放牧强度对半干旱草地优势植物赖草功能性状的影响
2022-08-31郭琛文姚蒙蒙董宽虎王常慧任国华
张 琦, 郭琛文, 姚蒙蒙, 董宽虎, 王常慧, 任国华*
(1. 山西农业大学草业学院, 山西 太谷 030801; 2. 草地生态保护与乡土草种质创新山西省重点实验室, 山西 太谷 030801;3. 山西右玉黄土高原草地生态系统定位观测研究站, 山西 右玉 037200; 4. 农业农村部饲草高效生产模式创新重点实验室,山西 太谷 030801)
放牧是草地生态系统中最为普遍的干扰方式[1],研究放牧对草地植物的影响对于维持草地结构和功能的稳定以及制定科学合理的草地管理政策具有重要意义。放牧干扰对植物生长会产生一定的影响,植物本身也会通过调节其外部形态及生理特征产生避牧或耐牧机制以适应环境变化[2]。一方面,放牧引起草地植物植株变矮、叶片变短变窄、节间缩短、冠(丛)幅变小、地上生物量减小和根系分布浅层化等[3-4],迫使植物形态结构矮化以躲避放牧干扰;另一方面,长期放牧干扰下,草地由高抗逆性的耐牧植物主导,其具有高的比叶面积、叶氮含量和光合能力等[5],通过增强获取资源的能力和提高生长速率适应家畜的采食。
目前,对于植物功能性状的研究,人们已经经历了从植物性状单一水平到性状间资源权衡和从单一器官种类到植株整体以及各器官(根、茎、叶、种子等)间资源权衡的两方面的发展[6-7],从功能性状变化来探索植物对放牧的适应性和生态对策受到广泛关注,探索植物生理生态学适应机制已成为近100年来放牧研究的主要内容[8]。植物功能性状对表征生态系统功能具有指示作用[9],对放牧干扰来说也是很好的判断指标。有研究表明,比叶面积可作为荒漠草原植物在放牧干扰下资源利用对策的指示指标[10];植株高度能够反映植物在不同放牧干扰梯度下获取资源的能力[11];叶片的高度和位置体现了植物性状对草食动物采食的抵御能力[12];叶氮含量和叶干物质含量等体现了植物性状在放牧干扰后的恢复能力[13]。但也有研究发现,单一性状并不能完整表征和指示植被的变化[14],放牧管理中也应当考虑植物性状间的权衡。李西良等[3]指出放牧下植物性状的权衡可表现在性状之间的正相关与异速变化的关系上,如在草甸草原的放牧研究中,羊草性状可塑性程度与其变异性之间符合指数增长的拟合关系[15]。Damhoureyeh和Hartnett[16]研究发现,放牧会影响牧草的繁殖策略,使种子变小、花期提前,减少了有性繁殖分配,体现植物营养繁殖策略的权衡。近年来,对植物各功能器官的研究已取得重大进展,人们陆续发现了叶经济型谱(Leaf economic spectrum,LES)[6]、木材经济型谱(Wood economic spectrum,WES)[17]和根经济型谱(Root economic spectrum,RES)[18]等功能性状协变和权衡的生态策略,这些虽是在大尺度的基础上建立的,但仍具有广泛参考意义。植物叶片的光合作用是植物获取营养物质的主要来源[19],有研究指出,放牧对植株表型特征的影响,往往通过叶片经济性状的权衡来实现[20]。植物响应放牧的干扰,不只体现在叶性状的变化上,也包括植株和根系等植物性状的差异化响应上。大量研究表明,随牧压强度的不同,植物会通过地上、地下等不同功能器官资源分配模式和繁殖分配策略的变化达到表型性状的改变,最终使自己朝着有利于提高种群适合度的方向发展[8]。因此,无论是植物的各个性状还是器官之间,研究其在环境胁迫下的资源权衡方式是功能生态学研究的重要内容,对探索植物生理生态适应能力甚至生态系统功能稳定性具有重要意义。
