冷藏期间滩羊肉保水性变化的机制
2022-09-01刘吉娟罗瑞明李子欣尤丽琴
刘吉娟,杨 波,罗瑞明,李子欣,尤丽琴
(宁夏大学食品与葡萄酒学院,宁夏 银川 750021)
保水性是衡量滩羊肉品质的重要指标之一,可用蒸煮损失率和离心损失率来表示。目前,关于滩羊肉的研究主要集中于宰前控制对于滩羊肉品质形成的影响。宰后因素对滩羊肉品质影响方面,吴亮亮等研究发现不同食盐添加量对滩羊肉保水性、剪切力以及水分分布均有显著影响。张丽文等研究发现冻融处理的滩羊肉中不易流动水相对含量较高,自由水相对含量较低。而从蛋白质水平解析贮藏过程中滩羊肉保水性的变化机制鲜有报道。
宰后贮藏期间,肉品的保水性受贮藏条件、糖酵解、肌肉pH值等因素影响,这些因素通过调节肌肉结构蛋白质的降解影响肌肉结构从而影响肌肉结构中水分的贮存情况,而蛋白质水平的变化是引起肌肉结构变化的根本原因。左惠心研究表明糖酵解关键酶磷酸丙糖异构酶可能是牦牛肉保水性的指示蛋白之一。刘秋凤研究表明,糖代谢通过改变肌肉pH值影响猪肉保水性、肉色等品质性状的形成。因此,从蛋白质水平解析贮藏过程中保水性变化的机制,从而寻找改善汁液损失的方法,对提高冷鲜肉品质、降低企业经济损失具有重要意义。
蛋白质组学是研究细胞、组织或生物体中所表达的蛋白质组成、变化及相互作用规律的学科。蛋白质组学在肉品保水性研究相关领域也取得了可观的进展。di Luca等采用高分辨率双向荧光差异凝胶电泳(twodimensional fluorescence difference gel electrophoresis,2D-DIGE)和免疫印迹法研究猪肉渗出液,发现磷酸丙糖异构酶和转铁蛋白可能是预测保水性的生物标志物。Zuo Huixin等研究表明,肌球蛋白轻链、热休克蛋白27和磷酸丙糖异构酶可能是牦牛腰长肌保水性的指示蛋白。然而,关于揭示宰后贮藏过程中滩羊肉保水性变化的机制鲜见报道。因此,本研究以6 月龄滩羊背最长肌为研究对象,测定4 ℃贮藏0、4、8 d时样品的pH值、蒸煮损失率和离心损失率;采用基于同位素标记相对和绝对定量(isobaric tags for relative and absolute quantitation,iTRAQ)技术的蛋白质组学研究贮藏期间滩羊肌肉组织中蛋白质表达情况,筛选差异蛋白质并对其进行生物信息学分析,鉴定0~4 d和4~8 d两个贮藏阶段共有的差异表达蛋白质并构建蛋白质-蛋白质互作(protein-protein interaction,PPI)网络,利用Cytoscape 3.8.0软件对PPI进行分析并筛选核心网络;通过分析品质指标与核心网络中蛋白质表达量的关系,初步揭示宰后贮藏过程中滩羊肉保水性变化的机制,为冷鲜滩羊肉品质控制技术提供理论指导。
1 材料与方法
1.1 材料与试剂
