青藏东缘高寒草甸植物群落结构及功能群特征对长期季节放牧的响应
2022-08-31锁才序孙书存
费 璇, 锁才序, 向 双*, 孙书存
(1.中国科学院山地生态恢复与生物资源利用重点实验室, 生态恢复与生物多样性保育四川省重点实验室, 中国科学院成都生物研究所, 四川 成都 610041; 2.中国科学院大学, 北京 100049; 3.南京大学生命科学学院, 江苏 南京 210023)
我国草地约占国土面积40%,是畜牧业生产的主要物质基础,同时也是重要的生态屏障[1]。放牧是草地主要利用方式之一。上世纪60年代后期开始,随着人口增长、放牧压力逐步增加,草地大面积退化[2]。过度放牧导致草地植物群落多样性下降,优质牧草减少,毒杂草增多,降低草地生产力,从而对土壤养分循环产生影响,限制草地可持续利用与发展[3]。围栏封育与季节性轮牧有力地缓解了放牧造成的草地退化,是对草地进行保护性利用的两种方式。“围栏封育”可提高草地初级生产力及增加优质牧草比例[4-5],但长期封禁会产生草地资源浪费,造成高生产力草地植物密度与物种丰富度损失,降低净初级生产力,从而影响碳、氮、磷草地生态系统循环及利用效率[6-7]。因此开展适度的载畜率放牧或季节性轮牧是对封禁这项草地保护政策的有效补充[8]。
季节性放牧可改变草地植物群落特征,丰富草地植物群落多样性和丰富度,同时提高草地初级生产力。北非干旱牧场在季节性放牧(春、夏季保护,秋、冬季放牧3个月)下植物群落多样性高于禁牧封育与连续放牧[9]。长期的四季放牧模式也会对草地的植物群落特征产生影响,秋季放牧(9月至11月)可增加钙质化草地的指示物种丰富度,从而增加物种多样性,冬季放牧(12月至2月)会显著增加牧草丰富度[10]。与全年连续放牧相比,单一季节放牧(每年6至10月)有助于植物群落中适口植物种类的增加,对其物种多样性的影响小于连续放牧[11]。内蒙古典型草原在夏季放牧(6月与7月下旬)下可提高植物地上净初级生产力并加速生态系统的养分循环[12]。还有研究表明在经过短期(一年)冷、暖季放牧后,相比暖季放牧(6月—10月),冷季放牧有利于青藏高原高寒草地群落地上生物量的积累[13]。因此,季节性放牧可能是逆转过度放牧负面影响的有效生态管理手段之一,其有利于草地恢复和可持续发展[14]。
青藏高原东部高寒草甸是黄河与长江流域重要的生态屏障,其具有调节气候、涵养水源、保护生物多样性等重要生态功能。近几十年来,由于放牧活动的加强和大规模气候变化导致青藏高原高寒草甸植被和土壤退化加速[15]。该地区自2006年开展“牧民定居”工程后,牧民们从原来逐水而居的季节性游牧方式转变为放牧地点固定的季节性放牧[16],季节性放牧的转场时间视不同地区牧草的植物返青和生长发育时期而有所不同。青藏高原东部高寒草甸牧场存在4种放牧方式:1)围栏封育,恢复植被,缓解放牧压力;2)冷季牧场,放牧时间约为11月至翌年5月,主要分布在牧民定居区附近,且靠近水源,放牧范围较小;3)暖季牧场,放牧时间大约为6月至10月,分布较为偏远且道路不畅,但草场植被丰富,范围较广,常为几户或村庄共同使用;4)全年放牧牧场,主要分布牧民定居周边[17-18]。由于该地区地形地貌错综复杂,水热条件分布不均,形成了植被种类丰富的不同小生境类型[19],因此其应对长期季节性放牧的响应可能存在差异。同时植物不同生长阶段对外界干扰敏感程度不同[20],在植物关键生育期放牧会破坏植物营养器官等构件,降低光合产物合成效率,减少生物量;而长期在植物繁殖时期进行放牧,不仅损失繁殖器官构件,同时减少种子产量,影响种子散播,从而影响植物后代种群数量,甚至改变种群结构;且植物的营养生长与繁殖发育具有很强的相互作用,顶端生长开始后受到放牧干扰可能会导致其分蘖的数量下降。