六堡茶渥堆过程中可培养微生物的分离与鉴定
2022-09-01黄紫衡滕建文韦保耀
黄紫衡,陈 欢,黄 丽,*,夏 宁,滕建文,韦保耀,陈 瑜,余 炼
(1.广西大学轻工与食品工程学院,广西 南宁 530000;2.贵州省兴义市敬南镇人民政府,贵州 兴义 562402)
六堡茶是中国黑茶的典型代表之一,以其原产地广西梧州六堡镇命名。六堡茶耐久贮藏,其外形色泽黑褐油润,汤色红浓明亮,滋味醇和爽口,香气醇陈,叶底铜褐色,故素以“红、浓、醇、陈”四绝而著称。六堡茶原料经杀青、初揉、堆闷、复揉、干燥工艺制成毛茶,再经过筛选、拼配、汽蒸或不汽蒸、渥堆、汽蒸、压制成型或不压制成型、陈化,最终得到成品,在生产中会根据不同产品需要而设计不同的压制工艺流程。其中,渥堆对六堡茶品质形成具有决定性的作用,渥堆过程是在高温高湿的环境下微生物大量繁殖并且分泌多种胞外酶,茶叶在这些微生物酶的酶促作用以及微生物呼吸代谢产生的热量以及水分的湿热作用下,发生了茶多酚的氧化聚合,蛋白质的分解、降解和碳水化合物的分解,还有各种产物之间的聚合缩合等一系列反应。因此微生物活动对六堡茶特殊风味和品质的形成具有重要作用,特别是在渥堆工序中微生物十分丰富和活跃。探明六堡茶在加工过程中以及成品中存在的微生物种类对研究六堡茶的品质和安全性具有重大意义。
为能客观真实地反映六堡茶微生物的群落特征,高通量测序技术采用免培养方法,可直接从样品中提取DNA进行扩增从而获得微生物信息。Zhao Ming、Ma Yan等认为曲霉属真菌来自于普洱茶原料且在整个发酵过程中为优势菌;赵仁亮等采用Illumina MiSeq技术研究报道了生产于不同地区的茯砖茶中曲霉属在每个样品中的丰度均在92%以上,是绝对优势菌种,厚壁菌门中的乳球菌属为第一优势细菌,还检测到毕赤酵母属、假丝酵母属以及耐碱酵母属,但相对丰度小于1%。Zhang Yongjie等应用高通量rDNA扩增子测序技术表明普洱茶的细菌群落结构在发酵前后变化显著,发酵后的普洱茶细菌多样性上升,真菌多样性下降;Li Qin等采用Illumina MiSeq测序技术分析茯砖茶加工过程中真菌群落的演替,结果显示在发酵早期(原料黑茶到发酵3 d)曲霉属、和念珠菌属占优势,随后,只有曲霉属仍占优势并且至发酵结束一直保持相对稳定。Mao Yan通过MiSeq测序技术得出在属水平上成品六堡茶的优势菌为曲霉属,其次是青霉属和酵母。陈庆金等采用MiSeq测序技术对陈化初期不同时段的六堡茶真菌多样性进行了分析,在属水平上真菌以曲霉属为优势菌。在属水平上,曲霉属无疑是黑茶中的优势菌属,葡萄球菌属、短状杆菌属、考克氏菌属、芽孢杆菌等是黑茶中普遍存在的优势细菌。