6 月龄公滩羊采集于宁夏盐池县大夏牧场食品有限公司。
尿素、二硫苏糖醇、碘乙酰胺、IPG buffer、甲酸美国GE公司;十二烷基磺酸钠、三羧基氨基甲烷、三氯乙酸、过硫酸铵、四甲基乙二胺 美国Amresco公司;三乙二胺碳酸盐、考马斯亮蓝G-250 美国Sigma公司;胰蛋白酶 美国普洛麦格公司;乙腈 美国Thermo公司。
1.2 仪器与设备
便携式pH计 上海德图仪器国际贸易有限公司;TG16W型离心机 湖南湘仪实验室仪器开发有限公司;DZKW-C型电子恒温水浴锅 沧州新三思试验仪器有限公司;Sciex Triple TOF 5600质谱仪、Eksigent nanoLC-1D plus液相色谱系统(配有纳升喷雾III离子源、Analyst TF 1.6数据处理软件) 美国应用生物系统公司;5910R冷冻离心机 德国艾本德公司;Image Scanner III光密度扫描仪 美国GE Healthcare公司;真空冷冻干燥机 美国赛默飞公司;VCX130超声波细胞粉碎机 美国Sonics公司;Mini PROTEAN tetra Cell电泳仪 美国伯乐公司。
1.3 方法
1.3.1 样品采集
选取体质量相近的6 月龄公滩羊,屠宰后用灭菌刀取右侧背最长肌,编号后置于聚乙稀薄膜内,于风速3 m/s、相对湿度80%、温度4 ℃条件下贮存。
取贮藏0、4、8 d的样品各200 g用于测定pH值、离心损失率及蒸煮损失率;取0、4、8 d的样品各200 mg迅速置于液氮中放置2 h后转移至-80 ℃冰箱中保存,用于蛋白质组学分析。
1.3.2 蛋白质组学分析
采用尤丽琴等的方法从滩羊肉中提取总蛋白。iTRAQ-蛋白质组学分析委托上海鹿明生物科技有限公司进行,运用Analyst TS 1.1软件系统获得蛋白质组学数据。蛋白质的定性及定量信采用Protein Pilot 5.0软件获得。检索的数据库为数据库,数据库来源为UniProt。数据进行归一化处理后,以差异倍数(fold change,FC)>1.5或FC<1/1.5且<0.05为标准筛选差异丰度蛋白质。筛选出的差异丰度蛋白质利用OmicsBean软件进行生物信息学分析,比对后获得差异基因的基因本体(gene ontology,GO)(包括生物过程、分子功能、细胞组分)功能信息,选择差异最显著的前10 个条目作图。采用京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)数据库对所筛选的差异蛋白质进行通路注释分析。利用String数据库(https://www.string-db.org/)和Cytoscape 3.8.0软件进行PPI分析及核心互作网络筛选。
1.3.3 指标测定
1.3.3.1 pH值