晋北农牧交错带是北方农牧交错带的一部分,是典型的干旱、半干旱地区[21],在气候和地貌的双重作用以及农业和牧业的双重影响下,生态环境极其脆弱。赖草(Leymussecalinus)为禾本科小麦族赖草属植物,属于中旱生根茎型牧草,其根茎繁衍迅速,抗寒抗旱,耐盐碱,对资源贫瘠的半干旱草地有较强的适应能力[22]。许多研究表明,稳定的自然群落是性状趋同和趋异构建模式的动态平衡,群落中优势种的功能性状对表征生境变化和群落演替方向具有一定的指示作用[23-24]。优势种可以更好地代表生态系统或者植物群落,其个体属性也可以更好地放大到群落水平[8]。赖草(L.secalinus)作为晋北农牧交错带半干旱草地的主要优势种[25],放牧干扰引起的赖草功能性状的改变可能会影响该草地生态系统的结构和功能。因此,本研究以半干旱草地优势植物赖草为研究对象,从多角度研究不同放牧强度对赖草功能性状的影响、各功能性状的变异程度以及各功能性状之间的关联,旨在阐明:(1)不同放牧强度对赖草各功能性状产生了怎样的影响?(2)随着放牧强度的增加,赖草采取的资源权衡策略是怎样的(包括各功能性状之间的和不同器官间的资源分配)?这些科学问题的解决,将为半干旱草地生态系统合理的放牧政策和植物在放牧干扰胁迫下的生理生态学响应机制提供科学理论支撑。
1 材料与方法
1.1 研究区概况
本实验在山西右玉黄土高原草地生态系统定位观测研究站的长期放牧平台进行,地理坐标为39°59′ N,112°19′ E。海拔1 348 m,属温带季风性气候,年均温4.7℃,年均降雨量435 mm,年蒸发量约1 000~1 500 mm,无霜期100~120 d[26]。植被类型主要为低地草甸类盐化低地草甸亚类,以赖草为主要优势种[25,27]。
1.2 样地设置
本实验区于2016年5月建立,2017年开始放牧,集中在每年的6—9月。实验采用完全随机区组设计,设4个放牧强度,依次为不放牧(Ungrazing,UG,0羊单位·hm-2·生长季-1)、轻度放牧(Light grazing,LG,2.35羊单位·hm-2·生长季-1)、中度放牧(Moderate grazing,MG,4.80羊单位·hm-2·生长季-1)、重度放牧(Heavy grazing,HG,7.85羊单位·hm-2·生长季-1)。每个处理4个重复,共计16个小区,每个小区面积为2 000 m2,用网状围栏隔开。放牧羊是体重为25~29 kg的杜泊羊,健康状况大致相同[26,28]。
1.3 功能性状测定
2020年8月,本实验以优势种赖草(L.secalinus)为实验对象,选择生长成熟、长势一致、无病害状况的赖草植株,将功能性状分为形态结构性状、生产力性状、光合生理性状和化学计量4类进行测定。其中形态结构性状包括株高(Plant height,PH)、茎长(Stem length,SL)、叶长(Leaf length,LL)、叶面积(Leaf area,LA)、比叶面积(Specific leaf area,SLA)和种子体积(Seed volume,SV);生产力性状包括整株干重(地上部分)(Whole plant dry weight (aboveground),DW)、茎叶比(Ratio of stem to leaf,S/L)和种子千粒重(Weight of 1000-seed,Sm);光合生理性状包括净光合速率(Net photosynthetic rate,Pn)、气孔导度(Stomatal conductance,Gs)和蒸腾速率(Transpiration rate,Tr);化学计量包括叶氮磷比(Leaf N/P ratio,LN/LP)和茎氮磷比(Stem N/P ratio,SN/SP),共14个性状。