虽然现代分子生物学技术(下一代测序技术)避免了微生物的培养,直接通过提取研究对象中微生物的总DNA分析样品中菌群的多样性信息,更能反映样品中微生物分布的真实样貌以及菌落结构的变化,但仅采用高通量测序的方法,无法获得纯种菌株,也无法对其进行功能性、产毒性等更进一步的研究。所以将传统纯培养技术和分子生物学技术两者结合起来,既能客观全面地揭示样品中微生物的多样性信息,也可以对感兴趣的菌株进行进一步的深入研究。
对于黑茶中微生物的鉴定已经有很多,而在六堡茶渥堆阶段微生物的研究较少。周梦珍等在梧州茶厂4 种六堡散茶中分离纯化得到菌核曲霉、江西青霉、阿姆斯特丹曲霉、赤曲霉、杂色曲霉变种、稻枝孢、、枝状枝孢、无花果假尾孢9 种真菌;欧惠算从12 种不同年份、不同茶厂的六堡茶茶样中分离鉴定得到8 个属20 株微生物,并表明六堡茶微生物具有多样性,主要以曲霉属、青霉属和酵母属为主;徐书泽利用传统分离培养技术在3 个不同茶厂的14 个六堡茶样品中分离鉴定得到5 个属18 个种的微生物,包括曲霉属8 种, 青霉属7 种,拟青霉属1 种,散囊菌属1 种和酵母1 种,真菌数量范围为3.9×10~1.1×l0CFU/g;温志杰等的研究结果表明,霉菌数量在六堡茶渥堆过程中最高可达1.5×10CFU/g,其研究认为六堡茶渥堆过程中细菌的数量最大,其次是酵母和霉菌。胡沛然从渥堆过程的六堡茶中分离鉴定得到嗜热菌,其中包括细菌5 个属、14 个种,霉菌5 个属、9 个种。但目前对六堡茶微生物多样性的探索还远不够,且以成品茶中的微生物为研究对象的研究较多,涉及到六堡茶渥堆生产过程的微生物尤其是细菌的研究较少。因此,本研究以渥堆前的毛茶和各个渥堆阶段的六堡茶为研究对象,从中分离出细菌、真菌和放线菌,并采用ITS和16S rDNA序列分析对其进行鉴定,旨在得到六堡茶发酵过程中微生物种类的组成及变化,从微生物的种水平上了解六堡茶渥堆工艺的微生物变化。
1 材料与方法
1.1 材料与试剂
1.1.1 材料
样品采集于广西梧州市,取渥堆前的毛茶、渥堆前期(发酵15 d)、中期(发酵45 d)、后期(发酵75 d)、末期(渥堆结束,发酵105 d)的样品,贮存于无菌容器,低温运输至实验室后立即进行分离。
1.1.2 培养基
ISP7培养基:甘油15 mL,-酪氨酸0.5 g,-天冬酰胺1 g,复合盐溶液10 mL,磷酸氢二钾0.5 g,蒸馏水1 L,琼脂1.4%,pH 7.3,每300 mL加重铬酸钾和磷酸氢二钾各1 mL;马铃薯葡萄糖琼脂培养基:称取200 g马铃薯,洗净去皮切碎,加水500 mL左右煮沸0.5 h,过滤,再加20 g葡萄糖,琼脂1.4%,加水至1 L;ISP2培养基:酵母提取粉2.0 g,麦芽提取粉2.0 