基于PAE编码系统对武陵洞蛭行为谱的初步研究
2022-09-13伍丽艳任伯凇严思思刘志霄
伍丽艳,刘 力,任伯凇,严思思,刘志霄
(吉首大学生物资源与环境科学学院,中国湖南 吉首 416000)
动物行为是动物所做的有利于眼前自身存活和未来基因存活的一切事情,或者说是在个体层次上,动物对来自体内的生理变化和来自体外的环境变化所做出的整体性反应[1]。研究动物行为可揭示动物活动规律、探索动物行为调节对环境的适应及动物与环境之间的关系[2]。研究动物行为的首要步骤是将所研究动物的各种行为,进行真实的观察记录、详细的辨析归纳、定义和描述[1,3],并按照一定的组织构建行为目录。行为谱(ethogram)是指研究对象近乎全部所能表现出来的行为目录[4]。行为谱编制作为动物行为研究的基础性工作,为进一步深入研究动物行为的机制、功能与进化等问题提供了前提和条件[5~6]。编制一套完整的行为谱对动物行为学的定量研究尤为重要[7]。
行为谱这一概念最早由Makkink于1936年提出,随后国内外学者参考该方法对大量的动物行为构建了“描述行为谱”,其不仅包括人工环境下的动物行为[8~14],也包括自然环境下的动物行为[15~16]。在所建构的动物行为的描述行为谱中,有学者将描述行为谱与行为绘画相结合[16~18];也有学者对动物行为进行层次划分,并根据行为的生态学意义对行为进行分类[19];同时还有学者对特定环境条件下动物某些特殊行为的发生频次及活动节律进行研究[20~21]。
2000年,蒋志刚[22]则以麋鹿为实例,通过辨识行为的基本单元,区别了“姿势”“动作”“环境”三要素,分解了动物行为的层次,然后根据行为的适应和社群机能进行归类,建立了以“姿势-动作-环境”为轴心、以生态功能为分类依据的动物行为分类编码系统(PAE编码系统)。之后,国内出现了许多PAE编码行为谱,其主要涉及哺乳纲的灵长目[23~30]、食肉目[31~33]、偶蹄目[34~38]、鲸目[39~40],以及鸟纲[41~47]。鱼纲[48]和两栖纲[49]也有研究报道。
迄今,对于无脊椎动物PAE编码行为谱的研究极为罕见,尤其是环节动物门的PAE行为谱尚缺乏研究报道。
武陵洞蛭(Sinospelaeobdellawulingensis)隶属于环节动物门、蛭纲、无吻蛭目、山蛭科、中国洞蛭属(Sinospelaeobdella),目前仅发现于湖南省湘西土家族苗族自治州(简称湘西州)吉首市、古丈县和永顺县境内的少数几个溶洞中,以及四川省邻水县的老龙洞内[50~51]。中国洞蛭属是2019年1月由刘志霄教授课题组所发表的一个新属[50]。与人们所熟知的水蛭和山蛭不同,洞蛭栖息于溶洞的顶壁,吸食洞栖性蝙蝠的血液,并在洞顶壁完成其生活史,已演化成为真洞穴动物(troglobites),因此对其PAE行为谱进行研究有助于深入探究其行为生态学特征及进化适应意义。
1 研究方法
1.1 研究样洞及时间
2019年6月—2021年4月,选择吉首市堂乐洞、水牛洞和旧寨坪岩洞作为主要观察样洞,同时把永顺县金鸡洞、黄泥洞以及古丈县的鸡公洞作为参考样洞进行野外观察与数据采集。
1.2 野外观察
研究期间,3~4人一组进入样洞,主要采取随机取样和扫描取样的方式观察并记录洞蛭的各种行为,以初步明确各类行为的发生频率,包括多发行为、明显行为和少见行为等。之后,采用目标取样和行为取样法观察具有代表性的特定研究对象的特定行为,用自制的行为观察描述表对行为过程及行为模式进行描述和记录,采用连续记录法对行为发生的频次和持续时间进行记录。
除了用笔和纸实时观察记录外,为了对武陵洞蛭(以下简称洞蛭)明显具有生理或生态功能的行为进行准确定义和描述,我们采用佳能全画幅专业单反EOS 5D MarkⅣ机身和佳能EF 24~70 mm f/2.8L镜头对洞蛭的行为进行摄录。
为了对洞蛭觅食行为、运动行为、寄(宿)主专一性等生物学特性进行研究,我们将小泡巨鼠(Leopoldamysedwardsi)(在洞内捕获的)、家鼠、家兔、家鸽、蝙蝠等脊椎动物作为受试物种,将其手托或装在套袋中,再将套袋固定在有洞蛭分布的岩壁上,观察并记录洞蛭对受试动物的觅食情况,同时观察洞蛭对人体手部皮肤的敏感性及吸血情况。
1.3 室内研究
对野外拍摄到的照片和录像进行反复观看与比较,提取并分析行为要素。期间,也将少量代表性的个体带到室内,在实验室条件下进一步地观察洞蛭的行为特征,以补充、修正野外观察到的行为要素与行为模式。
2 结果与分析
2.1 行为的姿势编码
通过反复观察和分析,共分辨和记录到洞蛭的5种基本姿势(图1):卧、倒挂、贴、爬、立。其中,卧、倒挂、贴属于静态姿势;爬、立为动态姿势。各姿势的编码如表1所示。