形态结构和生产力性状的测定:每个小区随机选择20株赖草,采用卷尺测量株高后,齐地剪取。茎叶分离后,使用50 cm钢尺测量茎长;为了实验的标准化,均选择植株旗叶下面的第一片叶子,即倒二叶进行单片叶子叶性状指标的测定。扫描仪和图像分析软件Adobe Photoshop CC 14.2测量叶长和叶面积;将采集的样品于105℃杀青5 min,85℃烘干48 h后测得整株干重(地上部分)、单叶干重(Single leaf dry weight,SLDW)、全叶干重(Leaf dry weight,LDW)和茎干重(Stem dry weight,SDW);比叶面积为叶面积与其单叶干重之比;茎叶比为茎干重与全叶干重之比;种子体积与种子千粒重的测定参考龚时慧等[29]。
光合生理性状的测定:使用美国LI-6400XT便携式光合仪测定,设置红蓝光源为饱和光强1 800 μmol·m-2·s-1,选择晴朗无风的天气,于每天的上午9:00—下午15:00进行赖草光合参数的测量。每个小区测10组,每组3个重复。
化学计量的测定:在每个小区中,随机性的齐地剪取长势一致、无病虫害的赖草植株,茎叶分离后,于105℃杀青5 min,85℃烘干48 h。赖草茎叶全氮含量的测定采用凯氏定氮法,参考标准LY/T 1269—1999[30];茎叶全磷含量的测定采用钼锑抗比色法,参考标准LY/T 1270—1999[31]。叶氮磷比为叶氮含量(Leaf nitrogen content,LN)与叶磷含量(Leaf phosphorus content,LP)之比,茎氮磷比为茎氮含量(Stem nitrogen content,SN)与茎磷含量(Stem phosphorus content,SP)之比。
1.4 数据分析
采用Excel 2010进行数据整理,SPSS 23.0进行赖草各功能性状的单因素方差、皮尔逊相关性和主成分分析。作图均由Origin 2019b完成。利用变异系数(CV=标准差/平均值×100%)表征不同放牧强度下各功能性状的变异程度[15],其中变异系数大于50.00%属于强变异,小于等于20.00%属于弱变异,在20.00%~50.00%之间属于中等变异[32],且将变异系数在20.00%以上作为判断敏感性状的参考值;利用可塑性指数(PI=|UG-G|/UG,UG表示不放牧处理下的平均值,G表示轻、中、重度放牧处理下的平均值)表示各功能性状的敏感程度,结合变异系数和可塑性指数得出敏感性状和惰性性状。重度放牧无种子,种子指标只进行单因素方差分析和显著性检验(α=0.05)。
2 结果与分析
2.1 放牧强度对赖草各功能性状的影响
除种子千粒重外,赖草地上部各部分的形态结构性状和生产力性状在不同放牧强度之间均存在显著性差异(P<0.05)。赖草形态结构性状中的种子体积随着放牧强度的增加显著降低(图1a,P<0.05),表现为UG>LG>MG,LG和MG处理分别比UG处理下降了0.10%和0.22%;株高、茎长、叶长和叶面积随放牧强度的增加呈现出先降低后升高再降低的趋势(图2a~d,P<0.05),均表现为UG>MG>LG>HG,MG处理表现出补偿性生长能力,与UG处理的值最接近,株高、茎长、叶长和叶面积分别降低了27.66%,31.62%,25.05%,40.03%;比叶面积随放牧强度的增加呈现出先升高后降低再升高的趋势(图2e,P<0.05),表现为HG>LG>MG>UG,比叶面积在HG处理下比UG处理升高了79.88%。