高寒草甸冷季牧场不同休牧期对土壤种子库的影响
2022-08-31马凯凯徐长林魏孔涛白梅梅刘媛媛鱼小军
马凯凯, 徐长林, 李 颖, 魏孔涛, 白梅梅, 刘媛媛, 鱼小军
(甘肃农业大学草业学院, 草业生态系统教育部重点实验室, 中-美草地畜牧业可持续发展研究中心, 甘肃 兰州 730070)
高寒草甸约占青藏高原草地面积的49%[1],是该地区典型的草地类型,在生物多样性保护、维持区域生态系统稳定和促进畜牧业发展等方面发挥了重要作用。近年来,受全球气候变暖和人类活动等影响,青藏高原高寒草甸发生严重退化,这不但影响了畜牧业生产和牧民生计,而且威胁着国家的生态安全[2-4]。有研究表明,过度放牧是引起高寒草甸退化的主要原因之一,且青藏高原海拔高、气候寒冷导致枯草期长,过度放牧造成的草地退化主要发生在冷季牧场[5-6]。因此,高寒草甸冷季牧场的科学管理和合理利用显得尤为重要。
休牧因操作简单、成本低而成为治理退化草地的重要措施之一[7]。返青期是青藏高原高寒草甸的忌牧期之一,因为刚返青的幼苗被家畜采食和践踏后光合面积减少,进而严重制约牧草后期的生长发育,且返青期牧草青黄不接,加之放牧家畜体况差,会造成家畜的“春乏”现象,返青期休牧植物群落高度、盖度及生物量等均明显提高[5,8-9]。而已有研究表明,在高寒草甸冷季牧场早于牧草返青期的土壤解冻临界期开始休牧有利于矮生嵩草(Kobresiahumilis)、线叶嵩草(Kobresiacapillifolia)和垂穗披碱草(Elymusnutans)的繁殖及线叶嵩草和垂穗披碱草的根系生长发育[7,10];可提高秦艽(Gentianamacrophylla)根系的营养物质含量和各类酶活性[11],说明关于青藏高原高寒草甸的适宜休牧期还需进一步探究。
土壤种子库是指存在于土壤表层和土壤中全部存活的种子总和[12],其直接来源是地上植被的种子雨,能直接影响地上植物群落结构和物种多样性[13]。土壤种子库作为潜在的植物种群[14],是植被经历干扰后恢复的储备资源,在植被更新、演替和恢复过程中发挥着重要作用[15]。关于土壤种子库的研究,一直是植物种群生态学的研究热点。目前,大多数学者主要对土壤种子库的物种组成、密度、时空分布格局、动态、与地上植被的关系、影响因素以及作用功能进行研究[16]。