取4 cm×4 cm×4 cm(60 g左右)滩羊背最长肌,使用便携式数字pH计测量肌肉pH值,探头沿肌纤维方向插入肉样内部,每个样品至少测定3 个点,结果取均值。测定前剔除样品可见脂肪与结缔组织。
1.3.3.2 蒸煮损失率
蒸煮损失率的测定参考文献[14]的方法并稍作修改。将样品切割成6 cm×3 cm×3 cm的肉块并称质量(),然后在80 ℃水浴锅中加热至样品中心温度75 ℃,取出后冷却至室温,用滤纸擦干表面水分后称质量(),按式(1)计算蒸煮损失率。

1.3.3.3 离心损失率
离心损失率的测定参照文献[15]的方法并稍作修改。将约5 g的样品准确称质量()后放置于10 mL离心管中,4 ℃以8 000 r/min离心10 min。除去离心管中渗出液后以滤纸吸干样品表面水分后准确称质量()。按式(2)计算离心损失率。
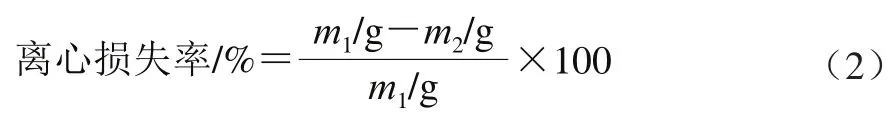
1.4 数据处理与分析
所有指标重复测定3 次,结果以平均值±标准差表示。采用Excel软件统计数据,采用SPSS软件进行单因素方差分析及Pearson相关性分析,采用Duncan检验进行显著性分析,<0.05表示差异显著,采用Origin软件作图。采用微生信在线平台(http://www.bioinformatics.com.cn)对筛选的差异蛋白质进行主成分分析。
2 结果与分析
2.1 冷藏期间滩羊背最长肌pH值及保水性变化
由表1可知,宰后0~4 d,pH值显著降低(<0.05),并于4 d时降至最低(5.58),4 d后略有回升。王薇等发现滩羊肉于0~4 ℃贮藏条件下连续放置7 d后其pH值呈先下降后上升趋势,本实验研究结果与其一致。国内外诸多学者研究表明家畜屠宰后初期ATP水解产生导致H含量增加以及能量代谢方式转变引起的乳酸积累是导致贮藏初期pH值下降的主要原因,而pH值4~8 d升高可能是由于分解代谢消耗肌肉中所积累的酸性物质。宰后贮藏过程中,肌肉pH值下降的速率和程度显著影响代谢酶的活性,对于肉色、嫩度、保水性和风味前体的形成有重要影响,是宰后初期肌肉最重要的品质指标。贮藏8 d内蒸煮损失率与离心损失率均呈持续升高趋势,二者均可表示肌肉的保水性,蒸煮损失率与离心损失率持续升高表明滩羊肉的保水性随时间的延长逐渐变差。这与李升升的研究结果一致。据研究表明,畜禽屠宰后影响肌肉保水性的主要因素有肌肉结构、pH值、糖酵解等。肌肉中有自由水、结合水和不易流动水3 种形式存在,共占肌肉的80%左右。肌肉结构、pH值及糖酵解等变化所引起的水分状态转换以及水分储存空间减少是导致滩羊肉保水性变差的主要原因。
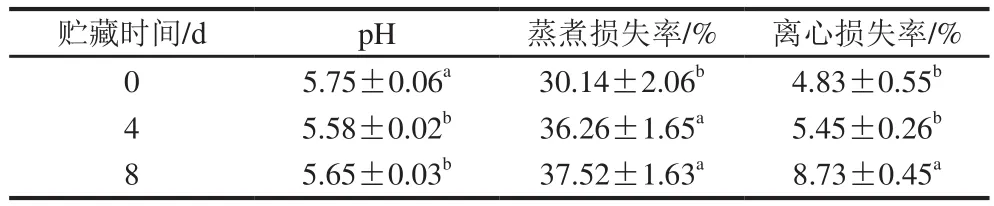
