沉默BLM DNA解旋酶基因对三阴性乳腺癌MDA-MB-468细胞增殖和凋亡的影响*
2022-10-19黄亚兰张望明刘小花余晓静安丽君吴介恒杨留启刘杰麟
黄亚兰,张望明,刘小花,余晓静,安丽君,吴介恒,杨留启,刘杰麟,2**
(1.贵州医科大学 基础医学院免疫学教研室,贵州 贵阳 550025;2.贵州医科大学 省部共建药用植物功效与利用国家重点实验室,贵州 贵阳 550014)
乳腺癌是全世界女性最常见的恶性肿瘤之一,也是女性癌症患者死亡的主要原因[1-2]。根据乳腺癌激素表达情况以及基因表达可将乳腺癌分为Luminal A型、Luminal B型、HER-2过表达型及三阴性型多种亚型[3],其中三阴性乳腺癌(triple negative breast cancer,TNBC)易发生侵袭转移、导致局部和远处复发率高,是乳腺癌亚型中预后较差的一种。TNBC的定义是不表达雌激素受体(ER)、孕激素受体(PR)和人类表皮生长因子受体-2(HER-2),因此对内分泌药物缺乏敏感性,以及靶向治疗的选择有限,使这类肿瘤的替代治疗策略备受关注[4]。RecQ DNA解旋酶是一个在细菌、真菌、植物和动物中发现的蛋白质家族,是一类通过水解ATP高能磷酸键、获得能量后解开DNA双链结构并产生单链DNA的酶[5]。人体内含有5种RecQ DNA解旋酶,分别是RECQL1、BLM、WRN、RECQL4和RECQL5,这些解旋酶在细胞中发挥重要作用,包括DNA复制、转录、DNA修复和端粒维持等[6-7]。有研究表明BLM DNA解旋酶与癌症的发展有着密切联系[8-9],BLM DNA解旋酶mRNA表达增加与乳腺癌患者无远处转移生存期的减少相关[10]。课题组前期研究发现BLM DNA解旋酶蛋白在结肠癌及癌旁组织中均有表达,但在癌组织中表达更高、且与TNM临床分期相关[11]。前列腺癌组织和细胞中BLM DNA解旋酶的表达明显高于非前列腺癌组织和细胞,干扰BLM DNA解旋酶的表达可抑制前列腺癌细胞增殖,并促进细胞凋亡,从而减缓疾病的进程[12]。目前关于BLM DNA解旋酶在TNBC中的作用研究较少,本研究在体外以三阴性乳腺癌MDA-MB-468细胞为研究对象,探讨沉默BLMDNA解旋酶基因对乳腺癌细胞增殖和凋亡的影响及机制,为TNBC分子靶向治疗提供依据。
1 材料与方法
1.1 材料
1.1.1组织标本 收集2012—2014年三阴性乳腺癌30例、乳腺纤维腺瘤20例和癌旁正常对照石蜡包埋组织12例,临床资料完整,所有患者在手术切除肿瘤前均未进行放化疗。全部的人组织标本使用按照贵州医科大学附属医院生物伦理学会的要求,对病人的隐私予以保密。