因此在植物不同生长阶段进行放牧,不同种类植物可能会产生不同适应策略,从而影响其在群落中优势地位。有研究将季节放牧与常年连续放牧做比较发现季节放牧提高高寒草地的植物群落高度、盖度,以及禾本科、莎草科等牧草产量与冷季放牧地的草地生产力[21]。同时不同季节放牧会改变植物群落优势物种,有研究发现在全年放牧下菊科与毛茛科植物占优势,暖季放牧下植物群落中菊科和禾本科植物占优势,冷季放牧下禾本科、毛茛科及莎草科植物占优势,且冷季放牧显著提高了植物群落高度、地上生物量及地下生物量[17]。大多数研究主要关注不同的放牧模式对植物群落特性影响,且放牧时间较短,然而青藏高原存在复杂且多样的生境类型,不同类型生境的高寒草甸植物群落应对长期季节性放牧在群落结构、优势种群等方面的响应及产生变化的具体原因还尚未清晰。
因此本试验在青藏高原东部地区选取具有10年以上固定季节性放牧历史的3种不同生境类型的草甸,探究不同季节性放牧格局下其植物群落结构、多样性特征和功能群变化,揭示它们之间的关系,阐明群落结构和功能群变化原因,从而理解季节性放牧对青藏高原高寒草甸影响,为青藏高原高寒草甸可持续进行季节性放牧提供科学理论依据。
1 材料与方法
1.1 试验区概况
试验地点位于四川省阿坝州羌族藏族自治州红原县与若尔盖县境内,地处青藏高原东北边缘,地理坐标东经101°51′~103°39′,北纬31°51′~34°19′,平均海拔3 406~3 500 m,属于高原寒温带湿润季风气候,常年无夏,年平均气温0.6至1.2℃,1月最冷月的多年平均气温为—10.6℃至—10.3℃,7月最热月的多年平均气温10.8℃至10.9℃,多年平均降水量600~753 mm,日照时数约为2 159~2 389 h,年平均蒸发量约为1 232 mm,相对湿度约为71%,草地类型为高寒草甸,土壤类型以高山草甸土为主[22]。
1.2 样地设置
根据实地走访牧民和查阅该地区历史资料发现[23],季节性放牧分为冷季放牧(Cold grazing,CG;放牧时间为11月至翌年5月)与暖季放牧(Warm grazing,WG;放牧时间为6月至10月);植物开始返青的时间通常为每年4月[15]。2021年7月在红原县邛溪镇与若尔盖县麦溪乡附近根据地势及地表水分状况选取具有10年以上冷、暖季放牧历史的湿生草甸(Hydric meadow,HM)、中生草甸(Mesophytic meadow,MM)与旱生草甸(Xerophytic meadow,XM)3种类型生境开展试验。湿生草甸所处地势为低洼的宽谷底部,排水性差,地表有临时性或季节性积水,土壤水分含量较高,以莎草科植物为优势物种,主要优势种为木里苔草(Carexmuliensis)、水麦冬(Triglochinpalustre)、钝苞雪莲(Saussureanigrescens)及紫羊茅(Festucarubra);中生草甸所处地势平缓,土层紧实,呈多优势结构,地表湿润,但无明显季节或临时积水现象,主要优势物种为冷地早熟禾(Poacrymophila)、嵩草(Kobresiamyosuroides)、钝苞雪莲及珠芽蓼(Polygonumviviparum);旱生草甸所处地势为缓坡,土层瘠薄,多砾石,表层草根紧盘结,通透性差,地表水分干湿中等,以莎草科与杂类草呈优势,主要优势物种为嵩草、珠芽蓼、苔草(Carexspp.)及狼毒(Euphorbiafischeriana)[24]。每组冷、暖季放牧草场间隔相近,地理环境相似(表1);放牧牲畜主要为牦牛,放牧强度约为1.50~1.79牛·hm-2,放牧强度属于轻度放牧[25]。

表1 样地基本情况Table 1 Basic situation of the sample sites
1.3 植物群落调查与生物量测定