赖草生产力性状种子千粒重在MG处理下显著低于UG和LG处理(图1b,P<0.05),依次降低了25.81%和30.64%;整株干重(地上部分)和茎叶比随放牧强度的增加呈现出先降低后升高再降低的趋势(图2f~g,P<0.05),均表现为UG>MG>LG>HG,可知,MG处理下整株干重(地上部分)和茎叶比也均表现出补偿性生长能力,分别比UG处理降低了49.61%和8.17%。除气孔导度外,赖草光合生理性状和化学计量比在不同放牧强度之间均存在差异不显著的结果。赖草光合生理性状中,MG与UG和LG之间净光合速率无显著差异,HG处理显著高于UG,LG和MG处理(图2 h,P<0.05);气孔导度在各处理间差异显著(图2i,P<0.05),表现为HG>LG>UG>MG;蒸腾速率在UG和LG处理间差异不显著(图2j),但与气孔导度的变化趋势相同。化学计量中叶氮磷比在HG处理下显著低于UG,LG和MG处理(图2k,P<0.05),依次降低了21.13%,30.18%和30.21%;茎氮磷比在各处理间差异不显著(图2l,P>0.05),但与叶氮磷比具有相同的变化趋势。

图1 不同放牧强度对赖草种子体积和种子千粒重的影响Fig.1 Effects of different grazing intensities on seed volume and weight of 1000-seed of Leymus secalinus注:不同小写字母表示不同处理之间差异显著(P<0.05);UG,不放牧;LG,轻度放牧;MG,中度放牧;下同Note:Different lowercase letters meant significant difference at 0.05 level under different treatments. UG,Ungrazing;LG,Light grazing;MG,Moderate grazing;the same as below

图2 不同放牧强度对赖草整体和茎叶各功能性状的影响Fig.2 Effects of different grazing intensities on the functional traits from the whole,stem and leaf of Leymus secalinus注:HG,重度放牧Note:HG,Heavy grazing
2.2 赖草各功能性状的变异性分析
随着放牧强度的增加,赖草各功能性状的变异程度不同(表1)。形态结构性状、生产力性状和光合生理性状的变异范围大,变异系数的范围分别为8.67%~49.11%,21.82%~52.92%和5.49%~43.39%;化学计量的变异范围小,为11.00%~29.95%。比较赖草各个功能性状的变异系数可知,随着放牧强度的增加,变异程度达到20.00%以上的功能性状有整株干重(地上部分)(52.92%)、茎长(49.11%)、茎叶比(45.70%)、气孔导度(43.39%)等11个性状,仅净光合速率的变异系数低于20.00%。HG条件下,形态结构和生产力性状的变异系数均高于其他放牧强度。MG条件下,形态结构和生产力性状的变异系数均低于LG和HG,光合生理性状均高于LG和HG。

表1 不同放牧强度下赖草各功能性状的变异系数Table 1 Variation coefficients of each functional trait of Leymus secalinus under different grazing intensities 单位:%