g,葡萄糖2.0 g,琼脂14 g,蒸馏水1 L,pH 7.2;氯硝胺18%甘油(DG18)琼脂培养基:酪蛋白胨5 g,无水葡萄糖10 g,磷酸氢二钾1 g,硫酸镁0.5 g,氯硝胺0.002 g,无水甘油200 g,琼脂15 g,氯霉素0.1 g,蒸馏水1 L;形态学观察所用的培养基为查氏(Czapek dox agar,CA)培养基和查氏酵母膏(Czapek yeast autolysate agar,CYA)培养基。上述培养基配制后于高压蒸汽灭菌锅中121 ℃条件下灭菌20 min备用。
1.2 仪器与设备
T100Thermal Cyeler聚合酶链式反应(polymerase chain reaction,PCR)仪、Universal Hood II紫外成像仪、PowerPacBasic电泳仪 美国Bio-Rad公司;MC6000微型离心机、DH300干式恒温器 杭州瑞诚仪器有限公司。
1.3 方法
1.3.1 样品处理
每个茶叶样品取2 g于预先灭菌的研钵磨碎,加入20 mL无菌蒸馏水中混匀,用旋涡振荡器振荡30 min。接种方法为取1 mL原液依次稀释至10、10、10、10、10,用移液枪分别吸取200 μL每个稀释度的样品菌悬液,均匀涂布于已经配置好的平板培养基上,每个样品每个稀释度每种培养基涂布3 个平板,分别置于28、37、48 ℃的恒温培养箱进行培养。
1.3.2 六堡茶中微生物的分离纯化
待菌落长出后,根据菌落的生长特征进行分离纯化。挑出平板培养基上的单个菌落,用接种针将其接种于适合生长的培养基上划线培养,反复接种分离2~3 次直至得到纯的菌株,至恒温培养箱中培养,记录编号后于4 ℃保存。
1.3.3 菌株DNA的提取
取50 μL灭菌的树脂于八连管,用灭菌的牙签挑取极少部分上述分离纯化的单菌落在装有树脂的八连管中研磨,金属浴10 min后离心10 min取上清液制得模板DNA以备用。
1.3.4 PCR扩增及测序
细菌PCR扩增引物:27F(5’-AGTTTGATCMTGGCTCAG-3’)和1492R(5’-GGTTACCTTGTTACGACTT-3’)。真菌PCR扩增引物:ITS1(5’-TCCGTAGGTGAACCTGCGG-3’)和ITS4(5’-TCCTCCGCTTATTGATATGC-3’)。PCR体系(25 μL):模板DNA 0.5 μL,mix酶12.5 μL,引物A 0.5 μL,引物B 0.5 μL,无菌水11 μL。细菌PCR扩增条件:94 ℃预变性4 min,94 ℃变性45 s,55 ℃退火45 s,72 ℃延伸1 min,30 个循环,最后72 ℃延伸8 min。真菌PCR扩增条件:94 ℃预变性5 min,94 ℃变性30 s,55 ℃退火35 s,72 ℃延伸1 min,35 个循环,最后72 ℃延伸8 min。