表1 滩羊宰后贮藏过程中肌肉pH值及保水性的变化Table 1 Changes of pH and water holding capacity in Tan sheep meat from different aging stages
2.2 冷藏期间滩羊背最长肌蛋白质组学数据分析结果及差异蛋白筛选
对3 个贮藏时间段样品的蛋白质组学结果进行主成分分析,如图1所示,贮藏时间可将样本很好地区分开,同一贮藏时间样本之间区分不明显,不同贮藏时间样本之间区分明显,表明本实验样品采集合理,蛋白质组学数据可靠,满足后续分析要求。

图1 不同贮藏期样本蛋白质组学数据的主成分分析结果Fig. 1 Principal component analysis plot of proteomic data of samples stored for different periods
2.2.1 差异蛋白质筛选
以FC>1.5或FC<1/1.5且<0.05为标准筛选差异丰度蛋白质,4 d组对比0 d组共鉴定出83 个丰度显著差异蛋白质,其中,61 个为显著上调,22 个为显著下调(图2A);8 d组对比4 d组共鉴定出104 个丰度显著差异蛋白质,其中,14 个为显著上调,90 个为显著下调(图2B)。
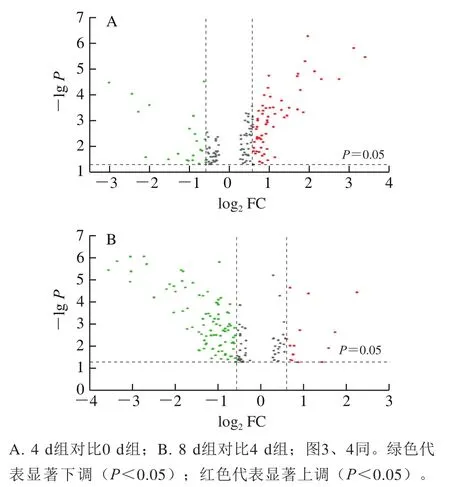
图2 不同贮藏期滩羊肉差异丰度蛋白质的火山图Fig. 2 Volcano plot of differential abundant proteins in Tan sheep meat during chilled storage
2.2.2 冷藏期间滩羊背最长肌差异蛋白质生物信息学分析结果
2.2.2.1 GO功能注释富集分析结果
通过GO功能注释对所筛选的差异丰度蛋白质进行生物信息分析,通过生物过程、细胞组分、分子功能三方面解析其功能,结果表明,4 d对比0 d组显著富集到441 个生物过程条目、117 个细胞组分条目和110 个分子功能条目;8 d对比4 d组显著富集到769 个生物过程条目、85 个细胞组分条目和180 个分子功能条目,富集最显著的前10 个条目如图3所示。

图3 不同贮藏期滩羊肉差异丰度蛋白质的GO注释Fig. 3 Gene ontology annotation of differential abundant proteins in Tan sheep meat during chilled storage
由图3A可知,宰后贮藏0~4 d,差异蛋白质主要涉及到的生物过程包括肌肉收缩、肌肉系统过程、横纹肌收缩、ATP代谢过程、骨骼肌收缩等;细胞组分主要包括肌原纤维、收缩纤维、收缩纤维部分、肌节、超分子纤维等;分子功能主要包括肌动蛋白结合、细胞骨架蛋白结合、谷胱甘肽转移酶活力、氢离子跨膜转运蛋白活力、电子载流子活度等。以上结果表明,滩羊屠宰后初期构成肌肉结构的肌原纤维、收缩纤维、肌节、超分子纤维等发生变化,这些变化可能引起了肌肉收缩、骨骼肌收缩等,发生变化的蛋白质在细胞质主要负责肌动蛋白结合、细胞骨架蛋白结合、离子转运等功能。已有诸多研究表明肌肉的收缩状态会引起肌肉结构的变化,从而引起肉品保水性的变化。