参考张璐等[33]和邢小青等[34]对草地植物功能性状敏感性的研究,在不同放牧强度之间均无显著性差异的性状中(P>0.05),以其中最大可塑性指数(33.27%)作为判断敏感性状的参考值,即随着放牧强度的增加,各功能性状指标的最大可塑性指数大于33.27%时,其可塑性强,敏感度大。由表2可知,可塑性强的性状包括整株干重(地上部分)、茎长和株高等8个性状;可塑性弱的性状包括茎氮磷比、净光合速率、蒸腾速率和叶氮磷比。除蒸腾速率和叶氮磷比外,随着放牧强度的增加,各功能性状的可塑性指数变化一致,均表现为HG>LG>MG。

表2 不同放牧强度下赖草各功能性状的可塑性指数Table 2 Plasticity indexes of each functional trait of Leymus secalinus under different grazing intensities 单位:%
由此,结合CV和PI的值,敏感性状有:形态结构性状株高、茎长、叶长、叶面积和比叶面积;生产力性状整株干重(地上部分)和茎叶比;光合生理性状气孔导度。惰性性状有:光合生理性状净光合速率和蒸腾速率;化学计量比叶氮磷比和茎氮磷比。
2.3 赖草各功能性状的相关性分析
根据相关性分析结果(表3),形态结构、生产力和光合生理性状之间的相关性多表现为极显著相关(P<0.01),仅茎叶比与蒸腾速率之间关系不显著;化学计量与形态结构、生产力和光合生理性状之间的相关性较弱,其中叶氮磷比与气孔导度之间和茎氮磷比与蒸腾速率之间为显著负相关关系(P<0.05),叶氮磷比与叶面积、叶氮磷比与整株干重(地上部分)以及茎氮磷比与气孔导度之间关系不显著。比叶面积与株高等形态结构性状、茎叶比等生产力性状和叶氮磷比等化学计量之间表现为极显著负相关(P<0.01),与净光合速率等光合生理性状极显著正相关(P<0.01)。除比叶面积外,形态结构与生产力性状之间均表现为极显著正相关关系(P<0.01);除叶面积和整株干重(地上部分)外,化学计量中叶氮磷比与株高等其他形态结构性状和茎叶比之间均为极显著正相关关系(P<0.01);茎氮磷比与形态结构和生产力性状之间均为极显著正相关关系(P<0.01)。形态结构性状(比叶面积除外)、生产力性状、光合生理性状和化学计量4类功能性状各类性状内部指标之间的相关性均为极显著正相关(P<0.01)。

表3 赖草各类功能性状之间的相关性系数Table 3 Correlation coefficients among each functional trait of Leymus secalinus
2.4 赖草各功能性状的主成分分析
以特征值大于1为原则,对不同放牧强度下赖草各功能性状进行主成分分析的结果表明(表4),可提取3个主成分,反映了原始数据的绝大部分信息(86.39%),其中PC1,PC2,PC3分别可以解释总方差的58.44%,15.77%,12.18%。根据综合得分排名结果,总体上,茎氮磷比、茎长、茎叶比、叶氮磷比、株高、叶长、叶面积和整株干重(地上部分)在各放牧强度下所占权重大,对不同放牧强度下赖草各功能性状发生变化的贡献较大。对不同放牧强度下的置信区间进行分析,UG处理与LG处理相切,与MG处理相交,与HG处理相离且分别在第一主成分的正向和负向轴上。MG处理下所占主要成分的功能性状更接近于UG处理,HG处理与UG处理下具有的主要成分类别不相同:UG处理下,形态结构和生产力性状具有主要贡献;HG处理下,比叶面积和光合生理性状占据主要成分的贡献(图3)。