研究土壤种子库是对物种多样性研究的一个重要补充,有助于了解植被更新及演替动态,对退化植被的重建与恢复具有重要意义[17]。关于放牧管理模式对土壤种子库的研究主要集中在不同放牧强度、放牧时间、放牧干扰方式以及封育等对土壤种子库的影响。大多研究结果表明,放牧在一定程度上会降低土壤种子库的密度并改变其物种组成[18-21],但春季不同休牧期对土壤种子库的影响尚不清楚。因此,本试验以东祁连山天祝高寒草甸为研究对象,通过研究天祝高寒草甸冷季牧场不同休牧期对土壤种子库的影响,明晰青藏高原高寒草甸冷季牧场的适宜休牧期,继而制定科学的休牧管理体系,为退化高寒草甸的恢复和未退化高寒草甸的合理利用提供参考。
1 材料与方法
1.1 试验地概况
试验地设在青藏高原东北缘的天祝高寒草甸,位于甘肃农业大学高山草原试验站附近的金强河河谷(37°40′N,102°32′E),海拔2 960 m。该地年均温-0.1℃,>0℃积温1 380℃,年降水量416 mm,年蒸发量达到1 592 mm,无绝对无霜期,仅分冷热两季[7]。研究区优势种为线叶嵩草,建群种为珠芽蓼(Polygonumviviparum);伴生种有矮生嵩草、藏异燕麦(Helictotrichontibeticum)、球花蒿(Artemisiasmithii)、阴山扁蓿豆(Medicagoruthenicavar.inschanica)等。主要放牧家畜为牦牛(Bosgrunniens)和藏羊(Ovisaries)。
1.2 试验设计
休牧试验于2018年开始,根据当地土壤解冻深度和牧草返青情况设置5个休牧期:(1)土壤解冻临界期(土壤表层开始解冻)—牧草枯黄期(RP1);(2)土壤解冻后期(土层解冻深度>10 cm)—牧草枯黄期(RP2);(3)牧草返青初期(牧草返青达到30%)—牧草枯黄期(RP3);(4)牧草返青后期(牧草返青达到70%~80%)—牧草枯黄期(RP4);(5)以当地传统休牧(优势牧草高5 cm—牧草枯黄期)(RP5)为对照,共设5块样地。其中,土壤解冻深度于3月1日至牧草开始返青每日下午14:00用自制的细钢条测定[22],牧草返青数采用针刺法测定。