表1 武陵洞蛭的基本姿势及其编码(P码)Table 1 Basic postures and their codes of S.wulingensis

图1 武陵洞蛭的5种基本姿势(A)卧;(B)倒挂;(C)贴;(D)爬;(E)立。C图中一只洞蛭的口吸盘正贴附于一只蝙蝠的足趾间;D图是洞蛭在尺蠖状爬行过程中,后吸盘被放置在口吸盘的后面,使得身体呈极度弯曲时的状态。Fig.1 Five basic postures of S.wulingensis(A)Lying;(B)Hanging upside-down;(C)Attaching;(D)Crawling;(E)Erecting.In photo C,the oral suction cup of S.wulingensis is attached between the toes of a bat.In photo D,the posterior sucker is placed behind the oral sucker in the inchworm crawling process of the cave leech,making the body extremely bent.
2.2 行为的动作编码
按照洞蛭动作发生部位的不同(头部、体部、肛门部、尾吸盘),初步分辨和记录到22种动作。这些动作的编码如表2所示。

表2 武陵洞蛭的基本动作及其编码(A码)Table 2 Basic acts and their codes of S.wulingensis
2.3 洞蛭行为发生的环境及编码
此次研究共分辨和记录到洞蛭行为发生的环境11种,包括生物环境和非生物环境,其中生物环境6种,非生物环境5种。这些环境的具体编码如表3所示。