1.3.5 BLAST比对和系统发育分析
将PCR产物送往生工生物工程(上海)股份有限公司进行序列测定,测序后的结果在NCBI上用BLAST软件序列比对,初步确定所分离菌株的种属,并将测得的序列与已登录的该属其他标准菌株进行系统发育树的构建。
2 结果与分析
2.1 六堡茶渥堆过程中的细菌种类及其作用
2.1.1 细菌和放线菌的分子学鉴定
将六堡茶中分离出的细菌和放线菌进行DNA 序列分析,分子鉴定结果显示序列相似度超过 97%,分别如表1、2所示,根据16S rDNA序列鉴定用Neighbor-Joining构建系统发育树,分离出的菌株可以确定为细菌13 个属、27 个种,放线菌7 个属、8 个种。系统发育树分别如图1、2所示。分离到的微生物,大多都是芽孢杆菌属、地芽孢杆菌属、考克氏菌属、乳杆菌属、葡萄球菌属等。
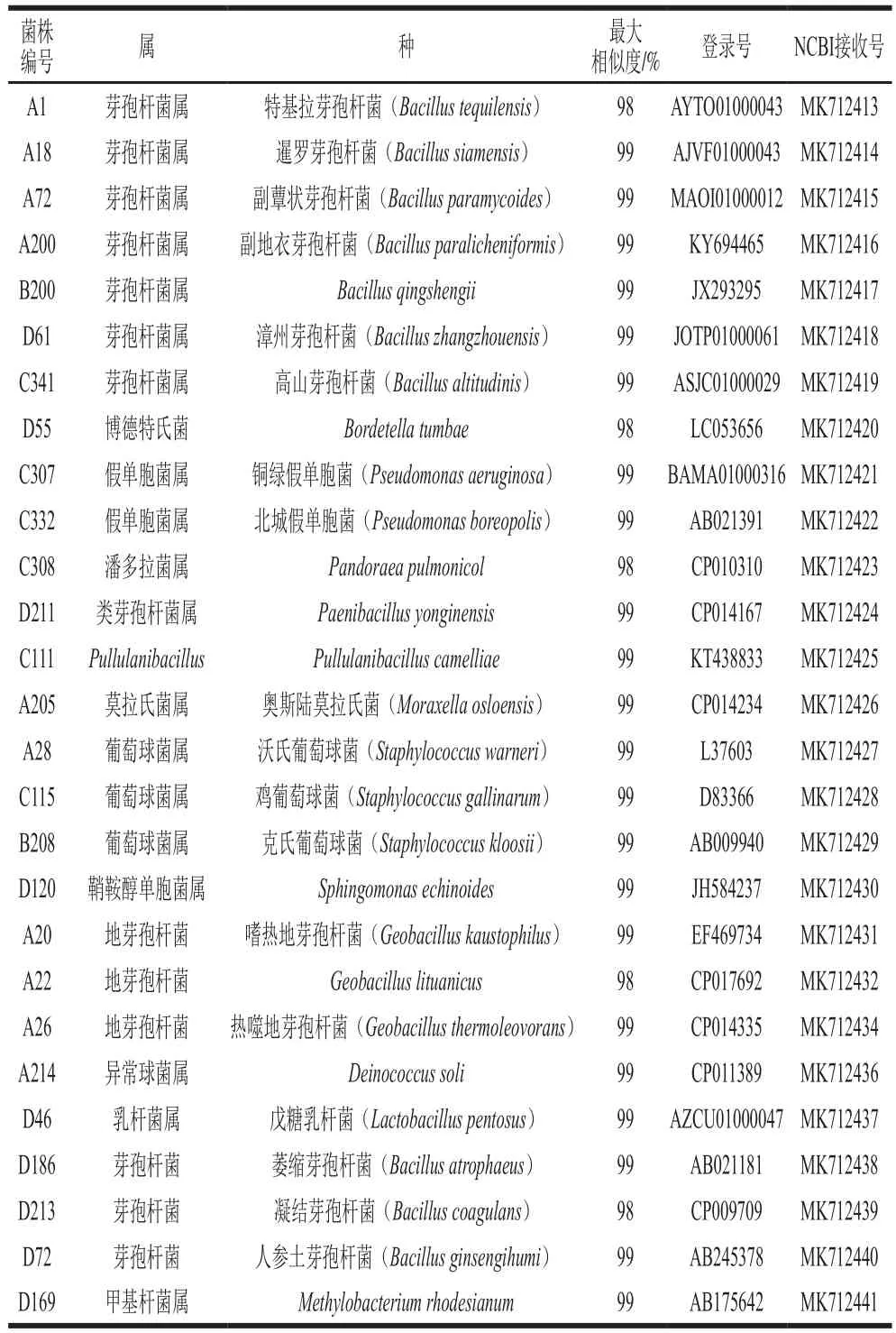
表1 细菌的16S rDNA分子鉴定Table 1 Molecular identification of bacteria from Liupao tea by 16S rDNA sequence

表2 放线菌的16S rDNA分子鉴定Table 2 Molecular identification of actinomycetes from Liupao tea by 16S rDNA sequence

图1 从六堡茶中分离的细菌与BLAST比对结果的系统发育树Fig. 1 Phylogenetic tree constructed for bacteria isolated from Liupao tea using basic local alignment search tool (BLAST) results

图2 从六堡茶中分离的放线菌与BLAST比对结果的系统发育树Fig. 2 Phylogenetic tree constructed for actinomycetes isolated from Liupao tea based on BLAST results
2.1.2 细菌和放线菌的分布情况
由表3、4可知,六堡茶渥堆过程中各阶段、各层级之间的菌落结构不尽相同。芽孢杆菌属在毛茶及渥堆阶段的样品中均存在,且分离所得数量最多;其次是地芽孢杆菌,但仅在渥堆前期茶堆内部样品且高温培养条件下得到;再次之是葡萄球菌属,在渥堆前期、后期及末期均有检出;考克氏菌属在各渥堆阶段的样品中均存在,分离所得数量较多。同时,在渥堆末期分离鉴定到4 株戊糖乳杆菌。毛茶中仅分离到高地芽孢杆菌,渥堆前期和末期的菌落结构较渥堆中期及后期复杂,渥堆前期以芽孢杆菌属、葡萄球菌属以及地芽孢杆菌属等为优势菌,渥堆末期以葡萄球菌属、考克氏菌属等为优势菌。