1.1.2细胞与主要试剂 人正常乳腺上皮MCF-10A细胞购自中国科学院细胞库,乳腺癌MCF-7细胞及三阴性乳腺癌MDA-MB-468细胞由贵州医科大学免疫学实验室保存,DMEM培养基、青霉素和链霉素(美国Gibco公司),胎牛血清(杭州四季青公司),Lipofectamine 2000(美国 Invitrogen公司),RIPA 强裂解液、BCA 蛋白浓度测定试剂盒(江苏碧云天公司),MTT 试剂(北京索莱宝公司),细胞凋亡试剂盒(杭州联科生物公司),BLM抗体(美国Santa Cruz Biotechnology公司),γ-H2AX、Bax、Bcl-2、cleaved Caspase-3、cleaved Caspase-8抗体(成都正能生物公司),Rad51抗体(北京博奥森生物公司),BRCA1抗体(武汉三鹰生物公司),GAPDH抗体(武汉赛维尔公司)。
1.1.3主要仪器 凝胶成像系统(上海天能公司),恒温干燥箱(上海医疗器械厂),超微量微孔板分光光度计(美国biotek公司),正置荧光显微镜、倒置荧光显微镜(日本尼康公司),流式细胞分析仪(美国BD公司)。
1.2 研究方法
1.2.1免疫组织化学(IHC)染色检测BLM DNA解旋酶的表达 利用IHC二步法检测三阴性乳腺癌、乳腺纤维腺瘤和癌旁组织中BLM DNA解旋酶的表达情况,BLM DNA解旋酶单克隆抗体工作浓度为1∶500。由有经验的病理学专业老师对切片IHC染色进行评估,细胞核或(和)细胞浆出现棕黄色即认为是BLM DNA解旋酶蛋白质的阳性表达。免疫组织化学评分(IHS)根据染色的强度和阳性表达细胞的百分比。染色强度的评分标准:阴性(未着色)为0分,弱阳性(淡黄色)为1分,中度阳性(棕黄色)为2分,强阳性(棕色)为3分。阳性细胞百分比的评分标准:阳性细胞比例≤5%(阴性)为0分,阳性细胞比例6%~25%(弱阳性)为1分,阳性细胞比例26%~50%(中度阳性)为2分,阳性细胞比例51%~75%(阳性)为3分,阳性细胞比例≥76%(强阳性)为4分。每张切片的蛋白质IHC染色指数为染色强度评分与阳性细胞比例评分的乘积。
1.2.2细胞培养 人正常乳腺上皮MCF-10A细胞用专用培养基培养,乳腺癌MCF-7细胞及三阴性乳腺癌MDA-MB-468细胞,在含10%胎牛血清的DMEM培养液中培养,均置37 ℃、5% CO2培养箱中培养。
1.2.3siRNA设计合成及转染 由上海吉玛公司合成3对靶向BLMDNA解旋酶基因的siRNA序列和1对阴性对照siRNA。siRNA-BLM 1正义链为5′-CGGCCAAUUAAAUCAGUAUTT-3′,反义链为5′-AUACUGAUUUAAUUGGCCGTT-3′;siRNA-BLM 2 正义链为 5′-GGAGGUGGUAAGAGUUUGUTT-3′,反义链为5′-ACAAACUCUUACCACCUCCTT-3′;siRNA-BLM 3正义链为5′-GCCACAUGUAGAAAGAUAUTT-3′,反义链为5′-AUAUCUUUCUACAUGUGGCTT-3′;Negative Control正义链为5′-UUCUCCGAACGUGUCACGUTT-3′,反义链为5′-ACGUGACACGUUCGGAGAATT-3′。