连通性的猕猴大脑PE细分分析及同源性研究
2022-09-23王千山李斌强柴静文李海芳
王 玥,王千山,姚 蓉,李斌强,柴静文,李海芳
(太原理工大学 信息与计算机学院,山西 晋中 030600)
猕猴是人类的天然过渡模型,基于影像特征研究人脑与猴脑的跨物种比较是当今信息科学、神经科学等学科的共同前沿。构建更加精细的脑区亚区可以为揭示更全面的脑区认知、神经环路等提供更多的可能性。
PE区(Brodmann’s Area 5)是猕猴顶上小叶(superior parietal lobule,SPL)的一部分,参与手臂运动的控制。已有研究表明,人类Brodmann 5区与PE区是同源区域,了解PE区可为人类手臂运动临床诊断和康复训练提供依据。现有研究表明从细胞构筑的角度来看PE能够细分。基于白质纤维束的解剖连接对于PE的分割还需更多验证,同时细分后的PE与人类脑区的同源关系尚不明确。本文所关注的猕猴PE区是猕猴顶上小叶(superior parietal lobule,SPL)的一部分。SPL参与整合视觉和体感皮层区域的信息,以执行伸展和抓取动作[1]。目前,SPL研究最多的区域是PE、PEc和V6A.其中,PE区位于顶叶后皮质(posterior parietal cortex,PPC)[2],本质上是一个以躯体感觉输入为主的高级躯体运动区,在触达过程中参与深度和方向信息的运动加工。1982年PANDYA et al[3]确认PE存在,并将其定义为细胞构筑同质实体。后期有研究在连接特性上指出了PE内可能的细分[4-6],功能研究也显示了PE可细分[7]。
最近一项神经示踪剂研究表明PE区域并不同质[8]。研究发现根据细胞构筑以及一些被检查受体的平均密度和层状分布模式的不同,PE能够细分为3个部分[1]:前外侧PE区(PEla区)、外侧PE区(PEl区)和内侧PE区(PEm区)。PEla在区域PEl的前面形成长条;PEl在SPL的裸露表面上可见,为顶内沟边缘上的一条相对较细的条带,并一直延伸到PE内侧壁的外部顶内沟区域;PEm是位于背侧的皮质区域。PEla和PEl似乎与Brodmann5区显示的体位图重叠[7],而PEm区域仍需要进行功能表征。然而,解剖规范定义的PE区及其亚区还未应用于影像连接分析。利用脑连接信息实现PE的边界识别和PE的精细划分,并刻画这些脑区的功能和解剖连接模式,可以为研究PE结构和功能做导航。另外,目前有关PE的认知大多源于动物实验,这些认知是否可以直接转化到人类犹未可知。
大脑白质纤维束研究一直是脑科学研究中的热点。其中,弥散张量成像所体现的白质纤维轨迹具有明确的神经学意义。它可以获得每一个体素从出发到跟踪终点的白质纤维路径,这被称作该体素的解剖连接。每个体素的解剖连接可以作为特征来进行分区。当前有很多基于解剖连接模式对大脑分区的研究,通过白质纤维连接的追踪可以构建更加精细的脑区亚区。此外,也有研究基于同源脑区进行概率纤维追踪构建连通性指纹,通过分析其相似性来探讨物种间的同源关系。
综上,本文基于解剖连接模式,采用概率纤维追踪的方法,面向弥散张量成像数据进行猕猴PE区域的分割及验证,进而对人类大脑Brodmann 5区和猕猴该区域展开跨物种比较。为PE亚区绘制了具有19个预定义同源目标脑区的连通性指纹图,找到了所划分出的PE亚区在人类大脑中的同源区域,对该区域连接模式上进行初步探索。通过以上方法所得到的分割结果能够更好地理解PE区的功能组织,并为其在亚区层面上的信息交互和整合提供新的见解。对猕猴模型相关研究的精准展开和对人类特有高级运动功能的探索具有重要的理论意义和应用价值,有助于影像连接分析和神经外科手术定位研究,为人类手臂运动临床诊断和康复训练提供依据。