观测数据显示3月18日土壤表层开始解冻,4月1日土壤解冻深度达到10 cm,4月15日牧草开始返青,5月1日牧草返青达70%,5月20日优势牧草高度达5 cm,各样地依次对应的休牧起始日分别为3月19日、4月2日、4月16日、5月2日和5月21日。冷季牧场一般从10月开始放牧,但在试验过程中,考虑到10—翌年2月牧草枯黄、土壤冻结,放牧对草地的影响较小,因此各样地统一从翌年3月1日开始放牧。每个样地在放牧期间选择健康、毛色一致、体重为(180±20) kg的牦牛和(45±5) kg的藏羊进行混合放牧,按照当地牧民的习惯时间出牧和归牧,出牧前和归牧后自由饮水。每个样地的面积根据合理载畜量的草地面积计算方法计算,计算公式如下:
式中:A为样地面积(m2);I为放牧家畜的日食量(牦牛:5.8 kg干草,藏羊:1.7 kg干草[23]);B为放牧家畜的日补饲量(根据实地调查:牦牛和藏羊的日补饲量分别约为1.23 kg和0.22 kg燕麦干草);D为放牧天数;T为放牧家畜头数(表1);Y为样地牧草产量(样方刈割约为2 895 kg·ha-1);U为草地牧草利用率(80%)[10]。
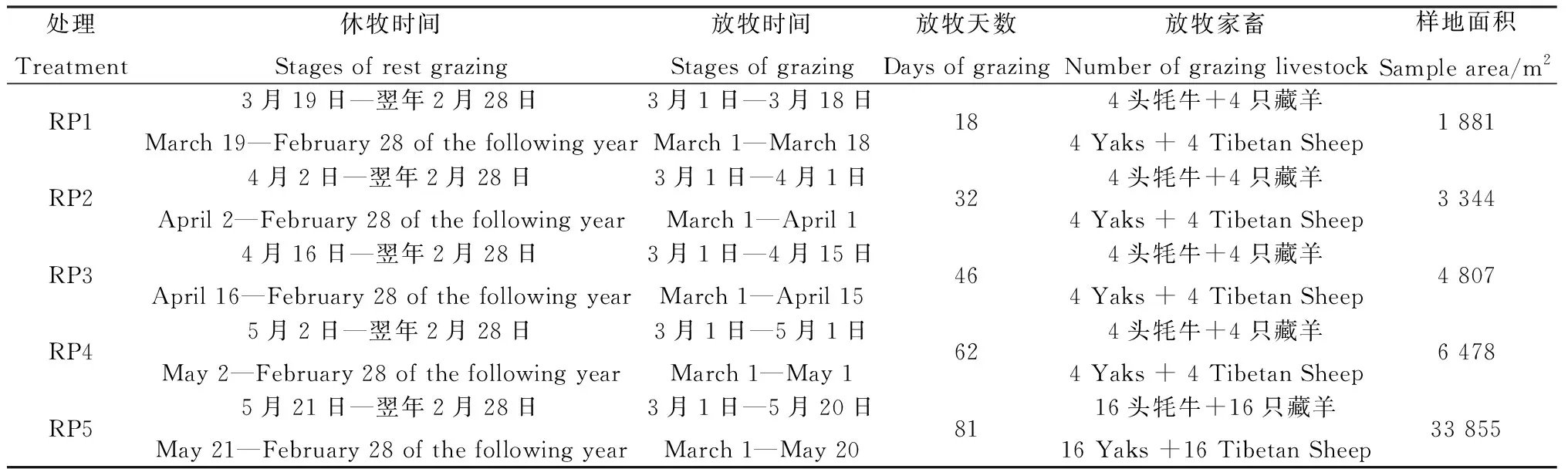
表1 休牧时间、放牧时间及样地面积Table 1 Stages of rest grazing,Stages of grazing and sample area
1.3 测定指标与方法
1.3.1土壤种子库的取样与萌发试验 于2021年5月21日,在5个休牧样地中使用自制土壤种子库取样器“S”型取样,取样面积为10 cm×20 cm,深度5 cm,每个样地重复15次,将土壤种子库样品分别装袋编号带回实验室进行萌发试验。
在萌发试验前先去除土样中较大的石砾和植物根系,将土样均匀地置入装有5 cm细沙垫底的萌发盒(24 cm×19 cm)中,土样厚度2~3 cm,垫底细沙预先在120℃的干燥恒温箱中烘24 h以消灭细沙中潜在的种子并防止污染。然后将萌发盒置于光照良好的实验室进行萌发试验,每日洒水保持土壤湿润。待幼苗长至形态特征较为明显时,及时识别鉴定并记录出现的物种及其数量,鉴定后将幼苗连根拔除,不能识别的继续培养至可以鉴定。8月中旬,将萌发盒中的土样翻动,使下层的土样置于上层进行二次萌发,直至连续2周内无种子萌发即结束试验。该土壤种子库萌发试验时间为2021年5—10月。