染前24 h将MDA-MB-468细胞以每孔5×105个铺至6孔板,转染时设置5个组分别为转染组(Control)、siRNA-NC、siRNA-BLM 1、siRNA-BLM 2和siRNA-BLM 3,以Lipofectamine 2000为转染试剂,转染过程参照转染试剂说明书。转染6 h后,用含10%胎牛血清的DMEM培养液更换6孔板中的转染液体,于37 ℃、5%CO2培养箱中继续培养48 h后提取细胞蛋白检测沉默效果,软件测量各组条带灰度值(A)。沉默效率(%)=[1-(A干扰组BLM×A未转染组GAPDH)/(A干扰组GAPDH×A未转染组BLM)]×100%,选取沉默效率最高的siRNA序列构建shRNA干扰载体慢病毒,由上海吉凯基因公司完成。
1.2.4慢病毒感染及稳定细胞株筛选 将对数生长期的MDA-MB-468细胞以5×105个/孔接种到6孔板中。将实验分为shRNA-NC组和shRNA-BLM组,分别为感染阴性对照慢病毒组和感染shRNA-BLM重组慢病毒组。感染步骤:去除6孔板内液体,加DMEM 1 mL、慢病毒感染增强液HitransG P 40 μL 及病毒液15 μL,培养12 h更换细胞培养液,继续培养3 d,用含嘌呤霉素2 mg/L的培养基筛选稳定沉默BLMDNA解旋酶基因的细胞株用于后续实验。shRNA-NC序列为TTCTCCGAACGTGTCACGT,shRNA-BLM序列为GCCACATGTAGAAAGATAT。
1.2.5MTT法检测细胞活力 将shRNA-BLM组和shRNA-NC组细胞接种至96孔板,接种密度为1.2×104个/孔,于37 ℃、5% CO2培养箱及饱和湿度条件下培养24 h、48 h、72 h后,每孔加入MTT 20 μL继续培养4 h,弃去上清液,每孔加DMSO 150 μL,震荡后用微量微孔板分光光度计在490 nm处检测各孔光吸收值(OD)。
1.2.6平板克隆形成实验检测细胞集落形成能力 shRNA-BLM组和shRNA-NC组细胞以每孔3×103个/孔接种在含有1.5 mL培养液的12孔板中,置于37 ℃,5%CO2培养箱中培养。8 d后,弃去培养液,PBS洗涤两次,4%多聚甲醛室温固定20 min,0.1%结晶紫染色20 min,蒸馏水将染液冲洗干净,计数细胞集落。
1.2.7流式细胞术检测细胞凋亡 收集shRNA-BLM组和shRNA-NC组细胞,用预冷PBS洗涤两次,离心后弃掉上清液。加入1×Binding Buffer工作液重悬细胞,在每管细胞样本中各加入7-AAD 10 μL和Annexin V-APC 5 μL,用移液器轻轻混匀,室温条件下避光染色5 min上机检测。