表3 六堡茶渥堆过程中细菌分布情况Table 3 Distribution of bacteria in the fermentation process of Liupao tea
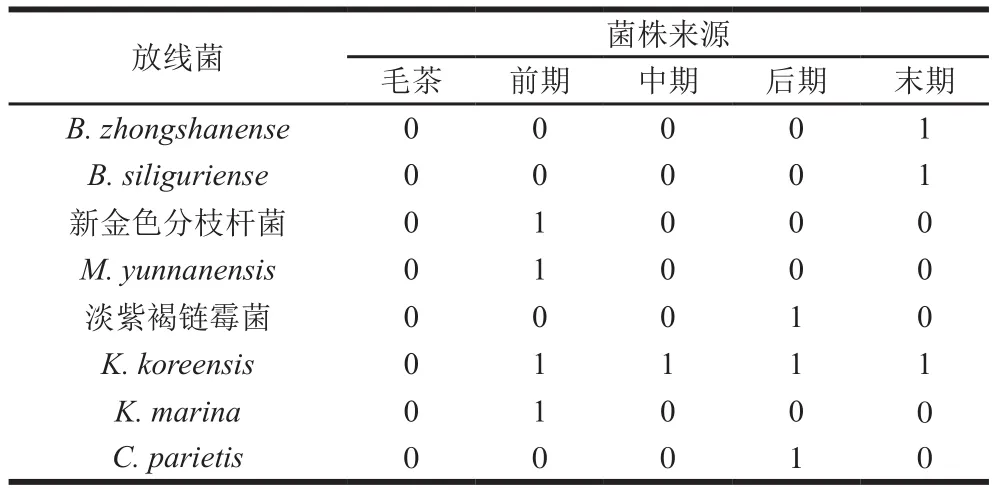
表4 六堡茶渥堆过程中放线菌的分布情况Table 4 Distribution of actinomycetes in the fermentation process of Liupao tea
2.2 六堡茶渥堆过程中的真菌种类及其作用
2.2.1 真菌的形态学观察与分子学鉴定
运用可培养方法从渥堆过程的六堡茶中分离出种类较为丰富的霉菌菌株,将分离纯化得到的菌株接种于CA和CYA培养基上,进行形态观察,部分霉菌菌落的形态特征如表5所示,参照《真菌鉴定手册》和《中国真菌志》对菌株进行观察,结合表6以及系统发育树的分子鉴定结果得到的20 种真菌分别为橘青霉、草酸青霉、、、微小根毛霉、、黑曲霉、塔宾曲霉、高渗曲霉、冠突曲霉、、聚多曲霉、杂色曲霉、烟曲霉、unknown_p_Ascomycota、、球孢枝孢、芽枝状枝孢、伞状毛霉菌、sp.,各菌株的形态特征如图3所示。

图3 菌株在CA培养基和CYA培养基上形态学特征Fig. 3 Morphological characteristics of fungal strains grown on CA medium and CYA medium

表5 菌株的形态特征及初步鉴定Table 5 Morphological characteristics and preliminary identification of fungal strains
根据表6所示的ITS序列鉴定结果,用Neighbor-Joining构建系统发育树,如图4所示。图3、4结果显示,分子鉴定结果与形态分类结果一致,序列相似度超过97%。由系统发育树可判断分离出的霉菌菌株的种属,本研究分离出的真菌可以确定为共11 个属、23 个种,具体见表6。分离到的微生物,大多都是曲霉属、、青霉属等。

表6 真菌的ITS序列分子鉴定Table 6 Molecular identification of fungi based on internal transcribed spacer (ITS) sequence