1.3.2地上植被调查 2021年7月下旬(植物生长旺盛期),每个样地选取5个50 cm×50 cm的样方,详细记录样方内的物种组成、物种数和盖度等。
1.4 数据处理与分析
采用Margalef丰富度指数、香农-威纳指数、辛普森多样性指数和Pielou均匀度指数[24]来描述5个休牧期土壤种子库的多样性特征。计算公式如下:
式中:S为物种总数;N为全部物种的个体总数;Pi为种i的个体在全部个体中的比例。
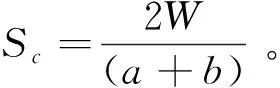
式中:Sc为Sorensen相似性系数;W为土壤种子库与地上植被共有物种数;a和b分别为土壤种子库与地上植被各自的物种数。
用Excel 2016整理计算数据及图表绘制,通过SPSS 19.0对数据进行单因素方差分析(One-Way ANOVA)和显著性分析(P=0.05)。
2 结果与分析
2.1 不同休牧期土壤种子库物种组成和密度
由表2可知,5个休牧期土壤种子库中共萌发27种植物,分属12科23属,其中禾本科植物出现比例最高,菊科植物次之,分别占22.22%和14.81%,垂穗披碱草、草地早熟禾(Poapratensis)、紫羊茅(Festucarubra)、狼毒(Stellerachamaejasme)、线叶嵩草和珠芽蓼在5个休牧期土壤种子库中均有出现。物种密度较大的植物为珠芽蓼和线叶嵩草,在RP1,RP2,RP3,RP4和RP5休牧土壤种子库中所占比例分别为51.17%,52.27%,49.89%,51.91%,49.81%和31.20%,37.98%,35.09%,34.43%,31.27%。
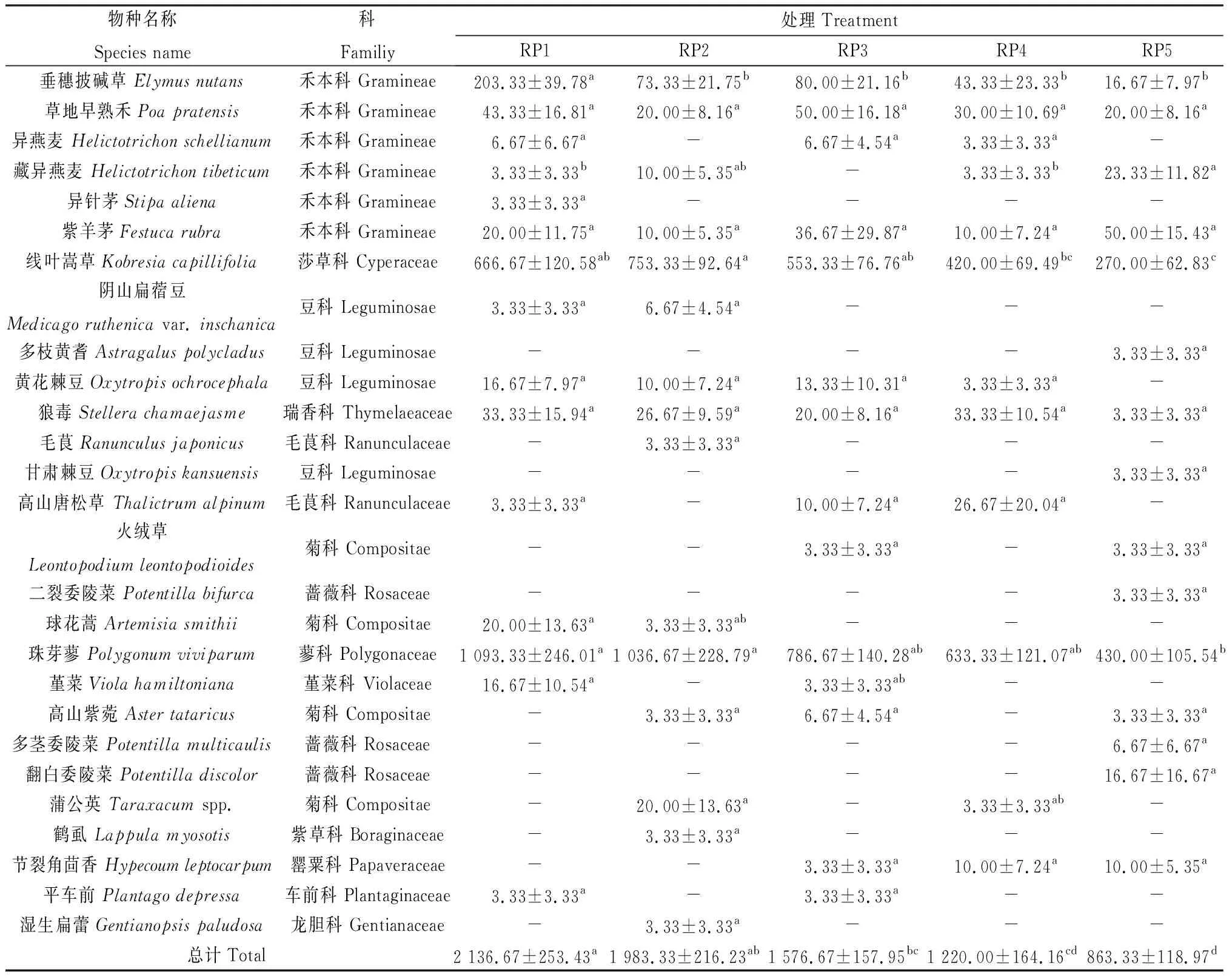
表2 不同休牧期土壤种子库物种组成和密度Table 2 Species composition and density of soil seed bank in different rest grazing periods 单位:seeds·m-2
不同处理间,RP1,RP2和RP5休牧土壤种子库中出现植物种类最多,达15种,分别隶属于9科14属、9科15属和8科13属,种子密度分别达2 136.67,1 983.33和863.33粒·m-2;RP3休牧土壤种子库中出现植物种类次之,达14种,隶属于10科14属,种子密度达1 576.67粒·m-2;而RP4休牧土壤中出现植物种类最少,为12种,隶属8科11属,种子密度达1 220粒·m-2。RP1休牧土壤种子库密度显著高于RP3,RP4和RP5(P<0.05)。