1.2.8单细胞凝胶电泳技术检测细胞DNA损伤 在shRNA-BLM组和shRNA-NC组细胞中加入PBS,调整细胞密度为1×106个/mL。配制0.5%普通正常熔点的琼脂糖,覆盖至载玻片上,干燥后为第一层胶。取细胞悬液 10 μL与0.7%低熔点琼脂糖 75 μL混匀,滴加至第一层胶上。将载玻片浸入细胞裂解液中,4 ℃裂解2 h,PBS洗涤。载玻片放置在水平电泳槽中,加入碱性电泳缓冲液,室温条件避光解旋1 h。设置电压为25 V、电泳30 min,取出载玻片,0.4 mmol/L Tris-HCL缓冲液中和3次、每次10 min。滴加PI染液20 μL至凝胶上,于正置荧光显微镜下观察,分析细胞尾部DNA含量。
1.2.9γ-H2AX免疫细胞化学染色 PBS洗涤细胞样本,4%多聚甲醛室温固定20 min,PBS洗涤3次、每次3 min;滴加1%Triton X-100通透液4 min,PBS漂洗3次;PBS配制3%BSA,室温封闭30 min,PBS洗涤;PBS以400∶1稀释γ-H2AX抗体,加至样本上,4 ℃过夜。室温平衡30 min后,加入山羊抗鼠二抗,37 ℃孵育1 h,PBS漂洗3次;DAB显色1 min,PBS漂洗,滴加苏木素染液,室温下染色3 min,蒸馏水冲洗。通风条件下晾干后滴加中性树胶封片,显微镜下观察计数γ-H2AX阳性细胞。
1.2.10Western blot法检测细胞DNA损伤修复相关蛋白及凋亡相关蛋白表达 提取各组细胞蛋白,BCA法检测蛋白浓度,加入蛋白上样缓冲液煮沸,使蛋白充分变性。加入蛋白样品后进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳。采用湿转法将蛋白转移至PVDF膜,5%脱脂牛奶封闭2 h,TBST洗涤三次,分别加入一抗BLM、Bax、Bcl-2、cleaved Caspase-3、cleaved Caspase-8、Rad51、BRCA1、GAPDH(TBST 稀释,均为1∶1 000),4 ℃孵育过夜,TBST洗膜,加入对应HRP标记的二抗(TBST稀释,1∶4 000),室温孵2 h,TBST洗膜后曝光显影。以GAPDH作为内参,Image J软件用于分析蛋白条带灰度值,计算目的蛋白相对表达量。
1.3 统计学分析
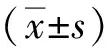
2 结果
2.1 BLM DNA解旋酶在组织与细胞中的表达