由图3B可知,宰后贮藏4~8 d,差异蛋白质主要涉及到的生物过程包括肌肉收缩、肌肉系统过程、横纹肌收缩、嘌呤核糖核苷三磷酸代谢过程、核糖核苷三磷酸代谢过程等;在细胞组分中主要包括细胞外膜结合的细胞器、胞外外泌体、细胞外囊泡、胞外细胞器、肌原纤维等;在分子功能中主要包括细胞骨架蛋白结合、过氧化物酶活力、FATZ绑定、抗氧化活性、肌动蛋白结合等。以上结果表明,滩羊肉贮藏4 d后,除了肌肉收缩等变化,差异丰度蛋白质还涉及了嘌呤核糖核苷三磷酸代谢过程、核糖核苷三磷酸代谢过程等。
2.2.2.2 KEGG通路分析结果
通过KEGG数据库对所筛选的差异蛋白质进行通路注释分析,结果表明,4 d对比0 d组显著富集到23 个KEGG通路,8 d对比4 d组显著富集到18 个KEGG通路。由图4A可知,宰后贮藏0~4 d差异蛋白质主要参与的KEGG通路包括代谢途径、碳代谢、氨基酸的生物合成、2-氧代羰基酸代谢、淀粉和蔗糖代谢等;由图4B可知,宰后贮藏4~8 d差异蛋白质主要参与的KEGG通路包括碳代谢、氨基酸的生物合成、代谢途径、2-氧代羰基酸代谢、糖酵解/糖异生等。这些代谢过程可能通过影响结构蛋白质参与的生物学过程间接影响肉品的保水性。

图4 不同贮藏期滩羊肉差异蛋白质的 KEGG通路注释Fig. 4 KEGG pathway annotation of differential abundant proteins in Tan sheep muscle during chilled storage
2.2.3 核心网络的构建
如图5所示,34 个蛋白质为0~4 d与4~8 d两个贮藏阶段共有的差异丰度蛋白质。利用String数据库(https://www.string-db.org/)对共有的差异蛋白质进行PPI分析,结果如图6所示,共有34 个节点,其中24 个差异蛋白质之间具有明显的互作关系。该网络中PPI富集的值为1.0×10,聚类系数为0.521。将PPI网络导入Cytoscape 3.8.0软件进行核心互作网络筛选,结果如图7所示,10 个蛋白质(原肌球蛋白2(tropomyosin 2,TPM2)、肌球蛋白轻链1(myosin light chain 1,MYL1)、肌钙蛋白I2(troponin I2,TNNI2)、肌钙蛋白C2(troponin C2,TNNC2)、肌钙蛋白T3(troponin T3,TNNT3)、肌球蛋白轻链2(myosin light chain 2,MYL2)、辅肌动蛋白α3(actinin alpha 3,ACTN3)、肌动蛋白(细胞质1)(actin cytoplasmic 1,ACTB)、肌动蛋白α1(actin alpha 1,ACTA1)、辅肌动蛋白α2(actinin alpha 2,ACTN2))处于该核心网络中,且全部为滩羊肌肉结构蛋白质。