表3 武陵洞蛭行为发生的环境及其编码(E码)Table 3 Behavioral environments and their codes of S.wulingensis
2.4 洞蛭行为的PAE编码
将所有观察到的行为划分为5大基本类型:休息行为、运动行为、觅食行为、生殖行为和排遗行为(表 4)。
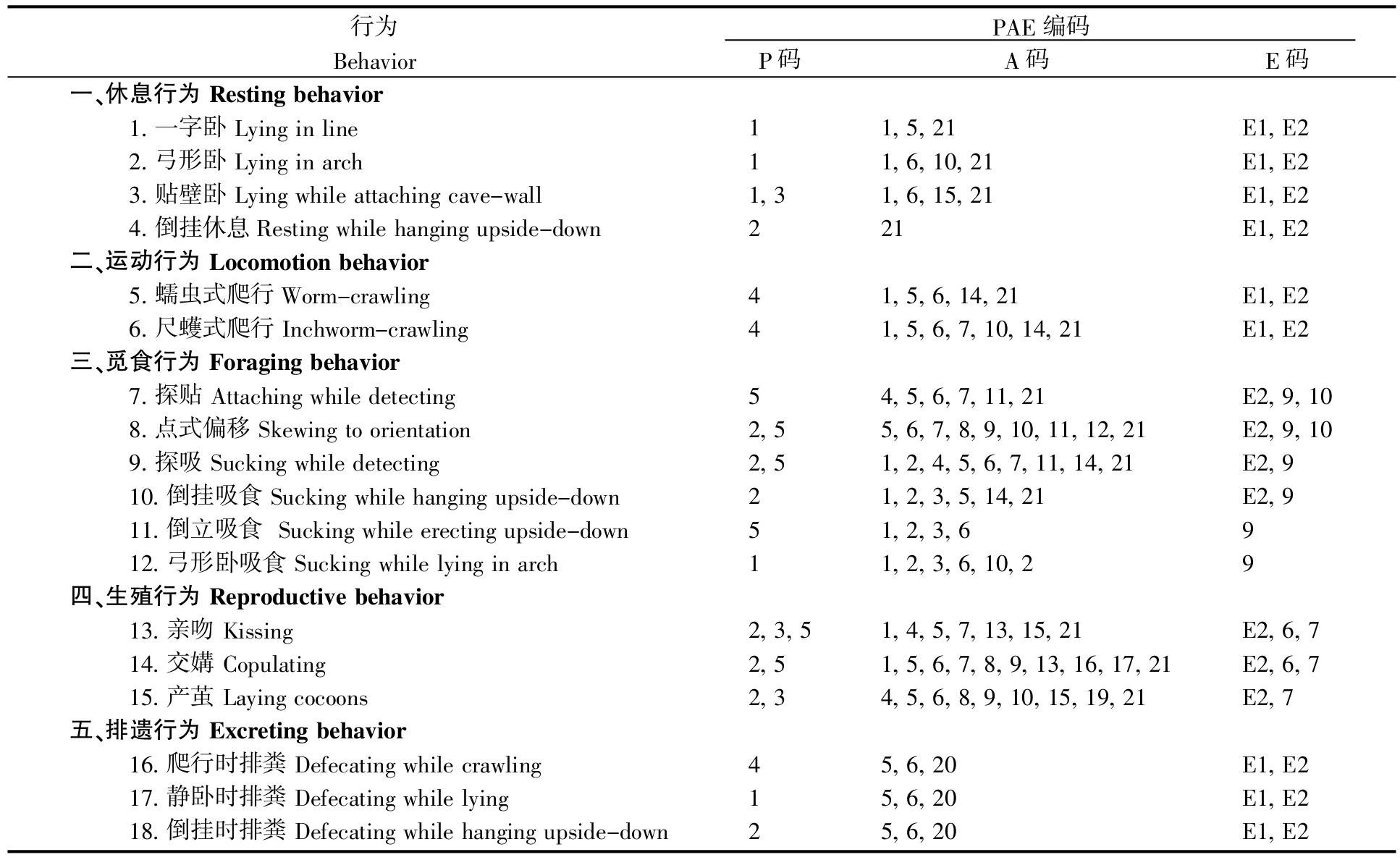
表4 武陵洞蛭行为的PAE编码系统Table 4 PAE coding system of behaviors for S.wulingensis
休息行为是指洞蛭在某种环境中身体呈放松状态并维持某种姿势不变的现象(图2)。洞蛭的休息行为主要表现为静卧休息和倒挂休息这两种基本方式,前者又包括一字卧、弓形卧和贴壁卧。一字卧:洞蛭口、尾吸盘固着,蛭体伸展且呈现“一”字型。弓形卧:洞蛭口、尾吸盘固着,蛭体缩短且腹向弯曲为弓形。贴壁卧:洞蛭口、尾吸盘固着,蛭体缩短且贴在洞壁上,蛭体形状因吸盘吸附位置和支持物形状的不同而各有差异,此类静卧较为常见。倒挂休息:洞蛭仅尾吸盘固着,蛭体因自身的重力而指向地心。

图2 武陵洞蛭的休息行为(A)一字卧;(B)弓形卧;(C)贴壁卧;(D)倒挂休息。Fig.2 Resting behaviors of S.wulingensis(A)Lying in line;(B)Lying in arch;(C)Lying while attaching cave-wall;(D)Resting while hanging upside-down.
运动行为是指洞蛭改变自身位置的过程,主要包括蠕虫式爬行和尺蠖式爬行(图3),后者更为常见。蠕虫式爬行是指处于静卧状态的洞蛭,其口吸盘松脱,蛭体向前端伸展,随后口吸盘吸附,蛭体体节由前端往后端逐步缩短,产生拉力,使尾吸盘松脱,并拉引蛭体后端向前,然后尾吸盘吸附,再重复这一动作程式,使身体发生位置变化。尺蠖式爬行是指处于静卧状态的洞蛭,其口吸盘松脱,蛭体向前端伸展,然后口吸盘吸附,蛭体前段的体节快速缩短产生拉力,使尾吸盘松脱,蛭体后段因拉力而抬高,接着蛭体腹向弯曲使尾吸盘紧靠在口吸盘的后面吸附,之后口吸盘再松脱,蛭体向前伸展,如此反复动作,身体发生位移。