在TCGA数据库中发现BLM DNA解旋酶在胆管癌、食管癌、胶质母细胞瘤等常见恶性肿瘤以及其对应的正常组织之间存在差异表达。与正常组织比较,BLM DNA解旋酶在大多数肿瘤中呈现高表达(图1A),在乳腺癌呈明显高表达(图1B)。在GEO数据库中发现BLM DNA解旋酶mRNA高表达与乳腺癌患者总生存期(OS)、无复发生存期(RFS)降低相关(图1C,D)。IHC结果显示,BLM DNA解旋酶的阳性着色定位于细胞核和细胞质(图1E),BLM DNA解旋酶在TNBC组织中表达高于乳腺纤维腺瘤和癌旁组织,差异有统计学意义(P<0.001,表1);在乳腺纤维腺瘤和癌旁组织之间比较,差异无统计学意义(P>0.05,表1)。Western blot结果显示,TNBC MDA-MB-468细胞中BLM DNA解旋酶蛋白表达水平显著高于正常乳腺上皮MCF-10A细胞和乳腺癌MCF-7细胞,差异有统计学意义(图1F,P<0.01)。

表1 三阴性乳腺癌、乳腺纤维腺瘤和癌旁组织中BLM DNA解旋酶的表达
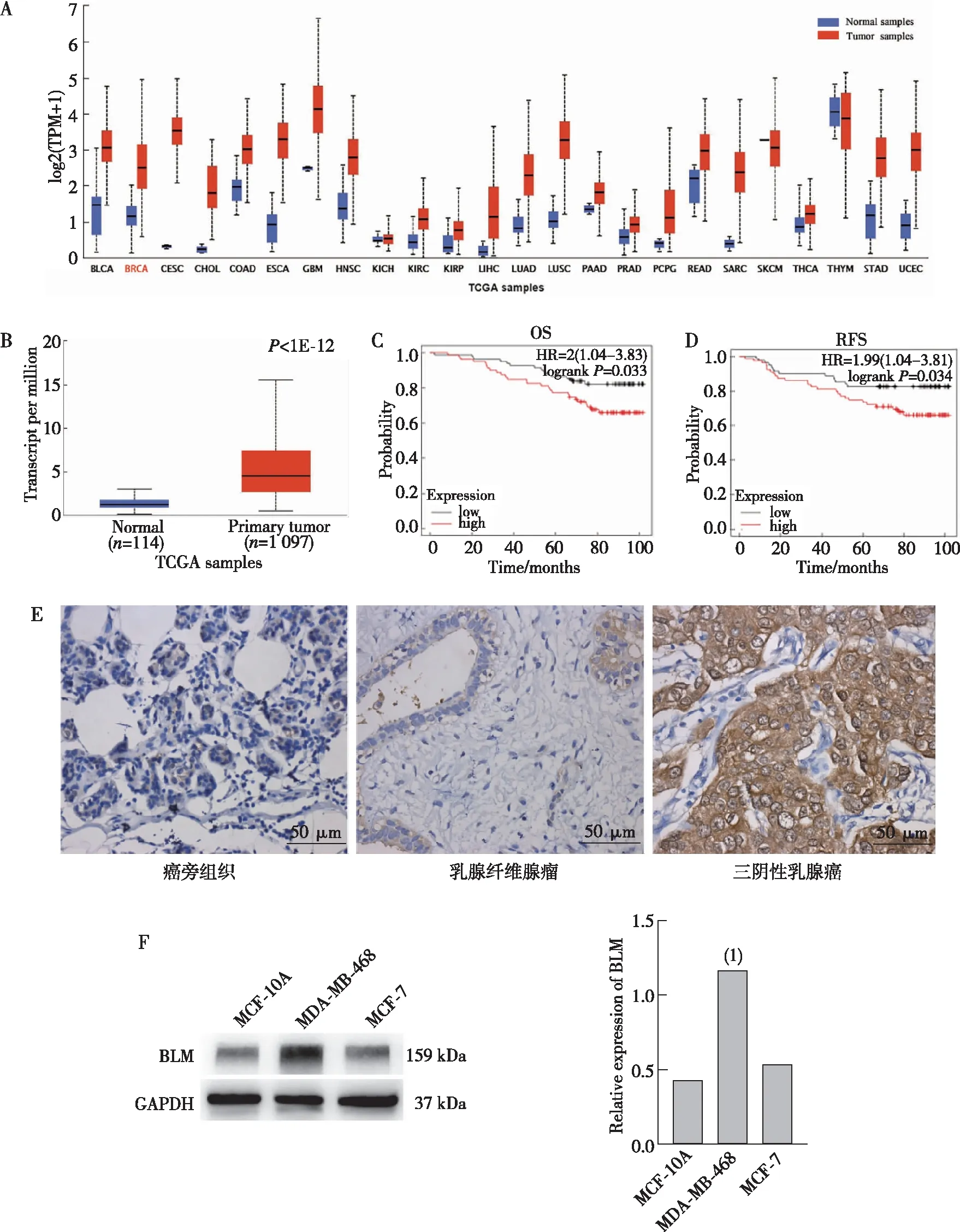
注:A、B为在TCGA数据库中分析BLM DNA解旋酶在组织中的表达情况,C、D为在GEO数据库中分析BLM DNA解旋酶在乳腺癌患者中的预后价值,E为IHC检测BLM DNA解旋酶在组织中的表达情况(DAB显色),F为Western blot检测BLM DNA解旋酶在细胞中的表达情况及定量结果;(1)与MCF-10A和MCF-7组比较,P<0.01。
2.2 沉默三阴性乳腺癌MDA-MB-468细胞中BLM DNA解旋酶基因的表达
为了沉默三阴性乳腺癌细胞中BLMDNA解旋酶基因的表达,将siRNA序列转染MDA-MB-468细胞(图2A)。蛋白印迹结果显示,与Control组比较,siRNA-BLM 3实验组BLM DNA解旋酶表达水平下降,蛋白沉默效率为60.49%(图2B,P<0.05)。选取沉默效率最高的siRNA-3序列构建shRNA干扰载体慢病毒,将shRNA干扰载体慢病毒感染MDA-MB-468细胞(图3A)。Western blot法检测BLM DNA解旋酶蛋白沉默效果,如图3B所示,与shRNA-NC组相比较,shRNA-BLM组BLM DNA解旋酶蛋白表达显著下调,差异有统计学意义(P<0.001)。

注:A为细胞转染FAM荧光基团标记的siRNA序列后于光学显微镜、荧光显微镜下视野(100×);B为BLM DNA解旋酶蛋白表达情况及定量结果;(1)与Control组比较,P<0.05;(2)与siRNA-NC组比较,P<0.01。
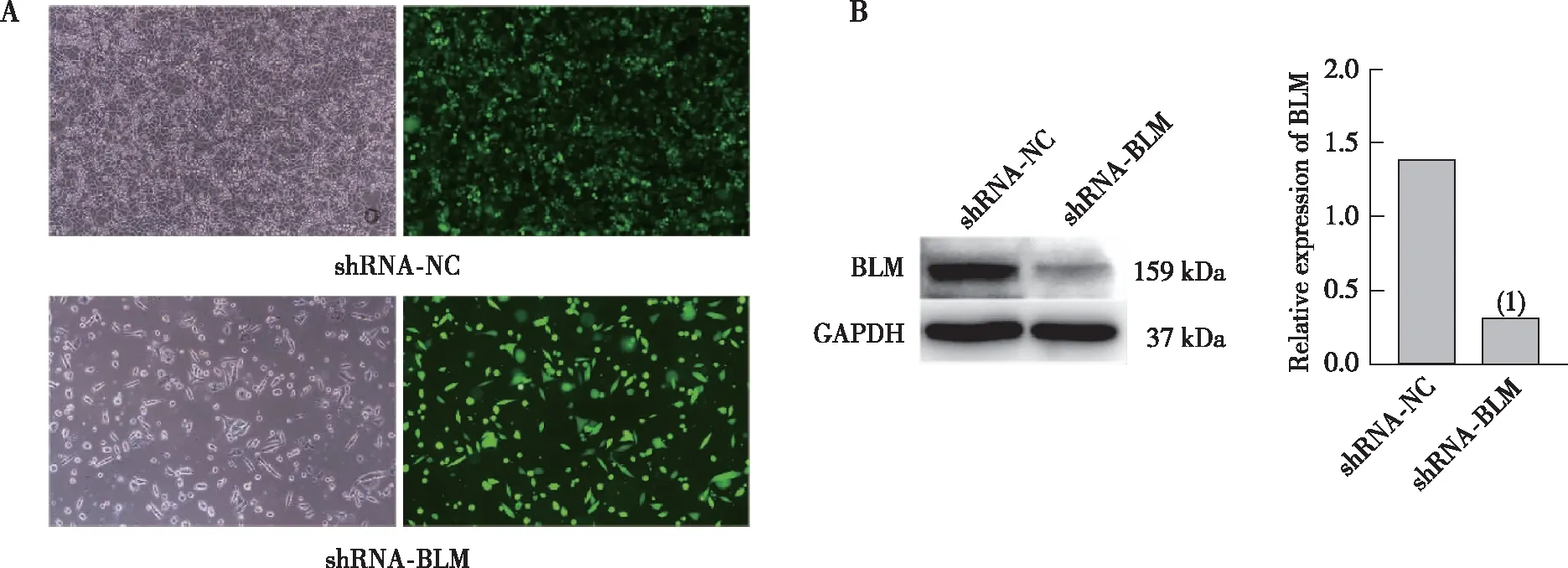
注:A为细胞感染GFP标记的shRNA质粒慢病毒后于光学显微镜、荧光显微镜下视野(100×);B为BLM DNA解旋酶蛋白表达情况及定量结果;(1)与shRNA-NC组比较,P<0.001。
2.3 沉默BLM DNA解旋酶基因抑制MDA-MB-468细胞增殖、集落形成能力