图5 不同贮藏期滩羊肉差异丰度蛋白质的维恩图Fig. 5 Venn diagram showing unique and shared differential abundant proteins
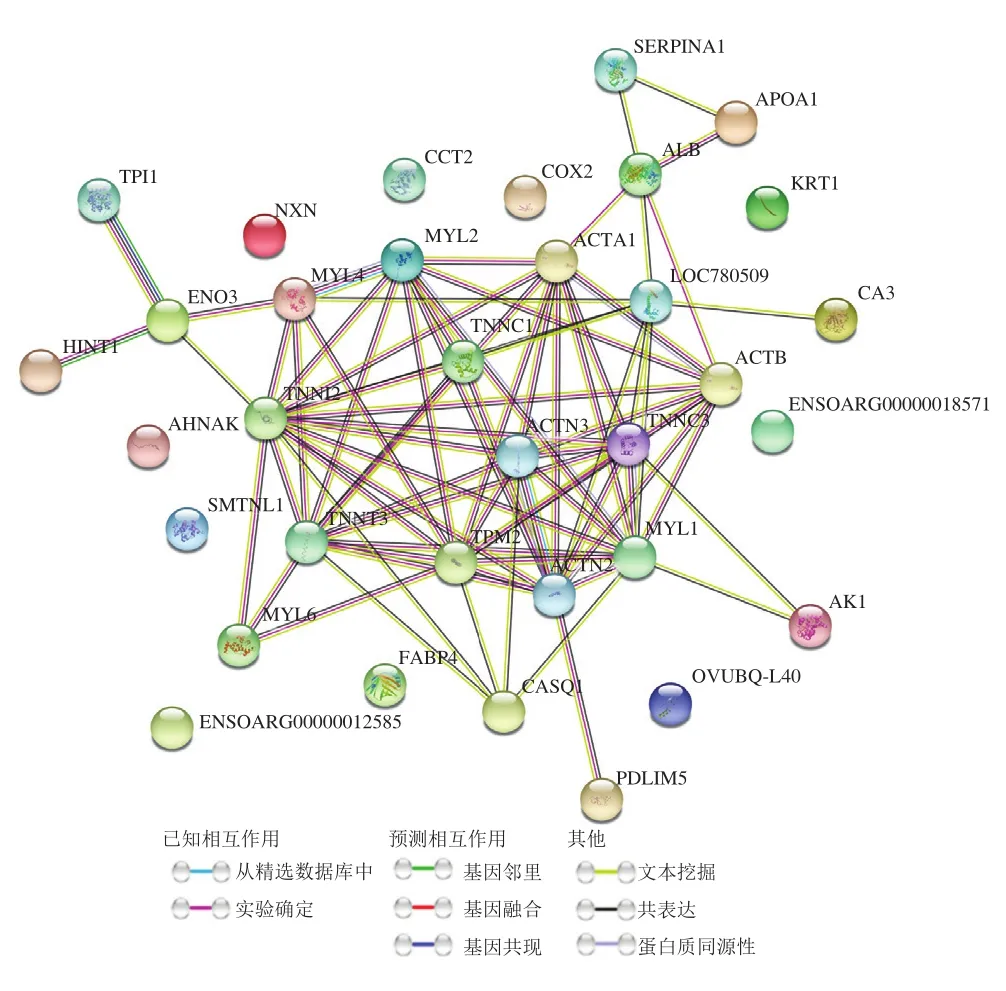
图6 共有差异丰度蛋白质的PPI网络Fig. 6 Interaction network of shared differential abundance proteins

图7 PPI核心网络的构建Fig. 7 Constructed core protein-protein interaction (PPI) network
由表2可知,TPM2、MYL1、TNNI2、TNNC2、TNNT3、MYL2、ACTN3、ACTB、ACTA1、ACTN2均为滩羊肌肉结构蛋白质,且绝大多数蛋白质的表达下调发生在4~8 d,可能是由于这些结构蛋白质于贮藏后期发生了降解,而肌肉中的水分又储存于肌肉结构中,因此对滩羊肉保水性具有重要影响。TPM2、MYL1和肌钙蛋白等在0~4 d内上调表达可能是由于应激蛋白的防护作用试图维持肌肉结构的稳定。这些蛋白质位于肌节、细胞器,与细胞骨架蛋白或离子结合,参与肌肉收缩以及ATP酶活性的调节,富集于心肌收缩和肌动蛋白细胞骨架调节的代谢通路,表明这些蛋白质通过肌肉的收缩及肌肉结构的变化来影响滩羊肉的保水性。
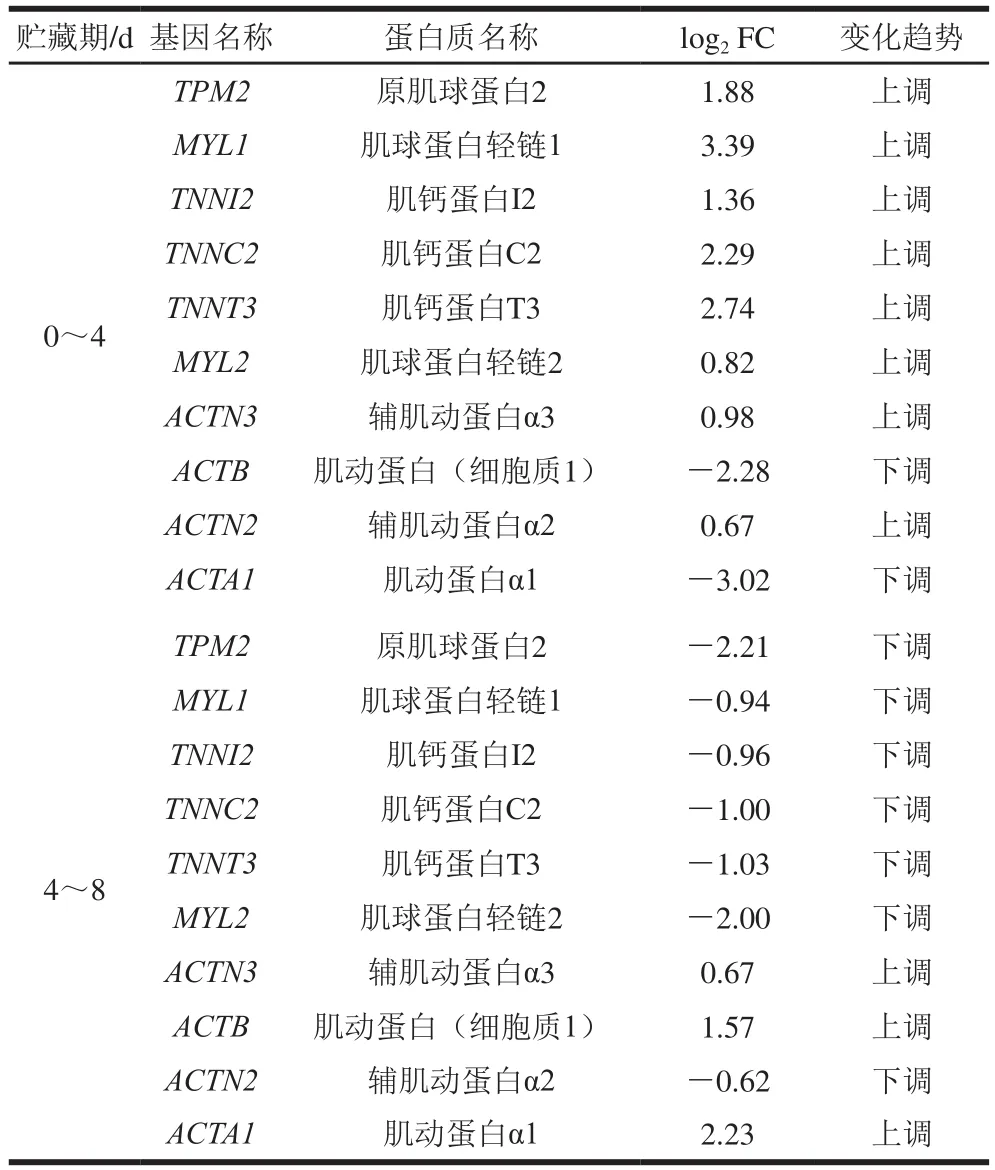
表2 核心网络中差异丰度蛋白质表达量的变化趋势Table 2 Changes in expression levels of differential abundance proteins in the core PPI network
2.3 影响滩羊肉保水性的机制
通过Pearson相关性分析确定核心蛋白质表达量与滩羊肉保水性的关系。由表3可知,蒸煮损失率与离心损失率及MYL1表达量呈显著正相关(<0.05),与肌肉pH值及ACTA1表达量呈显著负相关(<0.05),与ACTN3表达量呈极显著正相关(<0.01);离心损失率与MYL2表达量呈显著负相关(<0.05),ACTN)表达量呈极显著正相关(<0.01);pH值与ACTB、ACTA1呈极显著正相关(<0.01),与MYL1、TNNC2、TNNT3和ACTN2表达量呈极显著负相关(<0.01),与TPM2、TNNI2表达量呈显著负相关(<0.05)。相关性分析结果表明,MYL1、ACTN3、ACTA1直接影响滩羊肉蒸煮损失,MYL2和ACTN3直接影响滩羊肉离心损失,其他结构蛋白质可能通过间接方式影响贮藏过程中滩羊肉保水性的变化。