2.2 不同休牧期土壤种子库物种生活型特征
5个休牧期土壤种子库主要由地下芽植物构成,地面芽植物相对较少,一年生植物很少(表3)。地面芽植物和地下芽植物在5个休牧期土壤种子库中均有出现,其中RP1休牧的土壤种子库中地下芽植物种子密度显著高于RP4和RP5(P<0.05),一年生植物只在RP1和RP2休牧的土壤种子库中存在,但差异不显著。
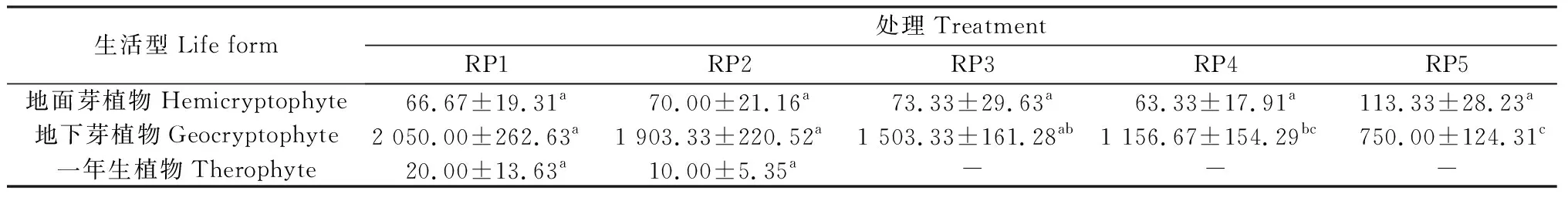
表3 不同休牧期土壤种子库生活型组成Table 3 Life form composition of soil seed bank in different rest grazing periods 单位:seeds·m-2
2.3 不同休牧期土壤种子库各功能群特征
由表4可以看出,5个休牧期土壤种子库中各功能群种子密度最高的均是杂类草,其次是莎草科植物、禾本科植物、有毒植物和豆科植物。禾本科植物、莎草科植物、杂类草和有毒植物种子在5个休牧期中均有出现,其中RP1休牧禾本科植物种子密度显著高于其他休牧期,RP2休牧莎草科植物种子密度显著高于RP4和RP5(P< 0.05),杂类草和有毒植物种子密度均在RP5休牧中最低,豆科植物只在RP1,RP2和RP5休牧中存在,但差异不显著。
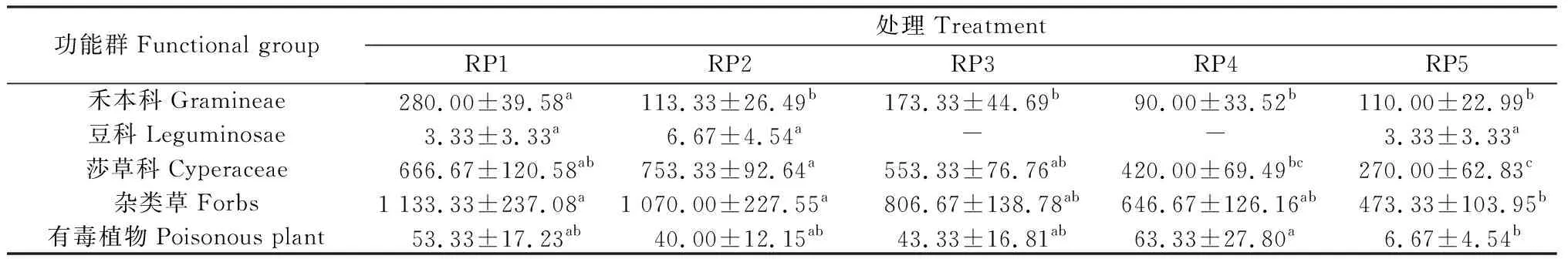
表4 不同休牧期土壤种子库各功能群种子密度Table 4 Functional groups seed density of soil seed bank in different rest grazing periods 单位:seeds·m-2
2.4 不同休牧期土壤种子库多样性特征
