黄芩MYB转录因子家族全基因组鉴定与分析
2022-08-24王晓童高冉冉
王晓童 向 丽 邬 兰 高冉冉
(1 中国中医科学院中药研究所/青蒿素中心,北京,100700; 2 东北林业大学生命科学学院,哈尔滨,150040)
黄芩(Scutellariae Radix)是我国传统中药,具有清热燥湿、泻火解毒的功效,其基原植物为唇形科(Lamiaceae)植物黄芩(ScutellariabaicalensisGeorgi)[1]。研究表明黄芩地上和地下部分积累不同的黄酮类活性成分,地上主要积累野黄芩苷、野黄芩素、芹菜素、木樨草素等,地下主要积累黄芩苷、汉黄芩苷、黄芩素、汉黄芩素等4′-脱氧类黄酮[2]。黄芩及其黄酮类活性成分均具有良好的临床药用价值,在新型冠状病毒肺炎的治疗中,黄芩作为传统中药发挥重要作用,是《新型冠状病毒诊疗方案》中“清肺排毒汤”“宣肺润燥解毒方”的成分之一[3]。现代药理研究表明,黄芩提取物可以体外抑制新型冠状病毒(SARS-CoV-2)中的蛋白酶3CLpro活性和SARS-CoV-2在Vero细胞中的复制[4]。此外,黄芩素及黄芩苷等成分被报道具有改善饮食引起的肥胖和肝脂肪变性、抗菌等活性。因此,揭示黄芩中调控黄酮类活性成分生物合成的转录因子及其调控机制,对于培育高黄酮含量的黄芩种质、黄芩相关药物研发等具有重要价值。
MYB转录因子(v-Myb禽成髓细胞病病毒癌基因同源物)是植物界中数量最多的转录因子家族之一,其在N端含有一段能够与DNA结合的高度保守的基序,该基序通常由1~4个不完整的重复片段组成,每个重复片段50~53个氨基酸,组成3个α-螺旋并形成螺旋-转角-螺旋(Helix-turn-helix,HTH)结构,MYB转录因子通过HTH结构插入到靶DNA的大沟中进行结合,从而调控靶基因的表达。依据重复序列的数量和种类,MYB可分为4个亚族:1R-MYB、2R-MYB(R2R3-MYB)、3R-MYB和4R-MYB,R2R3-MYB是数量最多、功能也最为广泛的亚族[5-7]。
MYB转录因子是目前研究较多的转录因子家族,广泛参与调控植物的生长发育、抗逆胁迫、次级代谢产物的积累。已报道研究中,MYB参与调控植物黄酮类成分合成的报道居多,如苹果(Malusdomestica)MdMYB9和MdMYB11都能与花青素合成酶(Anthocyanin Synthetase,ANS)、花青素还原酶(Anthocyanidin Reductase,ANR)和无色花青素还原酶(Leucoanthocyanidin Reductase,LAR)的启动子结合,调控花青素和原花青素的积累[8];蒺藜苜蓿(Medicagotruncatula)中的MYB转录因子MtLAP1可以结合MtGSTF7启动子区来激活其表达,从而调控蒺藜苜蓿中花青素的积累[9];拟南芥(Arabidopsisthaliana)、龙胆(Gentianatriflora)中的MYB转录因子也可以调控黄酮类成分的积累[10-12]。
迄今为止,黄芩活性成分合成的调控机制鲜有报道。目前报道的黄芩中参与黄酮类化合物合成的SbMYB只有3个[13-14]。随着生物技术的快速发展,黄芩的全基因组测序已经完成[15-16],黄芩中黄酮类活性成分的生物合成途径及关键酶已基本解析[17-19],基于全基因组的SbMYB转录因子家族的系统分析和研究提上日程。本研究首次基于黄芩的全基因组对其中MYB转录因子进行系统鉴定和分析,预测可能参与黄酮类化合物生物合成相关的调控基因,为深入研究黄芩中黄酮类成分合成的调控机制提供支撑。
1 材料与方法
1.1 基因家族的鉴定 黄芩基因组在the Genome Warehouse(https://bigd.big.ac.cn/gwh)下载(登录号:GWHAOTO00000000)。采用以下2种方法进行SbMYB转录因子家族成员的鉴定:1)从PFAM数据库(http://pfam.xfam.org/)下载MYB转录因子特有的保守结构域模型(PF00249),使用HMMER软件在黄芩基因组数据库中进行搜索,E-value设为e-5;2)从植物转录因子数据库(http://planttfdb.gao-lab.org/index.php)下载168条拟南芥AtMYB蛋白序列,通过本地BLAST,与黄芩基因组数据库进行同源比对。将二者的比对结果整合,使用NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)和PFAM蛋白数据库进行保守结构域预测。通过PROSITE(http://www.expasy.org/prosite/)针对筛选出的SbMYB转录因子进行蛋白质理化性质预测。通过WoLF PSORT(https://wolfpsort.hgc.jp/)对SbMYB进行亚细胞定位分析。