图4 从六堡茶中分离的真菌与BLAST比对结果的系统发育树Fig. 4 Phylogenetic tree constructed for fungi isolated from Liupao tea based on BLAST results
2.2.2 真菌的分布情况
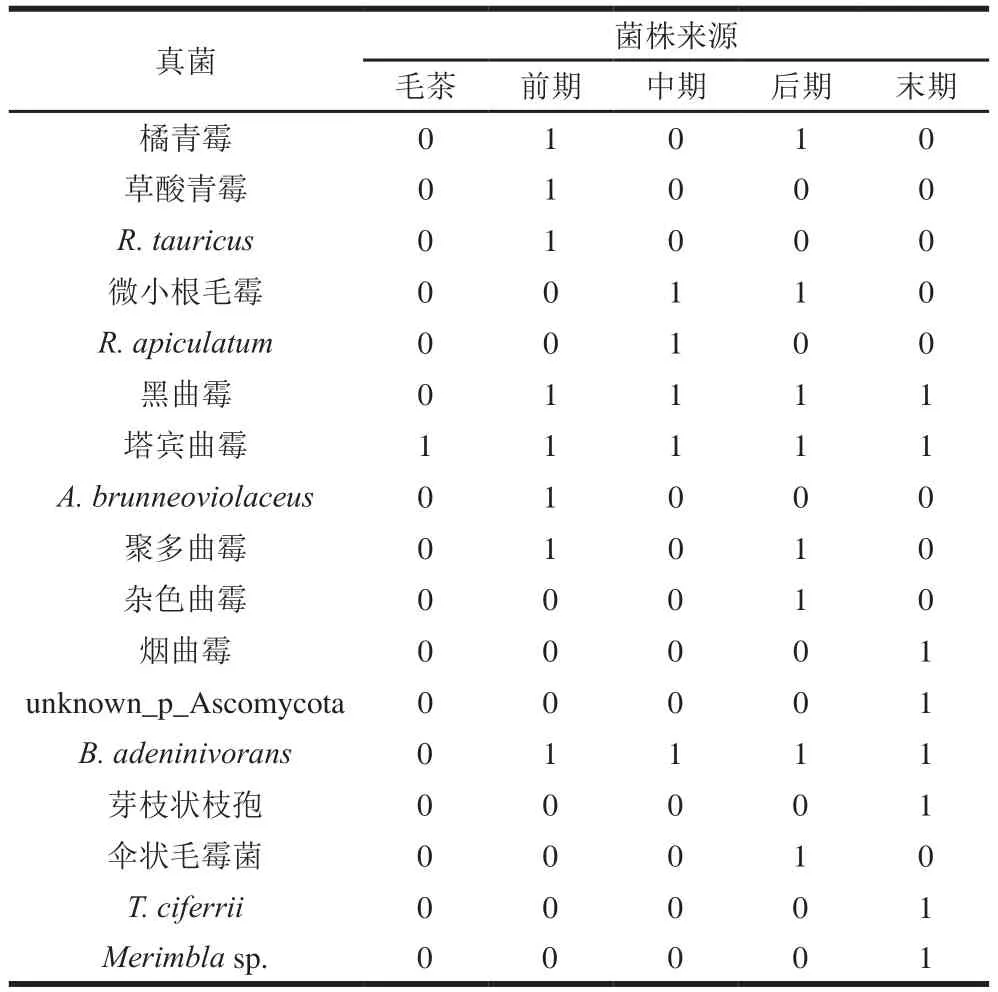
表7 六堡茶渥堆过程中真菌的分布情况Table 7 Distribution of fungi in the fermentation process of Liupao tea
在各渥堆阶段的样品中均存在,且分离所得数量最多;从毛茶到渥堆结束的样品中均分离到曲霉属,分离所得数量较大,以黑曲霉和塔宾曲霉为主;青霉属数量虽不多但在渥堆前期和后期被检测到;渥堆前、中、后期均发现根毛霉属。由表7可知,毛茶中仅分离到塔宾曲霉,渥堆开始后各阶段的菌群结构复杂程度相当,组成上既有差异也有联系,渥堆前期以酵母、曲霉、青霉、根毛霉为优势菌,酵母、曲霉在中期、后期、末期继续保持优势地位,同时在后期、末期也出现少量其他种属的真菌。本研究结果显示曲霉属是六堡茶渥堆过程中第一优势真菌,同时是第二优势真菌。
3 讨 论
通过可培养方法从渥堆过程的六堡茶中分离到细菌13 个属、27 个种,放线菌7 个属、8 个种,真菌11 个属、23 个种。以黑曲霉和塔宾曲霉为主的曲霉属和在整个渥堆过程均保持优势地位。芽孢杆菌属、葡萄球菌属以及地芽孢杆菌属等为渥堆前期的优势细菌,葡萄球菌属、考克氏菌属等为渥堆末期的优势细菌。与传统可培养方法相比,高通量测序技术能够检测到更多的微生物类群,例如六堡茶渥堆过程中已检测到鞘鞍醇单胞菌属、甲基杆菌属、、、乳杆菌属、考克氏菌属、、、短状杆菌属、短杆菌属、、葡萄球菌属、等属水平上的主要细菌,六堡茶渥堆过程中的优势真菌有曲霉属(包括黑曲霉、黄曲霉、灰绿曲霉、冠突曲霉)、青霉属、。本研究分离到的真菌的种属在属水平上与以上高通量测序结果中的优势真菌一致,通过纯培养分离到的细菌的种属与现有高通量测序结果中共同鉴定到的属有芽孢杆菌属、假单胞菌属、潘多拉菌属、葡萄球菌属、鞘鞍醇单胞菌属、乳杆菌属、短状杆菌属、短杆菌属、、链霉菌属、考克氏菌属、柠檬菌属,但发现博德特氏菌、类芽孢杆菌属、、莫拉氏菌属、地芽孢杆菌属、异常球菌属、甲基杆菌属、是高通量测序方法没有鉴定的属,因此采用两种方法相结合更有利于研究和应用。