由图1可知,5个休牧期土壤种子库的Margalef丰富度指数在0.94~1.18之间,香农-威纳指数在0.88~1.08之间,辛普森多样性指数在0.48~0.56之间,Pielou均匀度指数在0.62~0.77之间,其中丰富度指数、香农-威纳指数和多样性指数差异均不显著,但均在RP4休牧土壤种子库中最低,香农-威纳指数和多样性指数均在RP1休牧土壤种子库中最高,丰富度指数和均匀度指数均在RP5休牧土壤种子库中最高,RP5休牧土壤种子库的均匀度指数显著高于RP2休牧(P< 0.05),5个休牧期土壤种子库的香农-威纳指数和多样性指数均表现为RP1>RP5>RP3>RP2>RP4。
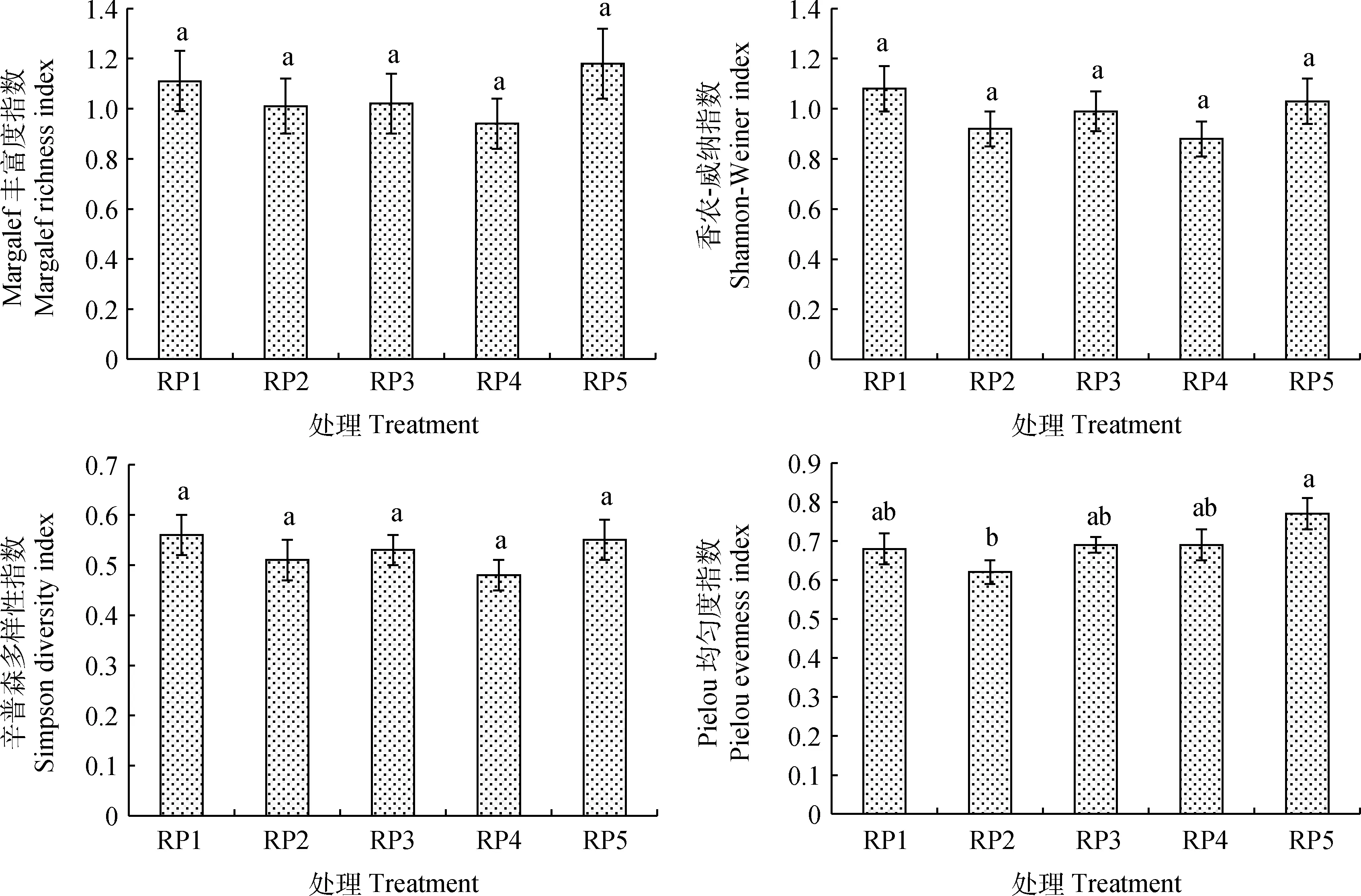
图1 土壤种子库物种多样性特征Fig.1 Species diversity characteristics of soil seed bank
2.5 不同休牧期土壤种子库与地上植被相似性
由表5可知,5个休牧期土壤种子库与地上植被的相似性系数为0.38~0.49。RP2,RP3和RP5休牧土壤种子库与地上植被相似性较低,相似性系数均为0.38,其中RP3休牧种子库与地上植被共有物种仅有9种,为处理间最低。RP4休牧土壤种子库与地上植被的相似性较高,相似性系数为0.41。RP1休牧土壤种子库与地上植被的相似性最高,相似性系数为0.49。
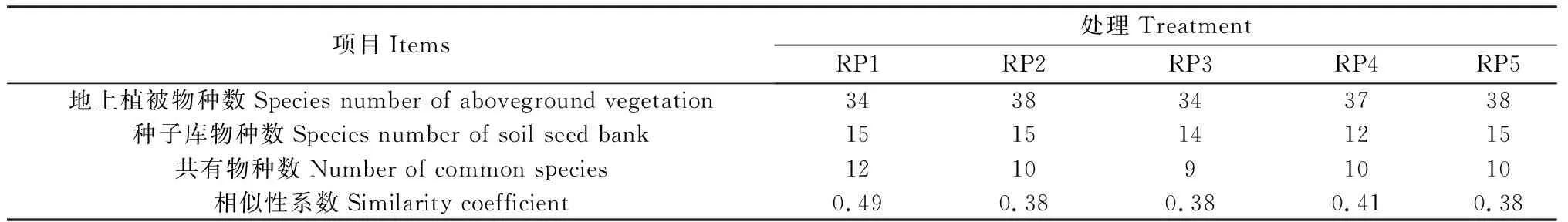
表5 土壤种子库与地上植被相似性Table 5 Similarity between soil seed bank and aboveground vegetation
3 讨论
3.1 不同休牧期土壤种子库物种组成和密度
物种组成决定着生态系统的稳定性、生产力及营养动态等功能[25]。本研究所涉及的5个休牧期土壤种子库中共萌发27种植物,分属12科。各处理中豆科植物种子数量很少,原因是草地豆科可食牧草组成较为单一。此外,阴山扁蓿豆等家畜可食豆科牧草自身的生长特性使其在研究区几乎没有竞争优势,从而导致其未能完成种子繁殖或种子繁殖很少。莎草科植物是高寒草甸的优势物种,但其种子萌发率低,多为持久土壤种子库[26],休眠特性使其长期存在于土壤中,有利于保存种子遗传资源。随着休牧期的推迟,禾本科和莎草科植物种子密度降低,原因是家畜采食、践踏刺激了禾本科、莎草科等优良牧草的分蘖,使其更多的倾向于营养繁殖[7]。而狼毒、高山唐松草和棘豆等含有毒物质,牛羊不采食,加之狼毒的生活史较短,种子成熟早,因此,5个休牧期土壤种子库中均有有毒植物种子。