分别在每个生境类型选择典型的冷季放牧与暖季放牧牧场,每个季节性牧场选择3个代表性样地,每个样地随机选择1个50 cm×50 cm的样方参照陆地生物群落调查方法[26]以百格样方框进行植物群落调查测定样方内植物种类、多度、盖度、高度。
完成植物群落调查后,挖取具代表性50 cm×50 cm土方,小心浸泡冲洗保持根系完整后,按照禾本科、莎草科、双子叶杂草及豆科四个功能群分类,将每株植物地上、地下部分分离,称其鲜重,并分别装入信封中,于65℃烘箱中烘至恒重后用万分之一天平称量后记为地上和地下生物量。
1.4 物种重要值与植物群落多样性计算方法
重要值(P):P=(RC+RH+RA+RF+RB)/5
式中:RC表示相对盖度;RH表示相对高度;RA表示相对多度;RF表示相对频度;RB表示相对生物量。
生物多样性用丰富度指数(R),香农-维纳指数Shannon-Wiener(H)、物种均匀度指数Pielou(E)和辛普森指数Simpson(D)进行表征。
R=S
H=-∑(PilnPi)
E=H/lnS
D=1-∑Pi2
式中:Pi为种i的相对重要值,S为种i所在样方的物种总数[27]。
1.5 数据分析
用Microsoft excel 2018进行数据整理及汇总,用SPSS 22.0统计软件对冷、暖季放牧牧场植物群落高度、盖度、香农-维纳指数、辛普森指数、物种均匀度指数、丰富度指数重要值、生物量及重要值等数据进行独立样本t检验分析与单因素方差分析(One-way ANOVA),并且利用Duncan检验比较相同季节放牧下各指标或同一草甸不同功能群之间的显著差异,统计显著水平为P=0.05;用Canoco for window 4.5对冷、暖季草甸植物群落多样性指数、生物量等数据进行主成分分析(Principal components analysis,PCA),对数据进行降维排序,分析不同季节放牧对植物群落结构的组成差异,同一颜色及形状样点距离远近说明样本的重复性强弱,不同颜色点的远近则反映了组间群落差异,用R 4.1.1软件的vegan包进行分析,采用相似性分析(ANOSIM)检验不同季节放牧下植物群落组成差异的显著性,利用Origin 2021软件绘制图表。
2 结果与分析
2.1 季节性放牧对青藏高原高寒草甸植物群落高度与盖度影响
三种生境植物群落高度在季节性放牧下表现相同趋势,暖季放牧下HM植物群落高度高于冷季放牧(P<0.05);季节性放牧下HM植物群落高度呈现高于MM与XW趋势,且在暖季放牧中出现了显著差异(P<0.05),HM与XW在冷季放牧下差异显著(P<0.05)(图1)。MM与XW草场暖季放牧下植物群落总盖度显著高于冷季放牧(P<0.05),而HM草场植物群落暖季放牧总盖度显著低于冷季放牧(P<0.05)。暖季放牧和冷季放牧对植物群落盖度的影响在不同生境的样地呈现出相反的趋势,暖季放牧下HM群落盖度最低,冷季放牧下HM群落盖度最高,均与另两种生境出现显著差异(P<0.05)(图2)。
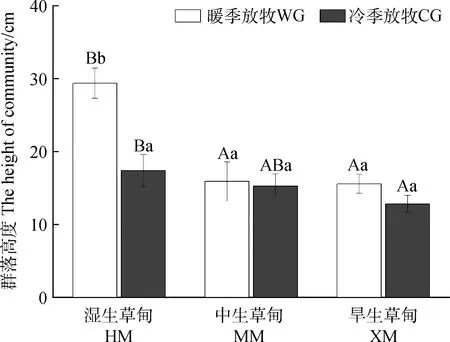
图1 季节性放牧对3种亚高山草甸群落高度的影响Fig.1 Effects of seasonal grazing on the height of plant communities along three subalpine grasslands of Qinghai-Tibetan Plateau注:不同大写字母表示同一季节放牧在不同生境中出现差异显著(P<0.05);不同小写字母表示在同一生境中不同季节放牧出现差异显著(P<0.05)。下同Note:Different capital letters indicated that grazing had significant difference in different habitats in the same season at the 0.05 level;Different lowercase letters indicate significant difference in grazing in different seasons in the same habitat at the 0.05 level. The same as below