MTT检测结果显示,与shRNA-NC组比较,shRNA-BLM组细胞生长曲线上升平缓。shRNA-BLM组48 h、72 h的OD值均低于shRNA-NC组(图4A,P<0.001)。平板克隆形成实验结果显示,shRNA-BLM组细胞集落数为(194±31),shRNA-NC组细胞集落数为(10±2),shRNA-BLM组细胞集落数低于shRNA-NC组(图4B,P<0.001)。提示下调BLM DNA解旋酶表达显著抑制MDA-MB-468细胞生长及集落形成能力,使其增殖减慢。
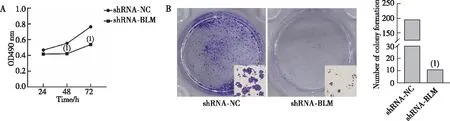
注:A为MTT实验结果,B为平板克隆形成实验结果及定量结果;(1)与shRNA-NC组比较,P<0.001。
2.4 沉默BLM DNA解旋酶基因促进MDA-MB-468细胞凋亡
通过流式细胞术检测shRNA-NC组和shRNA-BLM 组细胞凋亡情况,结果显示,shRNA-BLM沉默组的细胞凋亡率较阴性对照组shRNA-NC显著增加(图5A,P<0.01)。Western blot检测shRNA-NC组和shRNA-BLM细胞凋亡相关蛋白表达情况,与shRNA-NC组相比,shRNA-BLM组Bax、cleaved Caspase-3、cleaved Caspase-8蛋白表达上调,Bcl-2蛋白表达下调(图5B,P<0.05)。说明沉默BLMDNA解旋酶基因促进三阴性乳腺癌MDA-MB-468细胞发生凋亡。

注:A为两组细胞凋亡水平及定量结果,B为凋亡相关蛋白表达水平及定量结果;(1)与shRNA-NC 组比较,P<0.01;(2)与shRNA-NC组比较,P<0.05。
2.5 沉默BLM DNA解旋酶基因增加MDA-MB-468细胞DNA 损伤
为了分析沉默BLMDNA解旋酶基因对DNA损伤的影响,收集shRNA-NC阴性对照组和shRNA-BLM沉默组的细胞进行单细胞凝胶电泳、计算细胞尾部DNA含量;免疫细胞化学法检测DNA损伤标志物γ-H2AX灶的形成,Western blot法检测DNA损伤修复相关蛋白BRCA1、Rad51表达。结果显示,与shRNA-NC组相比较,shRNA-BLM组细胞拖尾更明显,尾部DNA含量更高,差异有统计学意义(图6A,P<0.001);γ-H2AX阳性细胞比例显著增加(图6B,P<0.001);BRCA1、Rad51蛋白表达下调,差异有统计学意义(图6C,P<0.05)。提示沉默BLMDNA解旋酶基因增加三阴性乳腺癌细胞发生DNA损伤,细胞DNA损伤修复能力下降。
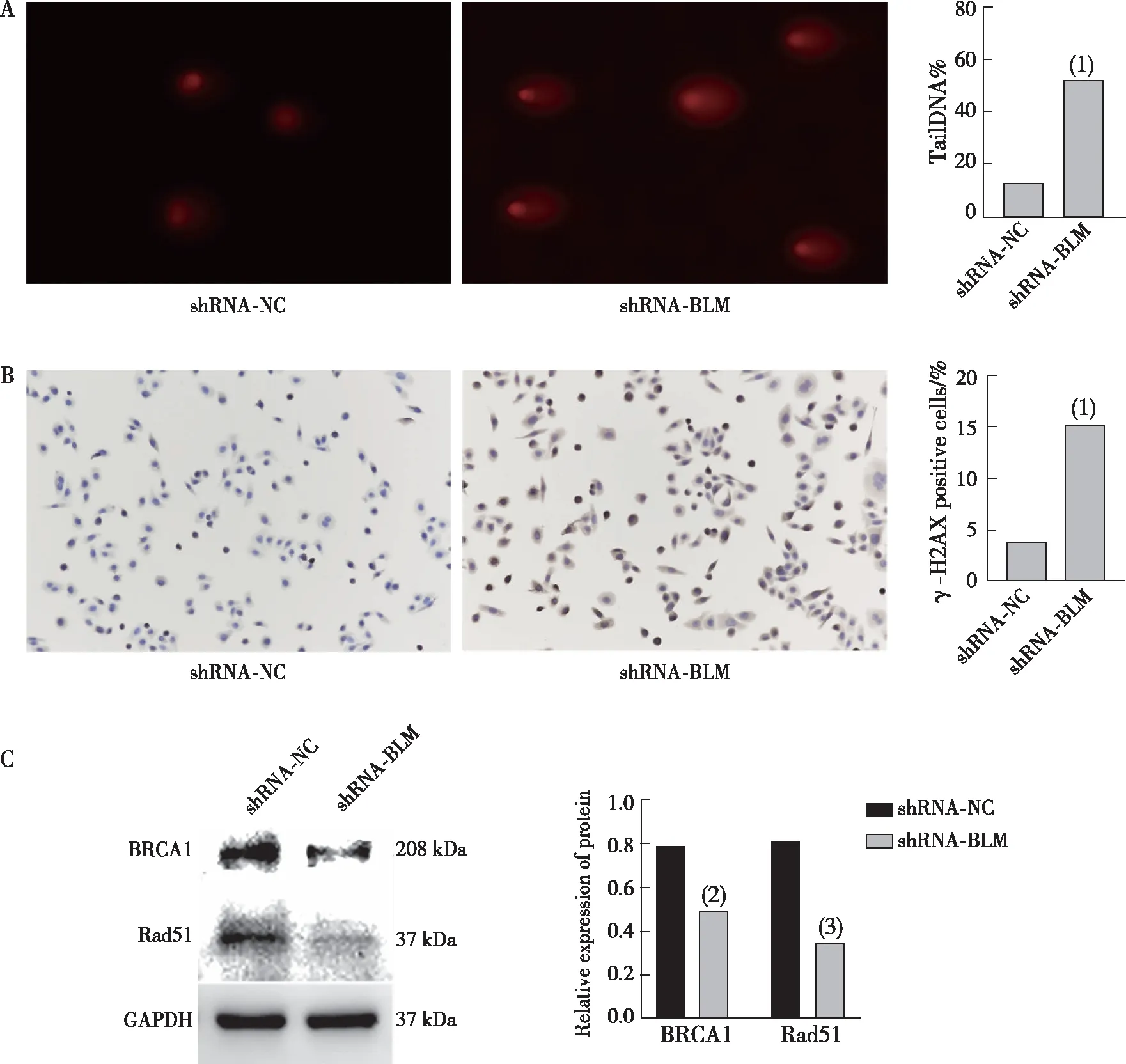
注:A为单细胞凝胶电泳结果及定量结果(DAPI染色),B为免疫细胞化学结果及定量结果(DAB显色),C为DNA损伤修复相关蛋白表达及定量结果;(1)与shRNA-NC 组比较,P<0.001;(2)与siRNA-NC 组比较,P<0.05;(3)与shRNA-NC组比较,P<0.01。