生活型是植物的一种生态学分类单位,是植物对生境条件长期适应后的一种生长类型[27]。有关高寒区土壤种子库研究表明,种子库中物种的生活型比较单一,一年生和二年生植物较缺乏,其密度占比不超过10%[28]。本研究表明,5个休牧期土壤种子库由地面芽植物、地下芽植物和一年生植物3种生活型组成,其中地面芽植物和地下芽植物较多,一年生植物较少,主要是因为天然草地物种组成多为多年生植物,这是植物为适应当地寒冷干旱的恶劣环境而长期选择、进化以及放牧利用的结果[29];另外,一年生植物比多年生植物休眠时间短,存在于土壤种子库中的一年生植物大多萌发,所以一年生植物相对较少。随着休牧期的提前,土壤种子库中地下芽植物种子密度增多,一年生植物出现,说明RP1提前休牧,可以有效减少放牧对土壤种子库带来的影响,增加种子密度,并且可为一年生植物的生长创造条件[30]。
放牧会影响土壤种子库的密度,这是由于放牧过程中家畜采食导致植物叶面积减少,光合产物降低,从而使结实率下降,加上家畜对植物的生殖枝和种子直接采食,导致种子产量下降,进而引起土壤种子库密度降低[31-32]。草地休牧可以提高土壤种子库密度[33],张建利等[34]研究表明退化山地草地休牧1年和2年后的土壤种子库密度分别为1 850 粒·m-2和2 950 粒·m-2,比放牧草地分别增加了37.9%和119.9%。本研究中,土壤种子库密度随休牧期提前而增加,RP1休牧土壤种子库密度显著高于RP3,RP4和RP5,原因是RP1休牧有利于牧草返青,牧草生长迅速,植物密度和高度有所增加,累积的凋落物可以更好地持有种子[19]。此外,土壤理化性质是影响土壤种子库密度的因素之一[35],解冻期的土壤呈现“表融里冻”或“冻融交替”,土壤表面潮湿,家畜践踏会破碎表层土壤,改变土壤结构或降水后造成沃土流失[7,36],因此RP1休牧土壤种子库密度相对较高。而随着休牧期的推迟,牧草返青家畜因跑青畜蹄会带走部分种子。另外,种子快萌发时家畜践踏也会损伤种子,所以土壤种子库密度降低。
3.2 不同休牧期土壤种子库物种多样性