图3 武陵洞蛭的运动行为(A)蠕虫式爬行;(B)尺蠖式爬行。Fig.3 Locomotion behaviors of S.wulingensis(A)Worm-crawling;(B)Inchworm-crawling.
觅食行为是指洞蛭为维持自身新陈代谢和生长发育的需要,在周围环境中探测、搜寻、抵近和摄取食物的姿势与动作过程(图4)。洞蛭觅食行为的基本程式如下:
1)对可能的宿主动物(食源刺激)到来的初始反应:当可能的宿主动物,如蝙蝠、鼠、鸽、兔或人等,在一定范围内靠近洞蛭时,洞蛭可感受到相应的刺激,通常呈静止状态的洞蛭对这类刺激的反应较慢,表现出身体前段伸展,然后抬起。
2)对可能的宿主动物的定向定位探测运动:当感受到可能的宿主动物的刺激而又不知其方向和位置时,洞蛭会以身体的前段进行探贴,即洞蛭用口吸盘贴附于支持物的表面,随即抬起蛭体的前段,然后洞蛭对支持物表面的其他位点重复以上动作。随着可能的宿主动物的逐渐靠近,刺激越来越强烈,这时洞蛭的身体开始摆动并向周围的各个方位作点式偏移,最后在一个确定的方位进行探贴,并用口吸盘吸附在支持物的表面,这表明洞蛭已经感觉到了可能的宿主动物的方位。
3)向可能的宿主动物爬行:口吸盘吸附于支持物的表面以后,洞蛭随即向可能的宿主动物的方向爬行,主要是尺蠖式爬行。在爬行过程中,洞蛭会抬起蛭体的前段进行探贴、摆动和点式偏移以对可能的宿主动物重新定位,确定下一处爬行点。
4)对可能的宿主动物进行识别:当爬至可能的宿主动物的躯体周围时,洞蛭会以尾吸盘牢牢地固着于支持物上,并以探贴的方式对宿主动物进行识别。
5)寻找适合的位置,咬住后吸血:以洞蛭对蝙蝠的探吸为例,其用口吸盘不断地在蝙蝠体表的不同位置进行试探,有时吸吮,有时不吸吮,有时吸吮片刻又脱开,另找位置再试探,最后多数是在蝙蝠的足趾关节或者翼膜等皮薄毛稀、毛细血管丰富之处,咬开皮肤吸血。在溶洞内,我们仅观察到洞蛭的倒挂吸食、倒立吸食和弓形卧吸食3种吸血行为(图 4)。

图4 武陵洞蛭的觅食行为(A)探贴;(B)点式偏移;(C)探吸;(D)倒挂吸食;(E)倒立吸食;(F)弓形卧吸食。Fig.4 Foraging behaviors of S.wulingensis(A)Attaching while detecting;(B)Skewing to orientation;(C)Sucking while detecting;(D)Sucking while hanging upsidedown;(E)Sucking while erecting upside-down;(F)Sucking while lying in arch.
生殖行为是指洞蛭在繁殖期发生的与繁殖直接相关的动作过程与活动,包括亲吻、交媾、产茧等。
亲吻行为是指两条洞蛭的尾吸盘固着,蛭体悬空且其环带区的前段腹面相贴,两口吸盘相互贴近,一前一后互抵,此运动是交媾行为的前奏(图 5A)。
交媾行为是指两条洞蛭的尾吸盘固着,蛭体悬空且其环带区的前段腹面相贴,以口吸盘互吻一段时间之后,两者的头部呈钩状,使身体的前段反向缠绕,并以环带区的腹面相贴互抵,将阴茎相互插入对方的阴道内,把精子输送给对方(图5B)。
产茧期是一个非常重要的时期,产卵行为极其有意思且复杂。洞蛭的尾吸盘固着,产卵时先由环带区分泌黏液,黏液分泌时因掺入空气而呈肥皂泡沫状,随着黏液分泌物的增多,这些泡状物向两侧背移动包住身体且粘住支持物,随后环带区分泌另一种黏液,形成一层卵袋包于环带周围,接着再分泌黏液形成另一层卵袋,然后雌孔产出一些黄色、较浓、含有受精卵的黏液,黏液落入卵袋和身体之间的空腔内。排完卵后的洞蛭,其身体的前段后缩,在退出卵袋的过程中,以口吸盘腺体分泌形成的栓塞住卵袋的前后两个开孔。刚产出的卵茧大致呈球形,外围透明呈泡沫状,里面可见白色卵袋(图5C),在受精卵孵化的过程中,卵茧的颜色逐步加深变为淡黄色或黄褐色,幼体孵出后,卵茧逐渐萎缩,呈现枯黄状。

图5 武陵洞蛭的繁殖行为(A)亲吻;(B)交媾;(C)刚产下卵茧后正在离开的洞蛭。Fig.5 Reproductive behaviors of S.wulingensis(A)Kissing;(B)Copulating;(C)A cave leech leaving after laying a cocoon of eggs.
排遗行为是指洞蛭将经消化道消化、吸收之后的食物残渣以粪便的形式排出体外的过程,包括爬行时排粪(图6)、静卧时排粪和倒挂时排粪3种排粪行为。