3 讨论
三阴性乳腺癌具有缺乏靶向治疗,患者生存率低、预后差等特点,非靶向治疗策略通常会对机体肾脏、心脏、肝脏和其他正常组织等器官造成毒性作用和不良反应[13]。因此,为三阴性乳腺癌寻找有效的抗癌分子靶点是基础与临床研究的热点,具有重要的临床意义[14]。BLM DNA解旋酶是RecQ DNA解旋酶家族的重要成员之一,由位于15q26.1区带上的BLM DNA解旋酶基因编码[15]。研究显示在恶性肿瘤组织及细胞中发现BLM DNA解旋酶的表达高于癌旁组织和正常细胞,在通过沉默BLM DNA解旋酶表达后,肿瘤细胞周期受到阻滞,细胞的增殖和迁移能力均受到抑制[16]。关于BLM DNA解旋酶在三阴性乳腺癌中作用的研究相对较少,本研究结果显示BLM DNA解旋酶在乳腺癌组织中表达上调,三阴性乳腺癌MDA-MB-468细胞中BLM DNA解旋酶蛋白表达水平高于正常乳腺上皮细胞,提示BLM DNA解旋酶可能参与乳腺癌的发生发展。本实验通过RNAi技术,构建shRNA-BLM干扰载体慢病毒感染MDA-MB-468细胞,探究沉默BLMDNA解旋酶基因表达对乳腺癌细胞增殖和凋亡的影响。结果显示shRNA-BLM组BLM DNA解旋酶蛋白表达水平低于shRNA-NC组,说明沉默BLMDNA解旋酶基因的细胞株构建成功,可用于后续研究。
恶性肿瘤最显著的特征之一是细胞增殖失控,抑制癌细胞增殖在抗癌和疾病防御中起着关键作用[17]。有研究报道下调BLM DNA解旋酶的表达不仅能够抑制人晶体上皮细胞的增殖,还促进细胞发生凋亡[18]。在前列腺癌细胞中BLM DNA解旋酶的过度表达有助于前列腺癌的进展,在沉默BLMDNA解旋酶基因表达后,抑制前列腺癌PC3细胞增殖,并诱导其凋亡[19]。为了检测沉默BLMDNA解旋酶基因的表达对三阴性乳腺癌MDA-MB-468细胞增殖的影响,本研究采用MTT法以及细胞克隆形成实验进行检测,结果显示下调BLM DNA解旋酶表达能够抑制MDA-MB-468细胞增殖以及集落形成能力,表明BLM DNA解旋酶参与了三阴性乳腺癌细胞的增殖,BLM DNA解旋酶有作为治疗三阴性乳腺癌分子靶点的潜力。
细胞凋亡是细胞程序性死亡的一种主要形式,促进细胞凋亡被认为是抗肿瘤的重要机制[20],常见的两条凋亡途径分为内源性凋亡途径和外源性凋亡途径,均有Caspase的参与[21]。Caspase蛋白家族中,Caspase-3属于凋亡执行因子,其活化是细胞进入凋亡的重要标志[22]。Caspase-8属于凋亡启动因子,对多数执行Caspase分子均能激活,进而诱发凋亡。肿瘤细胞通过活化Caspase-8,活化后的Caspase-8特异性切割Caspase-3,生成效应因子cleaved Caspase-3使肿瘤细胞解体导致细胞发生凋亡[23]。