1 实验材料及方法
1.1 数据采集和预处理
1.1.1人类数据
本文使用Human Connectome Project(HCP)发布的公开数据集[9]。经过数据清洗,去除含极端值或缺失值的数据,选取了22位健康的被试(13位男性,9位女性,年龄范围22~35岁),分别选择他们对应的预处理后的结构磁共振成像(structural MRI,sMRI)数据和弥散磁共振成像(diffusion MRI,dMRI)数据,其中sMRI数据包括0.7 mm高分辨率的各向同性T1加权(T1 weighted,T1w)和T2加权(T2 weighted,T2w)图像数据,dMRI数据包括多梯度和多弥散加权的1.25 mm各向同性数据。HCP的核磁共振数据用3.0T磁共振系统获得。弥散磁共振成像数据采集参数TR=5 520 ms,TE=89.5 ms,体素分辨率1.25 mm×1.25 mm×1.25 mm,无间隙。此外还采集了矢状位3D T1加权图像,采集参数TR/TE=2 400/2.14 ms,体素分辨率0.7 mm×0.7 mm×0.7 mm.该研究符合赫尔辛基宣言的最新版本。
1.1.2人类数据预处理
公开的HCP数据已经过预处理[10]。接下来,人类大脑采用基于FSL(FSL-FslWiki (ox.ac.uk))的两步配准法进行配准,并选取配准效果较好的个体组成实验的数据集,如图1所示。使用FMRIB提供的BEDPOSTX(双张量模型)[11]对两个纤维方向的弥散概率追踪成像和体素概率分布进行估计,以便执行概率纤维跟踪实验。之后,根据中科院自动化所提供的人类脑网络组图谱(Brainnetome Atlas)定义人类的5l区和5m区见图2,基于图谱配准到个体的结果,按照对应脑区标号进行区域提取,得到人类被试左右半球5l区和5m区。
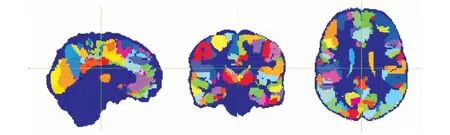
图1 Brainnetome图谱到人类个体大脑的配准结果Fig.1 Result of registration from Brainnetome atlas to individual human brain

图2 人类大脑5l区和5m区Fig.2 Area 5l and area 5m of the human brain
1.1.3猕猴数据
本文使用The PRIMatE Data Exchange(PRIME-DE)中的公开数据集[12]。采用同样的入组标准,选取来自加州大学戴维斯分校(University of California, Davis,UC-Davis)的9只雌性猕猴(年龄范围18.5~22.5岁,体重7.28~14.95 kg)作为被试,猕猴数据包含T1结构像和弥散磁共振成像。这些猕猴数据使用西门子Skyra 3T的4通道翻转线圈扫描收集得到。弥散加权像数据采集参数TE=115 ms,TR=6 400 ms,切片间隙为1.4 mm,b值为1 600 s/mm2,800 s/mm2,体素分辨率为1.4 mm×1.4 mm×1.4 mm.T1结构像数据采集参数TE=3.65 ms,TR=2 500 ms,TI=1 100 ms,翻转角度为7°,体素分辨率为0.3 mm×0.3 mm×0.3 mm.饲养的猕猴符合UC-Davis IACUC道德认证,且数据采集实验得到了当地伦理委员会的批准,并遵守欧盟关于保护用于科学目的的动物的指令(2010/63/EU).