物种多样性是指一个群落中物种数目的多少及其分布状况,而多样性指数则是度量这两层含义的综合指标[27]。本研究中,5个休牧期土壤种子库Margalef丰富度指数、香农-威纳指数、辛普森多样性指数差异不显著,但RP1休牧土壤种子库丰富度指数、香农-威纳指数、多样性指数均高于RP3和RP4,说明RP1休牧土壤种子库的丰富度和多样性高于RP3和RP4,原因是RP1休牧避免家畜践踏损伤植物地下器官,有利于牧草返青及生长,种子产量较高,因此土壤种子库的丰富度和多样性较高。另外,RP5休牧土壤种子库丰富度指数、香农-威纳指数、多样性指数均高于RP2,RP3和RP4,分析发现RP5休牧垂穗披碱草、线叶嵩草和珠芽蓼等物种的密度比其他休牧期少,但出现了多枝黄耆、甘肃棘豆及委陵菜属植物,原因是RP5休牧可食牧草被家畜反复采食和践踏,影响其生殖生长,种子产量降低,而家畜不食牧草得以繁殖,从而提高了土壤种子库的物种丰富度和多样性。均匀度指数则是RP3和RP4休牧高于RP1和RP2,原因是物种种类减少,其分布的均匀度会增加[37]。
3.3 不同休牧期土壤种子库与地上植被相似性
群落相似性在一定程度上可以反映群落间的相互关系,土壤种子库与地上植被组成完整的植物群落,相似性的研究对群落演替具有重要意义[21]。本研究结果表明,5个休牧期土壤种子库与地上植被的相似性整体较低,这与李春鸣等[29]和马妙君等[38]的研究结果一致。原因是在草地生态系统中,放牧家畜是植物群落组成和稳定性的重要决定因子,它们的采食和践踏行为会以各种各样的形式作用于植物群落[39],从而使土壤种子库和地上植被之间存在一定的差异。土壤种子库与地上植被相似性较低的另一个原因是高寒地区的植物种子小、质量轻,从母株掉落到地表以后[40],因冬季土壤冻结长时间不能进入土壤,留存在草地表面,冬春季强烈的风导致有些种子会被吹走。RP1休牧土壤种子库与地上植被的相似性最高,原因是RP1休牧时间长,可以有效的增加种子的产量和质量,所以土壤种子库与地上植被的相似性增大。
4 结论
随着休牧期的提前,青藏高原东北缘天祝高寒草甸冷季牧场地下芽植物种子密度增加,一年生植物出现,土壤种子库密度增加,土壤解冻临界期—牧草枯黄期休牧土壤种子库密度(2 136.67粒·m-2)显著高于其他休牧期,香农-威纳指数和辛普森多样性指数最高,5个休牧期土壤种子库与地上植被的相似性指数大小在0.38~0.49,其中土壤解冻临界期—牧草枯黄期休牧种子库与地上植被相似性最高。表明土壤解冻临界期—牧草枯黄期休牧更有利于土壤储备植物种子,增加草地的抗干扰能力。