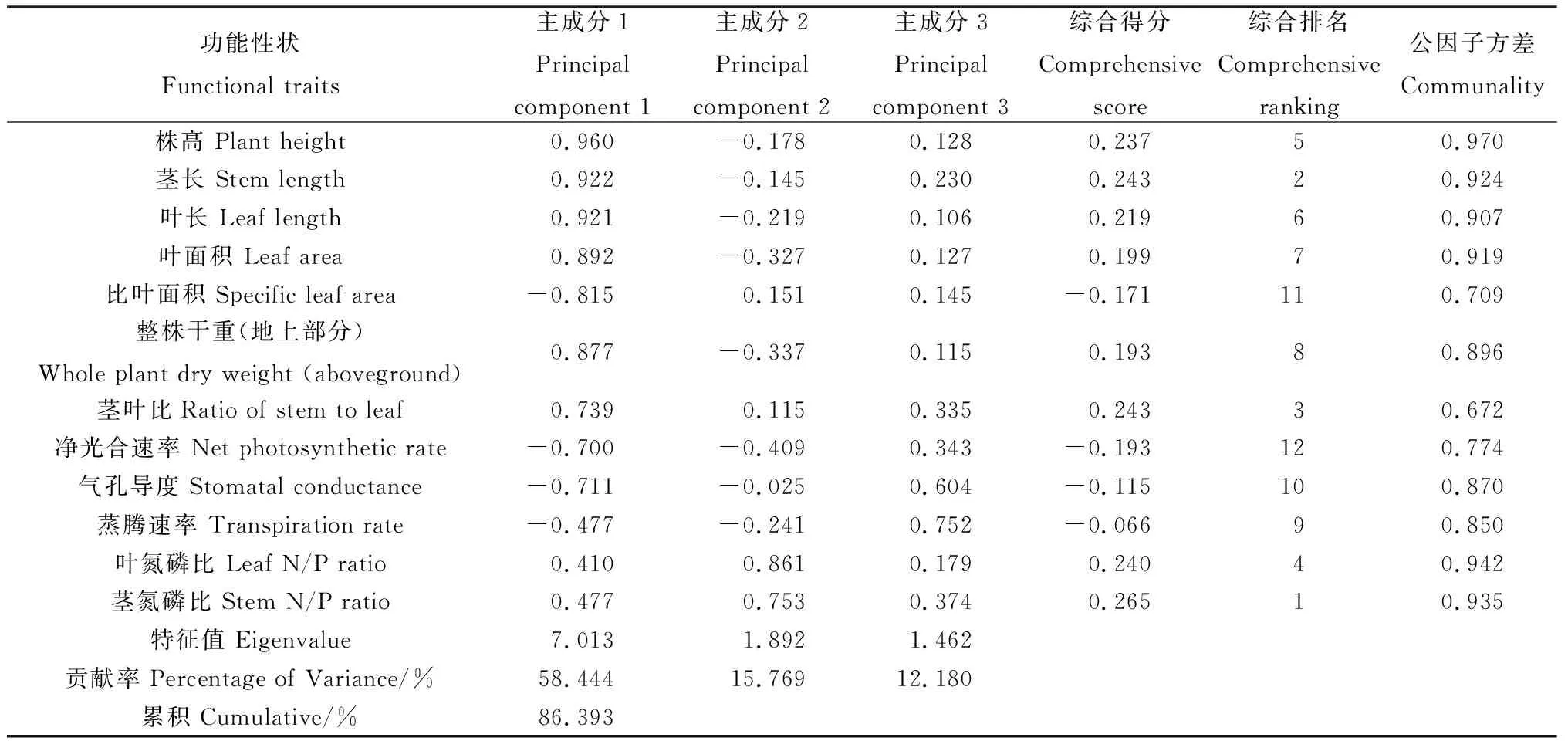
表4 赖草各功能性状初始因子载荷矩阵及主成分贡献率Table 4 Initial factor loading matrix and principal component contribution rate of each functional trait in Leymus secalinus
根据主成分分析结果,PC1和PC2能够解释赖草功能性状变异总方差的74.21%,如图3所示,与第1主成分相关的指标有茎叶比、茎长、株高、叶长、叶面积、比叶面积、整株干重(地上部分)和气孔导度;与第2主成分相关的指标有茎氮磷比和叶氮磷比。其中,比叶面积与茎叶比、茎长、株高、叶长、叶面积和整株干重(地上部分)负相关,与净光合速率、气孔导度和蒸腾速率正相关;茎氮磷比和叶氮磷比与净光合速率和蒸腾速率之间的关系均表现为负相关,与其他性状之间的关系接近于正交,相关性弱。

图3 不同放牧强度下赖草各功能性状的主成分分析Fig.3 Principal component analysis of functional traits of Leymus secalinus under different grazing intensities
3 讨论
3.1 放牧强度对赖草各功能性状的影响
不合理的放牧对草地植物的破坏性大,造成草地退化现象的发生[35]。适度放牧有利于牧草的补偿生长,能够保持一定的牧草生物量,维持较高的草地生产力[36]。本研究中,MG处理下,形态结构和生产力性状具有一定的补偿效应(图1,图2),且MG处理与UG处理表现为具有相同的主成分置信区间(图3),说明适度的采食和践踏以及粪便营养对草地的返还能够维持并提高赖草的生长能力,证明了“放牧优化假说”[37]对半干旱草地优势植物赖草的适用性。前人研究表明,长期放牧压力下,草地植物一方面整体呈现矮小化,表现出集群矮化特征:株高降低,茎叶变短,节间缩短,丛幅变小等[3,15];另一方面其生物量随牧压增强具有下降趋势[36]。本研究中,随着放牧强度的增加,赖草形态结构性状和生产力性状变化明显(P<0.05),放牧造成这两类性状的值的下降(比叶面积除外)。可知,矮小化以及生产力降低是草地植物在放牧干扰下所形成的一种普遍性的负反馈作用机制[3]。
植物对外界环境干扰的生理生态适应包括外在形态和内在生理两方面,比叶面积作为形态结构指标,表征植物叶片获取光合资源的能力,联系了光合等内在生理性状[38]。前人研究表明,较高的比叶面积是植物为了快速生长采取的一种生存策略[39-40]。本实验中,放牧增大比叶面积,赖草获取光合资源的能力增加,以此提高其耐牧性。因草食动物采食与践踏,放牧干扰造成草地植物光合作用、蒸腾作用、呼吸作用发生改变,营养物质再分配进而再生长[41]。植物的光合作用是植物生长和繁殖的重要过程,也是植物获取有机物的主要来源[19]。米雪[42]等研究表明,净光合速率、气孔导度和蒸腾速率随放牧强度的增加而增大,羊草光合“午休”现象的主要原因是非气孔因素。杨浩和罗亚晨[43]研究表明,自然降水下,高氮添加显著刺激了光合速率和蒸腾速率(P<0.05)。本实验中,MG与UG和LG之间净光合速率无显著差异(P>0.05);气孔导度在各处理间具有显著性差异(P<0.05),蒸腾速率与气孔导度的变化趋势相同,表明较低强度的放牧干扰(UG,LG和MG)下,其净光合速率与气孔闭合程度无直接联系,可能与赖草体内N素含量以及生理代谢方式和能力有关。N,P元素是限制陆地生态系统植物生长的主要营养元素[44-45],其化学计量特征是生态化学计量学研究的重要内容[46-47]。