1.1.4猕猴数据预处理
本文使用FSL工具对猕猴弥散磁共振成像数据进行了预处理。处理过程包括头动和涡流校正,颅骨剥离等。使用FSL工具进行两步配准,并选取配准效果较好的个体组成实验的数据集如图3所示。使用BEDPOSTX进行纤维方向估计。之后,根据D99图谱[13]定义猕猴PE区如图4所示,进行区域提取后得到猕猴被试左右半球PE区。

图3 D99图谱到猕猴个体大脑的配准结果Fig.3 Registration results of D99 atlas to individual macaque brain
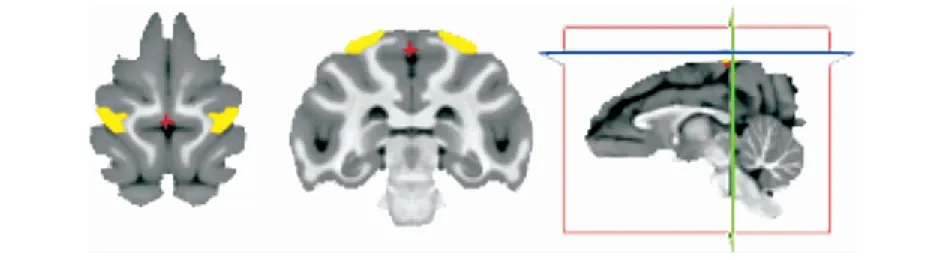
图4 猕猴大脑PE区Fig.4 Area PE of macaque
1.2 概率纤维追踪
PE分区实验中,对所选被试的PE区进行概率纤维追踪,估计每个种子体素和大脑其余部分中的每个体素之间的连通概率,得到PE对全脑的解剖连接矩阵。接下来,通过将连通性矩阵乘以其转置矩阵生成对称互相关矩阵,第(i,j)个元素值是种子体素i的连通性分布和种子体素j的连通性分布之间的相关性[14]。
本文在通过文献总结出已有人类与猕猴的同源区[15]的基础上,猕猴基于D99图谱选区,人类基于Brainnetome图谱找对应区域,最终选用主要运动皮层、主要躯体感觉皮层、辅助运动皮层等19个同源目标区域见表1.使用FSL对人类5l区与5m区到目标大脑区域进行概率纤维追踪,对猕猴PE区及其亚区到目标大脑区域进行概率纤维追踪,以映射区域每个体素和目标大脑区域之间的解剖连通性。最后,确定每个区域到每个目标脑区的解剖连接概率。
2 划分PE亚区及跨物种比较
2.1 分区
2.1.1分区过程
本研究使用边加权谱聚类对相似性矩阵进行处理,以确定2~5个不同数量的亚区聚类[16-21]。边加权谱聚类是把谱聚类和体素间的概率连接信息综合起来考虑,将具有相似全脑连接构筑特征的体素聚在一起,得到的聚类结果散点较少,此方法和K-means聚类、谱聚类等聚类算法比起来可以得到更加平滑的聚类边界,且计算速度快。

表1 本文所使用的人类与猕猴的同源目标区Table 1 Homologous target areas of human and macaque used in this article
接下来,将被试的个体分割结果从弥散空间转换到D99空间,为所有对象的每个解创建最大概率图(maximum probability map,MPM).MPM使用标准空间中所有被试的分割结果,通过将参考空间中的每个体素分配给最有可能定位的区域来计算[22-23]。最后对聚类结果进行中值滤波,配回到D99模板上,并将PE亚区配到D99图谱上。
2.1.2分区验证
Dice系数用于评估基于解剖连通性的分组结果之间的重叠程度,以识别PE的相应拓扑结构,从而确定PE中亚区的最佳分组结果[24]。Dice系数值越大说明PE分割效果越好。同时使用平均Crame’s V(CV)来测量个体之间区域的空间分布一致性,最终取CV值接近1的最佳分区方案。另外,使用标准化互信息(Normalized Mutual Information,NMI),通过相邻分区方案的比较来确定最稳定的分区方案。最后,使用信息差异指标(Variation of Information,VI)量化两个分区方案之间信息量的丢失或获取情况,来确定最稳定的分区方案。
2.2 跨物种比较
2.2.1连通性指纹
为了进一步评估人和猕猴之间的解剖连通指纹,每个感兴趣区域的解剖连通指纹在所有目标上进行最大最小归一化,使得感兴趣区域和任何目标大脑区域之间的最大连接概率值为1,最小连接概率值为0,公式如下:
(1)
其中,pi,j是第i个感兴趣区域和第j个目标脑区之间的连通概率。max(pi)是第i个感兴趣区域与所有目标脑区之间的最大连通概率,而min(pi)是第i个感兴趣区域与所有目标脑区之间的最小连通概率。归一化的解剖连接概率值用于构建每个感兴趣区域的连通性指纹。最后,为了定量表征人类和猕猴之间的相似性,计算了人类和猕猴指纹之间的余弦相似度和曼哈顿距离。
(2)
其中,p、q表示两个待比较的指纹图;n表示指纹图中目标区域个数;pi、qi表示p、q指纹图中第i个目标区域的连通值。余弦相似度计算所得到值的范围为[-1,1]:值越接近1,表明两个脑区的连通性越相似,即两个脑区同源的可能性越大;值越接近0,表明两个脑区相互独立,同源的可能性越小;值接近-1,表明两个脑区负相关。
(3)
式中:p和q是连通指纹的两个向量,i是指纹的n个目标区域。曼哈顿距离的值越小表示两个向量的相似性越高,表明两个脑区的连通性越相似,即两个脑区同源的可能性越大。
2.2.2置换检验
在基于连通性指纹的跨物种比较过程中,使用非参数置换检验来探测猕猴PE区、PE亚区与人类的5m区、5l区对应连接指纹图的汇聚性:零假设定义为“对应指纹图具有汇聚性”;检验标准值定义为5%显著水平。
1) 首先计算实际观测值为组平均指纹图(人类22被试;猕猴9被试)之间的余弦相似度值。
2) 组合两组指纹图然后随机分开成22和9两个组。
3) 计算新的两个组的组平均值指纹图及它们之间的余弦相似度值。
4) 重复执行第2),3)步1 000次,使用得到的余弦相似度值构建置换检验分布。
本文通过判断实际观测值在检验分布中是否为异常值来确定零假设是否成立。若实验的观测值落在置信区间内,则原假设成立,即所检验的脑区间有同源关系。
3 结果与讨论
3.1 猕猴PE的聚类分区
本文基于解剖连通性获得了猕猴PE区分别聚类成2、3、4、5个亚区的MPM见图5(a).计算4项分区指标,结果见图5(b).显示当分为2个亚区时,左脑CV=0.521、Dice=0.416、NMI=0.546,3个指标的值都大于分成其他数量的对应指标值,同时信息差异VI指标也是在分成两个区时值最小,右脑分区呈现的结果与左脑一致,因此最终得到2个亚区的最佳分组结果。其中,在D99模板里前侧亚区的位置为L1[95,128,201]、R1[179,134,201],背侧亚区的位置为L2[86,134,201]、R2[187,126,201].因此,在随后的分析实验中使用2个亚区分组,并将亚区配准到D99模板,如图5(c)所示。
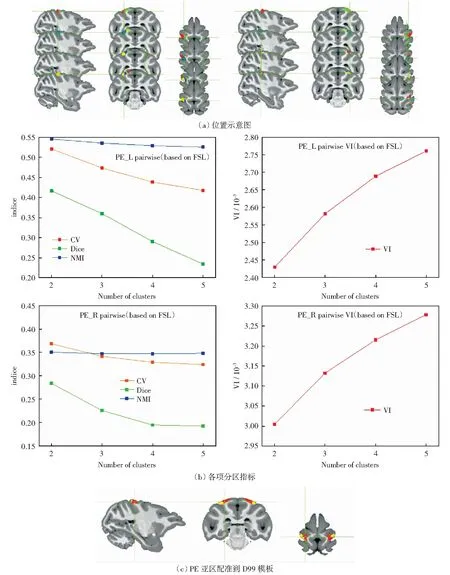
图5 基于FSL两步配准的猕猴左右脑PE细分结果Fig.5 Results of PE segmentation of the left and right brains of macaque based on FSL two-step registration
3.2 猕猴PE区的跨物种比较
基于连通性指纹的比较方法将两物种大脑带入到同源脑区构建出的公共空间如图6.连通性指纹和余弦相似度如表2所示。可发现,整个PE区更接近人类5l区,同时根据趋势图也可看出猕猴PE区与人类5l区趋势一致性较强。本文发现PE区与5l区均对pIPS和S1表现出了强烈的连接性。pIPS在人脑中的位置同样位于SPL,它在猕猴大脑中对应区域是5_(PEa),5_(PEa)本身与PE相邻,S1是人类和猕猴的初级体感皮质区[26, 27].