图2 季节性放牧对3种亚高山草甸植物群落盖度的影响Fig.2 Effects of seasonal grazing on the coverage of plant communities along three subalpine grasslands of Qinghai-Tibetan Plateau
2.2 季节性放牧对青藏高原高寒草甸植物群落多样性特征影响
HM暖季放牧下香农维纳指数与物种丰富度均低于冷季放牧(P<0.05),而XM暖季放牧植物群落香农维纳指数、辛普森指数、均匀度指数及物种丰富度均显著大于冷季放牧(P<0.05),MM在冷、暖季性放牧下多样指数未出现显著差异。暖季放牧下,MM与XM在香农维纳指数、均匀度指数及物种丰富度植物均显著高于HM(P<0.05),冷季放牧中,HM与MM草场植物群落多样性指数均显著高于XM(P<0.05)(表2)。

表2 季节性放牧对3种亚高山草甸植物群落多样性的影响Table 2 Effects of seasonal grazing on the diversity of plant communities along three subalpine grasslands of Qinghai-Tibetan Plateau
2.3 季节性放牧对青藏高原高寒草甸植物群落生物量影响
长期暖季放牧三种生境牧场地上生物量均显著低于冷季放牧(P<0.05);HM地下生物量高于冷季放牧(P<0.05),而XM地下生物量表现相反,由于地下部分生物量占总生物量比例较高,因此总生物量与地下生物量表现相同。暖季放牧下,HM的地下部分生物量显著高于MM与XM(P<0.05),而地上部分生物量呈相反趋势;冷季放牧下,MM地上生物量最高,其次为XM与HM,XM的地下部分生物量显著高于MM与HM(P<0.05)(图3)。

图3 季节性放牧对三种亚高山草甸植物群落地上、地下生物量影响Fig.3 Effects of seasonal grazing on the aboveground and underground biomass of plant communities along three subalpine grasslands of Qinghai-Tibetan Plateau
2.4 季节性放牧对植物群落不同功能群重要值影响
MM与HM中暖季放牧下莎草科重要值显著高于冷季放牧(P<0.05),而双子叶杂草重要值显著低于冷季放牧(P<0.05),HM中禾本科重要值显著低于冷季放牧(P<0.05);XM草场在冷暖季放牧下不同功能群重要值与HM呈现相反趋势,其暖季放牧下禾本科、双子叶杂草及豆科重要值均显著高于冷季放牧(P<0.05),而莎草科重要值显著低于冷季放牧(P<0.05)。在暖季放牧下,MM与XM双子叶杂草重要值最高,分别为0.55与0.51,显著高于同一草场其他功能群(P<0.05),而HM莎草科重要值最高,为0.43;冷季放牧下,MM与HM双子叶杂草重要值最高,分别为0.66与0.47,而XM莎草科重要值最高,为0.51(图4)。