随着放牧强度的增加,叶氮含量依次为UG(14.40±0.55 g·kg-1),LG(20.68±1.88 g·kg-1),MG(18.26±1.69 g·kg-1),HG(32.10±3.80 g·kg-1),叶磷含量依次为UG(1.13±0.11 g·kg-1),LG(1.40±0.07 g·kg-1),MG(1.24±0.13 g·kg-1),HG(3.08±0.16 g·kg-1),茎氮含量依次为UG(7.26±0.45 g·kg-1),LG(10.22±0.53 g·kg-1),MG(9.64±0.85 g·kg-1),HG(18.79±2.80 g·kg-1),茎磷含量依次为UG(0.99±0.10 g·kg-1),LG(1.34±0.18 g·kg-1),MG(1.28±0.18 g·kg-1),HG(3.60±0.35 g·kg-1),它们在UG,LG和MG处理之间无显著差异(P>0.05),在HG处理下均显著高于UG,LG和MG处理(P<0.05),植物具有维持自己生命活动稳定的生理机制,这表明LG和MG处理下赖草茎叶对N,P营养元素的吸收和利用仍维持在较稳定的生理状态,但HG处理下赖草增强了对N,P元素的吸收。Koerselman和Meuleman[47]研究指出,植物氮磷比<14意味着植物对N的养分吸收受到限制,氮磷比>16则表示植物受到P养分限制,若14<氮磷比<16,植物则可能同时受N,P元素的限制或者均不受限。本研究中,叶氮磷比依次为UG(13.17±1.40),LG(14.88±1.43),MG(14.89±0.82),HG(10.39±0.90),茎氮磷比均小于14,但随着放牧强度的增加,其变化趋势与叶氮磷比相同,由此,结合茎叶的氮磷含量在不同放牧强度下的变化发现,第一,赖草茎在不同放牧强度下均受N素限制,叶片在UG和HG处理下的生长受N素限制,在LG和MG处理下均不受N,P元素的限制;第二,赖草N,P元素在茎叶中存在一定的分配比例,放牧干扰对其营养分配无影响。总体上,赖草形态结构对放牧的响应敏感,但内在生理方面,较低强度的放牧干扰(UG,LG和MG)下基本无显著差异(P>0.05),表明其干扰程度不能够影响赖草的碳同化能力以及N,P元素含量的分配;而HG处理下的光合性状显著高于UG,LG和MG处理,叶氮磷比显著低于其他3种处理,本文研究认为重度放牧干扰可能破坏了赖草的生理平衡机制,为弥补地上部分形态结构的破坏和营养的过度流失,赖草可能通过增强光合作用以及地下根系等获取有机物等营养物质的方式增加对N,P的吸收。
3.2 放牧对赖草各器官构件的影响
植物叶片性状能够客观反映植物对生长环境的适应性[48],在不同放牧强度的干扰下,表现出多种功能性状的适应性变化。茎器官与植物固碳能力、养分水分传输和防御能力等密切相关,对植物生长发育状况具有指示作用[49]。种子的数量和质量是植物繁殖能力的体现。从赖草植株各器官构件出发,在形态结构、生产力、化学计量方面,放牧对赖草植株整体、茎、叶的影响趋势相同。综合赖草的变异性(表1,表2)和主成分分析(表4),茎长在敏感性以及在放牧影响的主要贡献上均高于叶长,且茎氮磷比高于叶氮磷比,可知放牧引起的茎器官的变异程度大于叶功能性状,这同张璐等[33]和李江文等[50]对草地植物茎叶的研究结果有相似之处,在放牧干扰这一生境下,赖草偏向于叶器官的形态构建和营养供给,这可能是赖草在放牧干扰下的器官上的资源权衡分配策略,以抵抗放牧干扰。