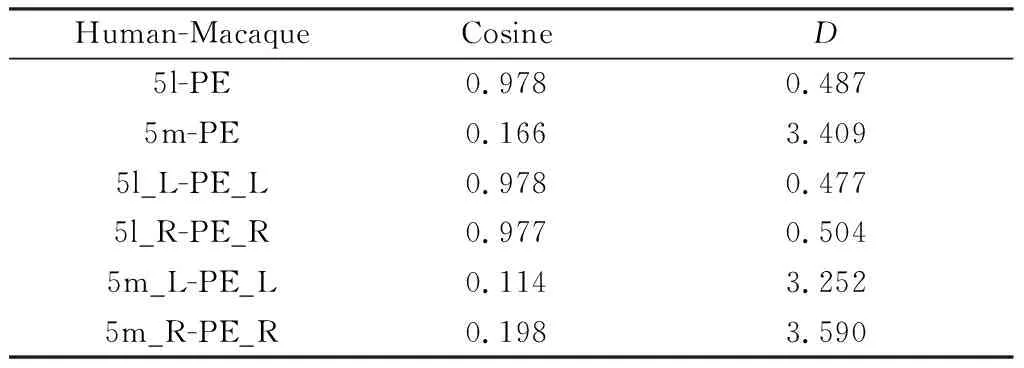
表2 人类5l区、5m区与猕猴PE区相似性度量Table 2 Similarity measure between 5l, 5m of human and PE of macaque
根据置换检验结果,左右脑对应的5l和PE,观测值均落在阈值内,因此本文判定他们具有同源关系。而左右脑对应的5m区和PE区在置换检验中观测值落在了阈值外,因此对于此前实验中它们相似度低的结论,可能是受样本量小的影响。
3.3 猕猴PE亚区的跨物种比较
图7为猕猴PE亚区的跨物种比较结果。表3为相似性度量。从结果可以发现(见图7和表3),PE_L1与5l_L、 PE_R1与5l_R相似度更高,认为PE _L1和PE_R1与5l具有同源关系。5m与PE亚区在本次实验中并未表现出明显的同源关系,这和PE整区结果一致。另外发现除了pIPS和S1,PE亚区也表现出对M1的强连接,M1是人类和猕猴的初级运动皮质区[27]。
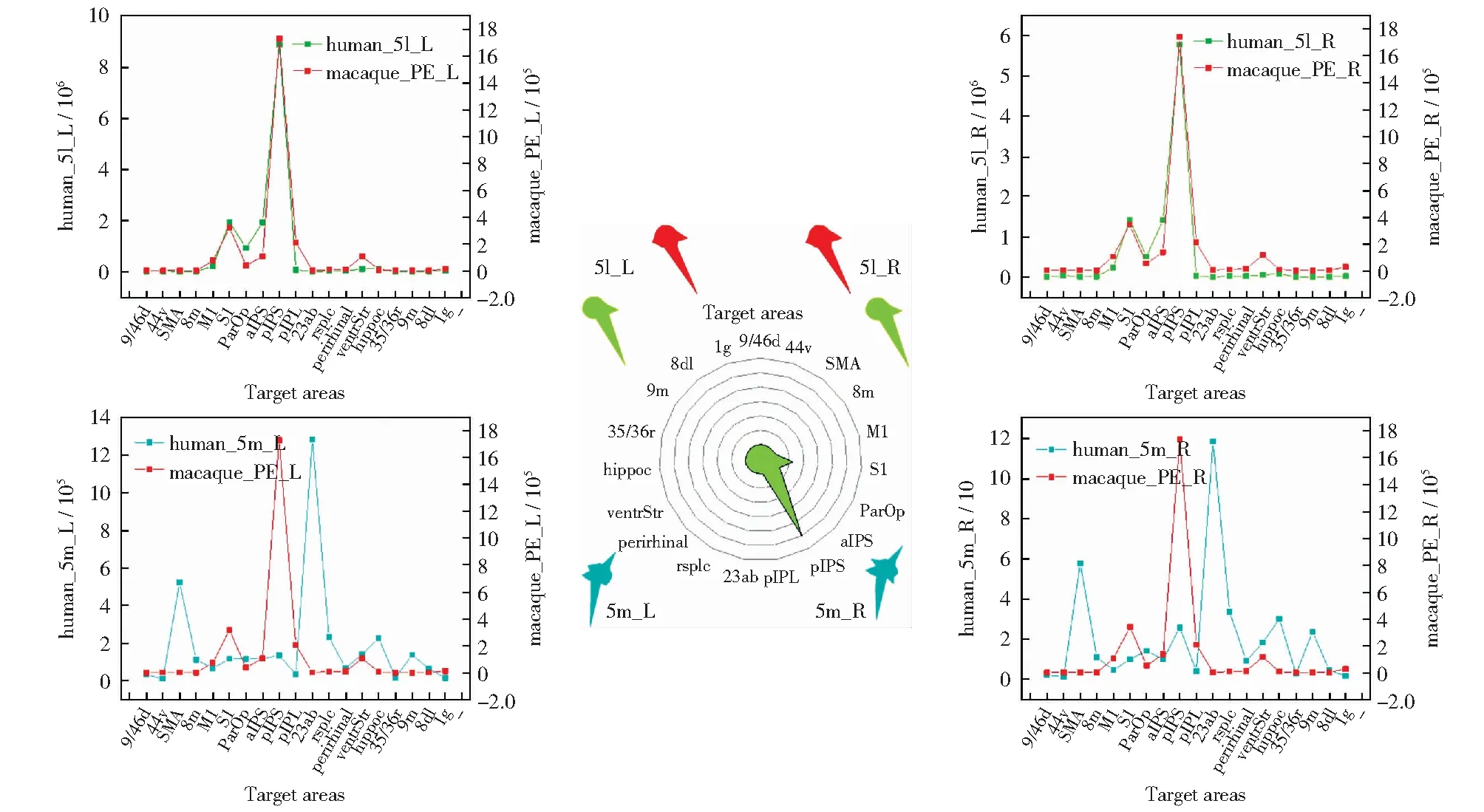
图6 人类5l区、5m区与猕猴PE区同源性比较Fig.6 Comparison of homology between 5l, 5m of human and PE of macaque
4 结束语
本文基于解剖连接,面向弥散张量磁共振成像数据,对白质纤维束追踪结果运用边加权谱聚类方法,进行猕猴PE区的分割及验证,并对PE区及其亚区进行了跨物种比较研究。结果发现PE可细分为一个前侧PE亚区和一个背侧PE亚区。