Bax是细胞内重要的促凋亡基因,Bcl-2为抑制细胞凋亡基因,两者属于Bcl-2家族。Bax与Bcl-2之间表达水平的改变在细胞凋亡过程中发挥重要作用[24]。研究表明,癌细胞内cleaved Caspase-3、cleaved Caspase-8、Bax表达水平升高,Bcl-2表达水平降低,表明细胞发生凋亡[25]。为了检测沉默BLM DNA解旋酶的表达对三阴性乳腺癌MDA-MB-468细胞凋亡的影响,本研究采用流式细胞术和Western blot法分别检测细胞凋亡率和凋亡相关蛋白表达情况。研究结果显示,沉默BLM DNA解旋酶表达后细胞凋亡率增加,Bax、cleaved Caspase-3、cleaved Caspase-8蛋白表达上调,Bcl-2蛋白表达下调,说明BLM DNA解旋酶沉默后可促进三阴性乳腺癌细胞凋亡,这与张等[26]研究发现沉默在膀胱癌细胞中高表达的BLM DNA解旋酶后,可以促进肿瘤细胞凋亡一致。
基因组的完整性和稳定性对细胞至关重要,当细胞DNA持续暴露于应激源下会导致DNA损伤。当DNA损伤严重时,细胞激活DNA损伤检查点和周期检查点以阻止细胞周期的进展,并促进DNA损伤修复,当DNA修复失败导致基因组不稳定,就会触发程序性细胞死亡[27]。在感知和修复DNA损伤中γ-H2AX起着核心作用,可作为DNA损伤的早期敏感生物标记物[28]。作为DNA修复基因,Rad51基因编码的蛋白质是参与同源重组的关键酶,在维持DNA的遗传稳定性和多样性方面发挥着重要作用[29]。同样作为DNA修复基因的BRCA1目前已知是一种重要的抑癌基因,BRCA1基因编码的蛋白参与细胞代谢的多个过程,包括DNA复制、转录调节等,BRCA1蛋白在DNA修复中的特殊作用是维持基因组的稳定性。BLM DNA解旋酶在DNA修复中起着重要作用,为了检测沉默BLM DNA解旋酶的表达对三阴性乳腺癌MDA-MB-468细胞DNA损伤及修复的影响,本研究通过单细胞凝胶电泳技术检测细胞尾部DNA含量,免疫细胞化学法检测DNA损伤标志物γ-H2AX、Western Blot检测BRCA1、Rad51的表达。结果显示,抑制 BLM DNA解旋酶表达后,细胞尾部DNA含量、γ-H2AX阳性细胞数增加,DNA损伤修复蛋白BRCA1、Rad51 蛋白表达下调,提示沉默BLMDNA解旋酶基因达,三阴性乳腺癌MDA-MB-468细胞DNA损伤增加,DNA损伤修复能力下降。
综上所述,BLM DNA解旋酶在三阴性乳腺癌MDA-MB-468细胞中高表达,shRNA介导的BLMDNA解旋酶基因沉默抑制MDA-MB-468细胞增殖,促进细胞凋亡,DNA损伤修复相关蛋白受抑制。当细胞发生DNA损伤,DNA修复失败导致基因组不稳定,从而触发细胞发生凋亡。因此,靶向抑制BLM DNA解旋酶的表达有望为TNBC治疗提供新的思路。