表3 滩羊肉品质指标与核心差异丰度蛋白质的相关性Table 3 Correlations between meat quality traits and core differential abundance proteins
相关性分析表明,结构蛋白质对蒸煮损失、离心损失均有显著影响,肌肉pH值影响宰后贮藏过程中TNNC2、TNNT3等结构蛋白质的变化,且肌肉pH值与滩羊肉保水性密切相关。诸多研究表明宰后贮藏期间钙蛋白酶体系和细胞凋亡酶体系等组织蛋白水解酶促使细胞骨架蛋白降解会引起肌肉结构的变化。Koohmaraie等研究表明,钙蛋白酶系统在宰后肌肉蛋白质水解中起着重要作用。其中-钙蛋白酶诱导的关键肌原纤维蛋白降解是宰后肌肉蛋白水解的原因之一。王琳琳等研究表明,通过控制宰后肌肉细胞凋亡发生进程来调节肌肉内环境变化。肌肉中的水分主要以自由水、结合水和不易流动水3 种形式储存于肌肉结构中,因此肌肉的收缩及肌肉结构的变化均可引起保水性的变化。郭兆斌等研究表明由于pH值的下降导致肌原纤维网格结构收缩,同时蒸煮条件下肌肉蛋白质的热变性使得肌原纤维紧缩,储存水分空间变小,部分不易流动水变为自由水而流失。此外,pH值降低会导致维持细胞骨架的连接蛋白变性,致使离心损失升高,且当pH值接近蛋白质等电点时,蛋白质静电荷为零,对水的吸引能力减弱,也会造成水分流失。
蒸煮损失及离心损失均可反映滩羊肉保水性的变化情况,本研究中蒸煮损失率与MYL1表达量呈显著正相关,与肌肉pH值及ACTA1表达量呈显著负相关,与ACTN3表达量呈极显著正相关;离心损失率与MYL2表达量呈显著负相关,与ACTN3表达量呈极显著正相关。肌动蛋白与肌球蛋白均为骨骼肌中主要的肌原纤维蛋白,其中肌球蛋白占骨骼肌总蛋白的1/3以上,构成肌肉结构中的粗丝,肌动蛋白构成细丝,二者在肌肉收缩中起着关键作用,产生收缩力。ACTN3作为骨骼肌Z线的构成部分,负责形成肌动蛋白-肌动蛋白交联,与快型肌纤维功能有关,而快型肌纤维功能又是引起肌肉剧烈收缩的原因,肌肉收缩会引起细胞间及细胞内储存水分的空间减小,且会导致不易流动水转化为自由水,由此引起水分流失的增加。因此,ACTA1、ACTN3、MYL2对滩羊肉的保水性有显著影响。由于ACTN3与蒸煮损失率、离心损失率均具有很高的相关性,推测其可作为滩羊肉保水性的指示蛋白。左惠心采用蛋白质组学技术研究宰后牦牛肉保水性机制的过程中发现结构蛋白质作为差异蛋白的一类,对牦牛肉保水性有重要影响,本研究结果与其一致。