此外,放牧引起植物繁殖策略的变化,有研究表明,放牧干扰下,植物具有有性生殖和无性生殖间的权衡[16]。本研究中,随放牧强度的增大,赖草种子有性生殖能力下降,种子质量降低,体积减小,与前人研究结果相同[2,16],且在HG处理下未采集到赖草种子,这进一步表明随着放牧强度的增加,赖草具有趋向于无性生殖的一致性。
3.3 赖草各功能性状的资源权衡策略
长期放牧条件下,草地植物为适应持续的干扰因素不断地权衡资源的分配,进而在外在形态和内在生理方面表现了特殊的植物功能性状的变化,植物性状间的权衡是植物对放牧的响应策略的重要部分[3,51]。汪诗平等[52]研究表明,随着放牧强度的增加,植物往往先采取高度和生物量降低的适应策略。本研究中,与UG相比,其他3种放牧处理下各形态结构(比叶面积除外)和生产力性状均显著降低(P<0.05),赖草对放牧干扰表现出形态矮小化和生产力降低的响应方式,矮化型变和生产力降低可能是赖草为适应长期放牧干扰所形成的避牧政策。Wright等[6]研究提出的新型叶经济谱理论指出,经济谱的一端是资源保守型的植物,另一端是资源快速获取型的植物。Cruz等[53]研究认为,长期放牧处理下的草地植物表现为低的叶组织密度,高的比叶面积、高的光合效率进行快速生长,以弥补损失的组织,增强地上部的生长能力,采取放牧干扰下的“快速资源获得型”策略,从而使植物具有更强的耐牧性。本研究中,在HG处理下,随着赖草形态结构的矮化,比叶面积和净光合速率、气孔导度、蒸腾速率等光合生理性状均显著高于其他放牧强度,可知,赖草通过高的比叶面积、高的光合效率进行快速生长,采取“快速资源获得型”策略获得生存能力,具有较强的耐牧性。植物通过一系列性状组合的调整和平衡以适应特定生境[54],各性状间的协同和变异是植物资源权衡最主要的方式。李西良等[15]通过对羊草性状的研究认为羊草为适应长期放牧干扰,各性状间的协同和变异形成的性状非对称响应是其重要的生理机制。本文中,放牧影响着赖草性状的协同变化,随着株高降低,茎长叶长缩短,叶面积减小,赖草整株干重(地上部分)下降,各功能性状之间大多表现为极显著相关(P<0.01),而变异性分析得出形态结构和生产力性状均为敏感性状,净光合速率、蒸腾速率、叶氮磷比和茎氮磷比为惰性性状,可以看出,赖草各指标在协同变化的同时,可塑性变小的程度不同,这是放牧干扰下的赖草各功能性状非对称响应的重要体现。
根据主成分分析综合得分排名结果(表4),茎氮磷比、茎长、茎叶比、叶氮磷比、株高、叶长、叶面积和整株干重(地上部分)是放牧干扰下半干旱草地赖草功能性状响应变化的主要原因,可作为判断半干旱草地赖草合理放牧强度的重要指标。茎叶化学计量比与形态结构和生产力性状之间的相关性弱,主成分分析结果中(图3),它们之间的关系接近于正交,表明赖草化学性状与这两种性状属于不同的生态策略维度。因此,不同性状类别在赖草适应放牧干扰过程中所扮演的角色不同,而对于各类功能性状的具体生物学功能和生态学意义还需进一步研究确定。
4 结论
本文探讨了半干旱草地优势植物赖草在不同放牧强度下整体(地上部)、茎、叶和种子在形态结构、生产力、光合生理和化学计量性状方面的响应差异。随着放牧强度的增加,赖草发生了单从外在形态的适应到外在形态与内在生理双重响应的变化,在中度放牧下具有补偿性生长的能力,“放牧优化假说”对半干旱草地具有适用性。赖草偏向于叶器官的形态构建和营养供给,降低了有性生殖能力。在长期放牧干扰下,茎氮磷比、茎长、茎叶比、叶氮磷比、株高、叶长、叶面积和整株干重(地上部分)是判断半干旱草地赖草合理放牧强度的重要依据。矮化型变特征和非对称性响应机制是赖草在放牧干扰下的主要适应方式,以达到避牧和耐牧作用。
