha H J, Liu H L, Zhao G X, Han Z M, Chang H L, Wang J G, Zheng H L, Zhang J F, Yu Y, Liu Y Q, Zou D T, Nie S J, Fang J. 2022. Elitealleles inrice and their breeding applications in northeast China., 10(1): 224–233.
Suharsono U, Fujisawa Y, Kawasaki T, Iwasaki Y, Satoh H, Shimamoto K. 2002. The heterotrimeric G protein alpha subunit acts upstream of the small GTPase Rac in disease resistance of rice., 99(20): 13307–13312.
Sun H Y, Qian Q, Wu K, Luo J J, Wang S S, Zhang C W, Ma Y F, Liu Q, Huang X Z, Yuan Q B, Han R X, Zhao M, Dong G J, Guo L B, Zhu X D, Gou Z H, Wang W, Wu Y J, Lin H X, Fu X D. 2014. Heterotrimeric G proteins regulate nitrogen-use efficiency in rice., 46(6): 652–656.
Sun Q, Yang S, Guo X F, Wang S T, Jia X T, Li S, Xuan Y H. 2021. RAVL1 activatesto negatively regulate rice resistance to sheath blight disease., 28(2): 146–155.
Ueguchi-Tanaka M, Fujisawa Y, Kobayashi M, Ashikari M, Iwasaki Y, Kitano H, Matsuoka M. 2000. Rice dwarf mutant, which is defective in the alpha subunit of the heterotrimeric G protein, affects gibberellin signal transduction., 97(21): 11638–11643.
Urano D, Colaneri A, Jones A M. 2014. Gα modulates salt-induced cellular senescence and cell division in rice and maize., 65(22): 6553–6561.
Vikram P, Swamy B P M, Dixit S, Singh R, Singh B P, Miro B, Kohli A, Henry A, Singh N K, Kumar A. 2015. Drought susceptibility of modern rice varieties: An effect of linkage of drought tolerance with undesirable traits., 5: 14799.
Wang F M, Yoshida H, Matsuoka M. 2021. Making the ‘green revolution’ truly green: Improving crop nitrogen use efficiency., 62(6): 942–947.
Wang L, Xu Y Y, Ma Q B, Li D, Xu Z H, Chong K. 2006. Heterotrimeric G protein alpha subunit is involved in rice brassinosteroid response., 16(12): 916–922.
Wang R N, Liu C, Li Q Z, Chen Z N, Sun S Y, Wang X L. 2020. Spatiotemporal resolved leaf angle establishment improves rice grain yield via controlling population density., 23(9): 101489.
Wang Z B, Zuo S M, Li G, Chen X J, Chen Z X, Zhang Y F, Pan X B. 2009. Rapid identification technology of resistance to rice sheath blight in seedling stage., 39: 174–182. (in Chinese with English abstract)
Yaish M W, El-Kereamy A, Zhu T, Beatty P H, Good A G, Bi Y M, Rothstein S J. 2010. The APETALA-2-like transcription factor OsAP2-39 controls key interactions between abscisic acid and gibberellin in rice., 6(9): e1001098.
Yang Y L, Xu J, Huang L C, Leng Y J, Dai L P, Rao Y C, Chen L, Wang Y Q, Tu Z J, Hu J, Ren D Y, Zhang G H, Zhu L, Guo L B, Qian Q, Zeng D L. 2016., encoding chlorophyllide a oxygenase 1, impacts leaf senescence and indirectly affects grain yield and quality in rice., 67(5): 1297–1310.
Yuan L P. 2014. Development of hybrid rice to ensure food security., 21(1): 1–2.
Zhu Y C, Li T, Xu J, Wang J J, Wang L, Zou W W, Zeng D L, Zhu L, Chen G, Hu J, Gao Z Y, Dong G J, Ren D Y, Shen L, Zhang Q, Guo L B, Hu S P, Qian Q, Zhang G H. 2020. Leaf width geneaffects plant architecture and yield in rice by regulating nitrogen utilization efficiency., 157: 359–369.
27 January 2022;
28 March2022
Copyright © 2022, China National Rice Research Institute. Hosting by Elsevier B V
This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/4.0/)
Peer review under responsibility of China National Rice Research Institute
http://dx.doi.org/10.1016/j.rsci.2022.03.001
Xu Jie (xujie198615@foxmail.com); He Haohua (hhhua64@163.com)
(Managing Editor: Wu Yawen)
杂志排行

Rice Science的其它文章
- Transcriptome Analysis Provides Novel Insights into Salt Stress Response in Two Egyptian Rice Varieties with Different Tolerance Levels
- QTL Mapping for Plant Height Using Introgression Lines Derived from Zhonghui 8015 and Wild Rice (Oryza rufipogon)
- Genetic Dissection of Quantitative Trait Loci for Panicle Traits and Heat Tolerance by High-Density Bin Map in Rice
- Improvement of Rice Production under Drought Conditions in West Africa:Application of QTLs in Breeding for Drought Resistance
- CRISPR/Cas9-Targeted Knockout of Rice Susceptibility Genes OsDjA2 and OsERF104 Reveals Alternative Sources of Resistance to Pyricularia oryzae
- Identification of Potential Zinc Deficiency Responsive Genes and Regulatory Pathways in Rice by Weighted Gene Co-expression Network Analysis