图4 季节性放牧对3种亚高山草甸植物群落不同功能群重要值影响Fig.4 Effects of seasonal grazing on the importance values of different functional groups along three subalpine grasslands of Qinghai-Tibetan Plateau注:不同大写字母表示相同样地中不同功能群出现差异显著(P<0.05);不同小写字母表示在同一生境中不同季节放牧,相同功能群出现差异显著(P<0.05)。下同Note:Different capital letters indicated significant difference in different functional groups in the sample plot at the 0.05 level;Different lowercase letters indicate grazing in the same habitat in different seasons,and there were significant differences among the same functional groups at the 0.05 level. The same as below
2.5 季节性放牧对植物群落不同功能群地上部分生物量影响
HM与MM生境中冷季放牧禾本科地上生物量均显著高于暖季放牧(P<0.05),HM与MM草场冷季放牧下莎草科地上部分生物量显著低于暖季放牧(P<0.05),XM莎草科地上部分生物量呈相反情况,其双子叶杂草地上部分生物量显著低于暖季放牧(P<0.05)。暖季放牧下,三种生境的莎草科地上部分生物量比例最高,显著高于其他功能群;冷季放牧下,三种生境中不同功能群地上生物量表现规律不同(图5)。

图5 季节性放牧对3种亚高山草甸的不同功能群地上部分生物量比例影响Fig.5 Effects of seasonal grazing on the proportion of different functional groups of plant communities along three subalpine grasslands of Qinghai-Tibetan Plateau
2.6 季节性放牧对植物群落结构影响
将植物群落多样性指数、地上地下生物量及不同功能群重要值等相关数据进行主成分分析并利用相似性分析(ANOSIM)检验长期冷、暖季放牧下植物群落结构差异。结果表明,不同生境草场在长期不同季节放牧群落结构均出现差异。在冷、暖季放牧下,三组草场群落结构存在极显著差异(P<0.01),R值分别为0.877与0.761,且主成分分析第一轴与第二轴能解释总方差比例为76.66%,第一轴共解释群落方差变化的58.09%,第二轴群落方差变化为18.57%,双子叶杂草、豆科重要值与香农维纳指数、丰富度指数相关性高,莎草科重要值与地下生物量具有较强相关性;同时季节性放牧对HM与XM植物群落影响较大,冷、暖季放牧下群落结构分离距离较大,对MM影响较小,冷、暖季放牧下群落结构分离距离较小(图6)。

图6 季节性放牧对3种亚高山草甸植物群落特征(A)及结构(B)影响主成分分析散点图Fig.6 Scatter plot of principal component analysis on plant community characteristics(A) and structure(B) of three subalpine meadows under seasonal grazing
3 讨论
3.1 长期季节性放牧对青藏高原高寒草甸植物群落多样性特征影响