由本研究结果可知,渥堆前期以酵母、曲霉、青霉、根毛霉为优势真菌,酵母、曲霉在中期、后期、末期继续保持优势地位。目前关于六堡茶渥堆、陈化过程和成品茶中的微生物分离鉴定结果已报道过41种真菌,本研究分离鉴定到的真菌中有9 种为首次在六堡茶中报道,即、、、高渗曲霉、、、布兰克念珠菌、球孢枝孢、sp.。众所周知曲霉属是黑茶后发酵过程中的主要菌群,数量庞大,参与茶褐素和其他优势风味物质的形成,因其具有丰富的泌酶系统而被广泛应用到工农业生产中。但其中有些菌种是重要的发酵工业微生物,有些菌种却是环境中的致病菌。例如,黑曲霉能够分泌多种水解酶,促进甘味物质和多酚类物质的转化,还对黑茶特殊的“陈香”的形成有重要贡献;塔宾曲霉在普洱茶中被用于生产高含量的茶褐素;聚多曲霉可以降解咖啡因,将其转化为茶碱,减少茶汤苦味;冠突曲霉分泌的各种酶作为高效的生化动力促进茶叶中相关物质的氧化聚合、转化和分解,使得茶汤颜色红亮,苦涩味减弱,同时,该菌的次级代谢产物具有调节胆固醇代谢、抗肿瘤等保健功能;首次在黑茶中被分离出,曾被报道从室内空气环境中分离出,不产生赭曲霉毒素A,通常具有比较好的产酸能力;米曲霉能够提高绿茶的抗氧化性,因此还用于生产发酵绿茶。青霉属被学者研究证明能够在普洱茶渥堆发酵过程中分泌纤维素酶并对其甘味物质的形成具有显著促进作用。纤维素酶作用于茶叶中的纤维素使之分解成回甘物质如单糖、双糖和寡糖。草酸青霉被证明适合于黑茶发酵,用其发酵后的茶样汤色棕红、香气甜浓、味道醇和,安全性高。枝状枝孢霉在自然界中主要存在于木本植物、土壤中等,根据已有报道此菌可高效降解纤维素,是很好的产纤维素酶酶源。球孢枝孢首次在黑茶中被分离出,具有产纤维素的能力,可以水解茶叶基质的纤维素。酵母菌在黑茶渥堆过程中数量庞大,有助于黑茶内含物质的转化,对其品质的形成具有重要作用。本实验分离到的优势酵母为(syn.),它是一种非常规、非致病性、具有耐热性和耐渗透性,属于酵母亚门,被誉为具有巨大科技潜力的二态酵母。温志杰等已证明是六堡茶渥堆发酵环节最主要的优势酵母菌,其产生的多种胞外酶在高温条件下表现出高活力,对茶叶成分转化及风味形成起着重要作用。几乎所有陈年普洱茶中都以酵母(spp.)为主,且酵母有助于改善在普洱茶发酵和陈酿过程中形成的滋味。布兰克念珠菌为耐高温酵母,在普洱茶发酵中期和后期被大量分离到,被认为是发酵过程的主要菌种之一,能够明显促进茶多酚转化为茶色素。本研究检出的微小根毛霉,在黑茶中多次被鉴定为主要的嗜热真菌。同样地,也是一种嗜热真菌,被报道为普洱茶固态发酵中的优势真菌,具有将茶多酚转化为茶褐素的能力。由渥堆的高温环境推测以上检出的嗜热微生物参与了六堡茶渥堆发酵的过程,已有研究表明高温嗜热菌能够分泌多种胞外酶,因此菌种鉴定结果为探明渥堆发酵过程中高温嗜热菌的种类和变化规律及其作用机制,都做出了积极的贡献。