pH值对ACTB、ACTA1、MYL1、TNNC2、TNNT3、ACTN2、TPM2、TNNI2均有显著影响,这些蛋白质分别为肌动蛋白(ACTB、ACTA1、ACTN2)、肌球蛋白轻链(MYL1)、肌钙蛋白(TNNC2、TNNT3、TNNI2)、原肌球蛋白(TPM2),是肌肉结构的主要构成蛋白质。其中,原肌球蛋白是一种高度保守的肌动蛋白结合蛋白,对肌动蛋白丝的稳定和许多肌动蛋白功能的协同调节至关重要;肌钙蛋白复合物由3 种调节蛋白组成,分别是肌钙蛋白C、肌钙蛋白I和肌钙蛋白T,可调节骨骼肌和心肌的肌肉收缩,TNNT3是一种仅在骨骼肌细胞中表达的快速骨骼肌肌钙蛋白;ACTB参与了细胞肌动蛋白细胞骨架的形成;ACTA1对交叉连接的肌动蛋白丝起作用,并将其稳定在相邻细胞之间、细胞与细胞外基质连接等结构上,在胞质分裂、细胞黏附和细胞迁移中发挥重要作用。pH值对这些结构蛋白质有重要影响可能是由于pH值变化直接影响了结构蛋白水解酶的活性。pH值对这些结构蛋白质的重要影响意味着屠宰后pH值变化显著影响肌肉保水性等品质指标。肖雄等研究表明,在屠宰初期,由于氧气供应中断,机体细胞很快处于无氧环境,葡萄糖及糖原的有氧分解变为无氧酵解,产生乳酸,肌肉pH值下降,从而使蛋白质分子的多肽链变紧密、分子间距缩小,肌肉中的部分水分子被挤出,因而保水性下降;且当pH值下降至蛋白质的等电点,蛋白质(尤其是肌球蛋白)所携带的净电荷为零,其正负电荷数量基本相等,蛋白质中的阳性和阴性基团互相吸引,对水的吸引能力减弱,导致水分流失。此外,肌球蛋白头部在低pH值下的部分变性也被认为是肌原纤维晶格间距收缩的主要原因,收缩可能会压迫肌细胞导致细胞内的水分流出,最终也会影响滩羊肉的保水性。
3 结 论
宰后0~8 d内,随着贮藏时间延长,滩羊肉pH值先降低后有所回升,蒸煮损失率与离心损失率持续升高。利用iTRAQ蛋白质组学技术分析鉴定发现TPM2、MYL1、TNNI2、TNNC2、TNNT3、MYL2、ACTN3、ACTB、ACTA1、ACTN2共10 个核心差异丰度蛋白质。其中TPM2、MYL1、TNNI2、TNNC2、TNNT3、MYL2、ACTN2在0~4 d表达上调,4~8 d下调;ACTB、ACTA1在0~4 d表达下调,4~8 d 上调;ACTN3 在0~8 d持续上调。这些蛋白质位于肌节、肌纤维、细胞膜,通过与细胞骨架蛋白或离子结合,参与肌肉收缩以及ATP酶活性的调节,引起肌肉收缩及肌肉结构变化,最终影响滩羊肉的保水性。肌肉pH值对肌肉结构的主要构成蛋白质ACTB、ACTA1、MYL1、TNNC2、TNNT3、ACTN2、TPM2、TNNI2有显著影响,pH值及MYL1、ACTA1、ACTN3对蒸煮损失有重要影响,MYL2与ACTN3对离心损失有重要影响。