植物群落高度、盖度及多样性指数能直观反映草地植被特征。研究结果显示,长期暖季放牧下湿生草甸植物群落高度高于冷季放牧,其群落盖度及物种丰富度低于冷季放牧,而旱生草甸群落盖度则呈相反趋势。由于生境不同,长期季节性放牧下植物群落特征结果存在差异。湿生草甸多样性特征研究结果与Ridding等[10]研究结果相似,旱生草甸多样性特征研究结果与刘玉等[13]、Zhang等[11]的研究结果相似。说明不同生境草甸植物群落特征应对长期季节性放牧的响应不同。长期暖季放牧下,湿生草甸植物群落高度高于冷季放牧,由于生境水分含量较高,适宜莎草科及部分湿生禾草生长,木里苔草、水麦冬、发草等个体高大的优质牧草在群落中占优势地位,并且该植物返青期受放牧干扰影响较小,因此长势较高,同时说明生境对其影响可能大于长期以来放牧对其的干扰。而又因为牲畜的偏食性,对草玉梅、珠牙廖及黄帚橐吾(Ligulariavirgaurea)等双子叶杂草的取食性较低,双子叶杂草在群落中长势较高,一定程度上提高植物群落高度。中生与旱生草甸中,暖季放牧增加了植物群落盖度并且提高了旱生草甸植物群落多样性特征,长期在植物生长旺盛期进行放牧干扰,牦牛等牲畜会偏向取食长势较高禾本科或莎草科植物等适口性较好的牧草[28],使得底层非优势物种竞争资源获得发展机会,从而丰富植被盖度,同时增加植物群落物种的多样性[13];同时暖季放牧中牲畜扰动会促进成熟种子传播,增加物种传播的机率,提高植物群落多样性特征。而冷季放牧中,生长季植物自由生长加剧植物物种间的竞争效应,减少物种多样性[29];同时有研究发现冷季牧场凋落物显著高于暖季牧场[30],而凋落物的积累可能会阻碍草地生态系统中植物的繁殖和更新,从而使植物群落组成发生变化,甚至导致生物多样性的丧失[6]。因此不同季节放牧会影响植物群落优势物种长势及多样性特征,进而影响植物群落结构。
3.2 长期季节性放牧对青藏高原高寒草甸植物群落生物量影响
植物群落生物量大小代表草地生产力,同时也是草场载畜能力的基础[31]。长期在不同季节进行放牧干扰,不同种类生活型植物会产生不同的资源分配方式,导致两种放牧制度下草地初级生产力之间出现差异。研究结果显示,长期暖季放牧下植物群落的地上生物量均低于冷季放牧,湿生草甸与中生草甸暖季放牧植物群落地下生物量高于冷季放牧,而旱生草甸则相反。冷季放牧植物在返青时期受到干扰影响较大,但其在生长季时未受到放牧干扰,冷季牧场植物具有一定时间的进行恢复生长,因此冷季牧场地上生物量高于暖季放牧[17]。然而不同生境草甸植物群落地下生物量应对季节性放牧产生响应不同。Angassa[32]认为草本物种丰富度在中等水平的生物量中最高,随着生物量的增加而下降。暖季放牧由于牦牛等牲畜对草甸植物个体或部分构件的直接啃食以及踩踏,导致草场初级生产力直接损失,同时,由于在生长季受到放牧取食压力,植物可能会产生避牧繁殖策略,将大量光合产物转移至地下,从而提高植物群落地下生物量[29];而冷季牧场植物在生长旺盛期未受到放牧直接取食压力,因此冷季放牧下植物群落具有较高的地上生物量。旱生草甸在季节性放牧中地上、地下生物量分配结果与Wu等[29]探究季节性放牧对高寒草甸物种多样性保护和表土养分固存的结果一致。冷季放牧在植物生长的时期受到扰动较少,在植物生长高峰期获得丰富的地上部分物质积累,待生长季结束后,将大量营养物质转移、储存至地下部分,为植物第二年生长做好储备;同时由于旱生草甸冷季放牧中苔草在植物群落中占显著优势地位,莎草科植物对地下部分生物量分配更高[33],因此冷季放牧有助于旱生草甸地下生物量的积累。长期冷季放牧其植物群落地上生物量高于暖季放牧,但不同生境草甸地下部分生物量对季节性放牧的响应不同,说明季节性放牧能促进物质与能量向地下部分转移,这对草地土壤有机质含量与增加土壤肥力具有积极意义。
3.3 长期季节性放牧对青藏高原高寒草甸不同植物功能群特征影响