通过绘制人类与猕猴的连通性指纹并计算余弦相似度得出,猕猴PE区与人类5l区具有相似的连接模式。本文所划分的亚区相对于5m区来说,与人类5l区具有更为相似的连接模式,且推断PE前侧亚区与5l同源。同时根据位置及实验得出的同源关系,本文推测PE前侧亚区对应之前基于细胞构筑得出的细分亚区PEla和PEl,PE背侧亚区对应之前基于细胞构筑得出的细分亚区PEm.根据实验结果, PE与5l显示出与pIPS、S1、M1的强连接性,进一步验证了解剖连接可以在一定程度上反映功能连接关系。此外,由于PEm是位于背侧的皮质区域,没有像PEl与PEla一样裸露在表面,因此本文猜想实验中该部分的相关结果也受图像的影响,另外前人对于PEm与5m同源的推断还需进一步验证。上述结论进一步验证了该区人类与猕猴间的同源关系,也提示本文对于小区域精细划分的精度还有提升的空间。此外,本文只基于白质纤维束的解剖连接来进行分区,未来研究将关注PE的多模态划分,同时进一步研究亚区的功能结构,进而探究人类和猕猴的同源关系,以促进该领域未来的临床、认知和进化研究。

图7 人类5l区、5m区与猕猴PE区亚区同源性比较Fig.7 Comparison of homology between 5l, 5m of human and PE subregions of macaque
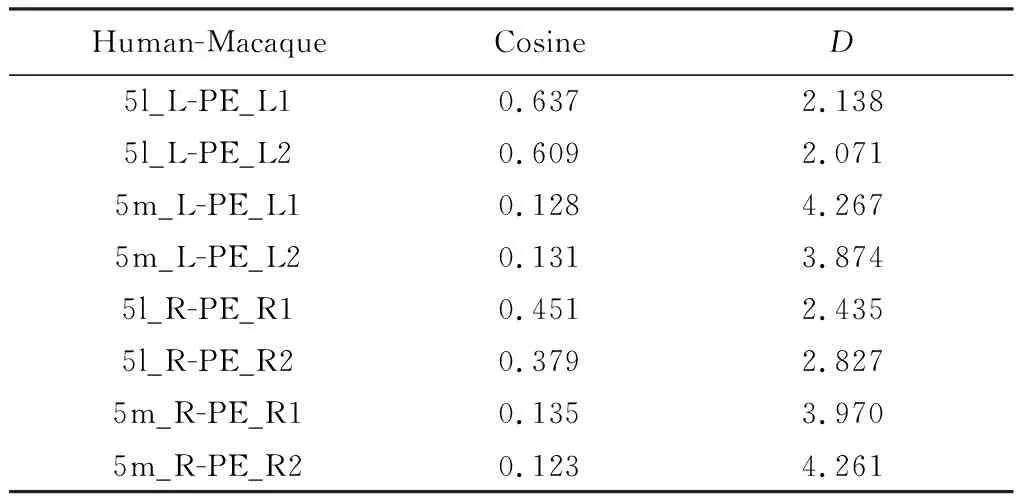
表3 人类5l区、5m区与猕猴PE亚区的相似性度量Table 3 Similarity measure between 5l, 5m of human and PE subregions of macaque