图6 武陵洞蛭的爬行时排粪行为Fig.6 Defecating behavior of S.wulingensis while crawling
3 讨论
中国洞蛭属是近年首先在湘西州境内发现的一个新属,其模式种为武陵洞蛭[50~51]。该新属种栖息于黑暗潮湿的溶洞顶壁,以暂时性吸食蝙蝠的血液(简称吸蝠血)来维持种群的繁衍,因此其行为生态学特征具有重要的蛭类学和洞穴生物学研究意义[52]。
与行为描述、行为绘制等传统的行为谱研究方法相比,以PAE编码系统构建动物的行为谱可操作性强,不仅能够客观系统地记述动物的行为要素与行为类型,而且PAE行为谱的可检验度高,便于不同类群之间行为系统与行为发育方面的比较研究,有助于理解和掌握动物行为系统的发育规律。
但是,任何一种动物PAE行为谱的制订都比较耗时费力,研究人员必须仔细观察、反复验证,才可能构建全面而准确的PAE行为谱,而且编码系统的各个元素要求具有独特性或不可替代性。此外,PAE编码系统制订之后,如果作者或其他学者又发现了该种动物行为新的姿势、动作和环境,还须要对整体行为谱编码进行调整、修改和补充。
基于对PAE编码系统的理解和对武陵洞蛭行为的反复观察,本文首次构建了洞蛭的PAE行为谱,这也是首次采用PAE编码系统研究蛭类及环节动物门的行为系统,为今后蛭类及其他环节动物行为的比较研究提供了基本的参比资料。
当然,限于时间、精力及条件,有些行为(如幼蛭从卵茧中孵出时的行为),以及行为的节律性(如日活动节律、季节活动节律和年活动节律),还有待今后进一步的观察与研究。
行为学研究的首要工作是对观察到的行为加以描述和定义,只有对行为进行准确的描述和定义之后,才能进一步研究行为背后的机制、功能和演化等问题[53]。而行为进化的一些重要发现来自对行为个体发育的研究。幼年动物的行为往往类似于或等同于一个较原始物种成年个体的行为,就好像它们是该物种的祖种一样[1]。
本研究发现,武陵洞蛭幼体的某些行为较之成体(指具有环带的繁殖体)有较多的不同。在运动行为方面,成体的尺蠖式爬行是借助身体前端至后端逐步收缩产生拉力,将后吸盘从被固着物上拔起,然后借助产生的惯性和肌肉的辅助作用使蛭体抬起,同时腹向弯曲;而幼蛭的尺蠖式爬行是借助肌肉收缩提拉蛭体的后端,并通过背向弯曲使蛭体的后段相对前移,再腹向弯曲。产生这种差异的原因可能是幼蛭的吸盘尚在发育之中。我们推测,洞蛭发达的尾吸盘是适应于洞顶壁特殊的栖息行为(面临着“倒悬身体负重运动”的选择压力)而逐渐衍生的,幼蛭可能存在运动行为模式与尾吸盘发育的相关性问题,而这种相关性可能具有形态及行为生态方面的进化适应意义。
幼蛭的觅食行为与成体也不尽相同。首先,幼体与成体吸蝠血的姿势和动作不同。多数成体是爬上蝙蝠的翼膜或足趾间吸血,但幼体吸血时,尾吸盘牢牢地吸附在洞壁上,同时尽量使蛭体伸长,以口吸盘吸附在蝙蝠的身上。其次,在幼体与成体的吸蝠血过程中,被吸血的蝙蝠的行为表现不同。幼体吸血时,蝙蝠的反应较为强烈,出现用后足踢动、抓挠幼蛭的行为,有时还可见其将后足伸至口部,用舌头舔舐后足的行为,因此幼蛭的吸血过程被多次中断,每次的吸血时间较短;而成体吸血时,从外部反应来看,蝙蝠似乎感受不到其自身正在被吸取血液。
已知吸血蛭类可通过口吸盘中的腺体分泌活性物质来麻醉寄主,同时产生抗凝血作用,但从幼蛭吸血过程中蝙蝠的外部行为表现来看,我们推测:洞蛭在幼体阶段吸血时尚不能使寄主麻醉,这意味着洞蛭口吸盘中的腺体结构及功能可能是在其从卵茧中孵出以后才逐步发育形成的。
此外,我们在观察中发现,洞蛭似乎仅对蝙蝠比较敏感,并嗜吸蝙蝠的血液,对人体皮肤及其他动物不敏感或不感兴趣。当然,有关洞蛭是否存在吸蝠血专一性及其可能的进化适应机制,我们还须从行为生态、生理生化、分子遗传及适应性进化等方面进行综合性的探究。
致谢:本研究得到吴涛、黄太福、龚小燕等学长多方面的帮助,谨此致谢。