功能群通常被认为与某种生态系统服务功能相关的一类特定的物种群,就草地物种组成而言,功能群决定着生态系统的生产力和稳定性[34]。研究结果显示季节性放牧下不同生境占优势地位的功能群是不同的,同时暖季放牧下各个生境中莎草科地上部分生物量在群落中所占比例最高,而冷季放牧中并未表现一致规律。长期暖季放牧群落中莎草科地上生物量比例较高;相较于暖季放牧,冷季放牧下禾本科地上部分生物量比重增加,同时季节性放牧在一定程度上使双子叶杂草在群落中占据优势地位。该结果与齐洋等[35]、Wu等[29]研究的结果类似。在一定放牧压力下,莎草科植物趋向无性繁殖(如根蘖)方式,可在植物群落中快速繁殖提高其地上生物量比例。同时每年4—6月是高寒草地植物返青时期,部分禾本科(如垂穗披碱草、发草、冷地早熟禾等)与莎草科(如嵩草、矮藨草(Scirpuspumilus)、苔草等)植物返青时间较早,豆科与双子叶杂草类植物返青期相对较晚[36],牦牛等牲畜会对长势较高的禾本科植物等进行取食,使群落下层呈莲座状且长势较矮的莎草科与双子叶杂草物种获得生长资源,从而增加其物种竞争优势[30],双子叶杂草植物(如狼毒、甘肃马先蒿(Pediculariskansuensis)、鹅绒委陵菜等)具有很强的繁殖能力且耐践踏,很少被家畜采食[13],因而季节性放牧使双子叶杂草植物在植物群落中占据重要地位。牦牛的消化作用有利于禾本科植物种子的萌发,莎草科部分种子由于其自身的深度休眠而不受影响[17]。冷季放牧中牦牛等牲畜在植物群落成熟时期取食了更多的植物种子,促进植物群落有性繁殖种子传播并提高禾本科植物发芽率,相较于暖季放牧,冷季放牧禾本科地上生物量在一定程度上有所增加。
3.4 长期季节性放牧对青藏高原高寒草甸群落结构影响
长期季节性放牧是在植物不同生长阶段对其进行干扰,而植物不同生长阶段对外界干扰敏感程度不同,植物繁殖期尤为敏感[20],因此长期季节性放牧下不同种类植物可能会产生不同适应策略,从而改变植物群落结构。研究结果显示,三种生境植物群落结构在季节性放牧下均产生显著差异,湿生草甸与旱生草甸群落结构差异较大,而中生草甸植物群落结构差异较小。长期季节性放牧下,植物群落多样性提高与双子叶杂草及豆科功能群物种增加有关,莎草科功能群的群落优势地位对地下生物量会产生影响[33]。且不同生境草甸应对长期季节性放牧敏感程度不同,湿生草甸与旱生草甸在长期不同季节放牧影响下敏感程度较高,植物群落结构差异较大。地形差异会使生境水热条件产生差异,形成的多种多样微气候环境,影响植物群落多样性特征,改变种间竞争,从而间接影响群落的组成[37]。湿生草甸地形为洼地,存在季节性积水,旱生性草甸地形为缓坡,其在小区域内水分与热量差异较大,土壤水分的减少可能限制了莎草和一些适应潮湿土壤条件的禾草及双子叶杂类草的生长,有利于耐旱物种生长[38],加上季节性放牧因素影响,因此可能加剧植物群落结构分化。Adler等[39]研究了美国西北矮草草原(Sagebrush steppe)和阿根廷巴塔哥尼亚草原(Patagonia steppe)发现,越靠近水域,植物群落对放牧的响应越明显,而中生草甸地势平坦,在其区域微环境内水热条件差异不大,因此其可能在长期季节性放牧影响下植物群落结构差异较小。
4 结论
长期季节性放牧会对青藏高原东部高寒草甸植物群落多样性、生物量、功能群特征及群落结构产生影响,然而不同生境类型高寒草甸的群落特征对长期季节性放牧的响应不同,这可能与其生境地势及土壤水分条件有关;在长期的季节性放牧下,湿生与中生草甸的群落结构表现较大差异,而中生草甸群落结构表现差异较小。因此未来可根据不同生境类型草甸对季节性放牧的响应特点设置合理的放牧格局,并可将冷、暖季牧场进行周期性地交替使用,这有利于青藏高原高寒草甸放牧生态系统可持续利用与发展。