另外,本研究结果显示,渥堆前期以芽孢杆菌属、葡萄球菌属及地芽孢杆菌属等为优势细菌,渥堆末期以葡萄球菌属、考克氏菌属等为优势细菌。目前关于六堡茶中的细菌分离鉴定结果仅报道过渥堆过程中的14 种嗜热菌、5 种乳酸菌和鞘氨醇单胞菌,本实验分离鉴定的细菌戊糖乳杆菌、嗜热地芽孢杆菌、副地衣芽孢杆菌、、热噬地芽孢杆菌外均为首次在六堡茶中分离。芽孢杆菌属中多数为有益菌,不仅具有耐酸、耐高温等特点,还能分泌多酚氧化酶和过氧化物酶,茶叶中的黄烷醇类及其糖苷类能够为二者提供底物,而多酚氧化酶对黑茶中茶黄素、茶红素等物质的形成起着关键作用。因此芽孢杆菌可能对六堡茶茶汤色泽以及功能活性物质的形成具有重要意义。特基拉芽孢杆菌作为益生菌能够利用碳源产生葡萄糖、半乳糖、甘露糖、阿拉伯糖和木糖等胞外多糖。从发酵的香豆分离出来,被认为参与了香豆香气化合物的形成。从植物叶组织分离出的漳州芽孢杆菌被证实会产生苯酚酸、黄酮类化合物和单宁等具有天然生物活性的酚类化合物,并对常见致病菌具有抗菌活性。高地芽孢杆菌具有产木聚糖酶的能力,将木聚糖转化为它的组成单糖。北城假单胞菌可以分泌不含纤维素酶的木聚糖酶。葡萄球菌属耐高温,兼性厌氧,付润华等用传统培养方法得到四川康砖茶中主要细菌也是芽孢杆菌属和葡萄球菌属。考克氏菌属能发酵产生木聚糖酶,玫瑰考克氏菌()被证明具有较强的蛋白质水解能力,可能与茶叶中氨基酸含量的变化有关,进而影响茶汤滋味。短杆菌属高产胆固醇氧化酶,而胆固醇氧化酶是胆固醇代谢过程中的关键酶之一,因此,推测六堡茶醇提物调节高血脂作用与其渥堆过程中含有的大量短杆菌属有关。甲基杆菌属被认为是黑茶渥堆过程中核心功能微生物之一,可能对六堡茶的品质形成具有一定作用。戊糖乳杆菌具有较强的抑制致病菌能力,同时产酸耐酸较强,此外乳酸菌还能分泌多种胞外酶,对六堡茶渥堆过程中微生物生长环境如pH值的变化以及六堡茶风味品质具有一定作用。姚静等从普洱茶中也检出了欧文氏菌属、考克氏菌属、假单胞杆菌属、克雷白氏菌属,但是其对普洱茶渥堆发酵的作用需要进一步研究。
目前关于六堡茶加工过程中放线菌的报道较少,除外,本研究分离到的放线菌也是第一次在六堡茶中报道。放线菌具有很强的生物合成能力,能够合成具有广泛生物活性的次级代谢产物。能产生多种糖苷酶,最适生长温度为28 ℃,是一种寡营养细菌。淡紫褐链霉菌是一种产葡萄糖异构酶(glucose isomerase,GI)的放线菌,而GI作为胞内酶能够参与进入体内的木糖的应用,作为胞外酶能够将-葡萄糖转化为-果糖,被大量应用于高果糖浆的工业生产中。本研究通过ITS和16S rDNA序列分析发现了在以往黑茶和六堡茶研究中尚鲜见报道的一定数量的新种。由鉴定结果可以从微生物学角度说明六堡茶是有效利用微生物作用的优良发酵食品,深入研究和挖掘这些新种在微生物分类学和发酵六堡茶微生物资源的保护方面具有积极意义。