1.2 基因家族分类与系统发育树 为研究黄芩MYB转录因子的系统进化关系并确定其亚族分类,以模式物种拟南芥为参考,利用最大似然法对168条AtMYB和146条SbMYB蛋白的全长序列构建系统发育树。序列与MAFFT进行比对,比对结果使用IQ-TREE2.0.3构建最大似然法系统发育树,MFP参数选择最优模型,BOOTSTRAP设为1 000次。系统发育树在ITOL在线网站(https://itol.embl.de/)进行编辑。根据黄芩与拟南芥MYB转录因子的系统发育关系,对黄芩SbMYB基因家族成员进行亚族分类。
1.3 基因结构与染色体分布分析 从黄芩基因组注释文件中提取146个SbMYB基因的注释信息,将其提交到GSDS2.0(http://gsds.gao-lab.org/index.php)网站进行基因结构分析,确定基因的内含子数量,并绘制内含子和外显子模式图。从SbMYB基因注释文件中获取基因位置信息,并进行统计整理,使用Tbtools进行可视化分析。
1.4 保守结构域与基序分析 采用基序诱导多个最大期望值(Multiple Expectation Maximization for Motif Elicitation,MEME)在线软件(http://meme-suite.org/)对SbMYB的保守基序进行分析。参数设置为:同一基序在一条序列中出现的次数为“any”,基序长度范围6~100个氨基酸残基,基序最大发现数目15个,其他参数为默认值。将以上结果与PFAM数据库获得的保守结构域信息一同提交到ITOL网站进行可视化分析。
1.5 基因表达模式分析 根据黄芩SbMYB转录因子在根、茎、叶和花4个不同组织中编码基因的表达量FPKM值,绘制SbMYB转录因子在不同组织部位的表达量热图。黄芩中黄酮合成途径结构基因已经被注释[17],筛选15个已验证有功能或表达量高的黄酮类化合物合成相关基因(SbPAL3、SbC4H1、SbCLL-1、SbCLL-5、Sb4CLL7-1、Sb4CLL8、Sb4CLL10、SbCHS1、SbCHI、SbFNSII1、SbFNSII2、SbF6H1、SbF8H、SbF6H2、SbUBGAT),根据已有的转录组数据,提取这15个基因在根、茎、叶和花中的表达量,与SbMYB转录因子绘制表达量热图。所有热图均在在线网站(https://www.omicstudio.cn/tool)绘制,FPKM值进行“+1”预处理后,取log2值,并对基因进行聚类分析。
1.6 基因调控网络构建 为了进一步研究SbMYB转录因子与黄酮类化合物合成相关结构基因间调控关系,本研究将所有SbMYB和高表达的黄酮类化合物合成相关的结构基因蛋白序列提交String(https://www.string-db.org/)进行蛋白质-蛋白质相互作用(Protein-protein Interaction,PPI)预测,预测结果使用Cytoscape 3.8.0构建PPI网络图。筛选FPKM值大于5的SbMYB转录因子,使用Omicstudio平台的Cor工具计算SbMYB转录因子与黄酮类化合物合成相关结构基因间的相关性,构建无向量网络图。相关性系数绝对值大于0.8,皮尔逊相关系数(p-value)小于0.05。
2 结果
2.1 黄芩MYB转录因子家族的筛选与鉴定 本研究共鉴定得到146条SbMYB蛋白序列。蛋白质理化性质和亚细胞定位分析表明,各成员的氨基酸序列长度在87 aa(SbMYB106)~957 aa(SbMYB105)之间,平均分子质量在9 884.46(SbMYB109)~107 846.15(SbMYB105)之间。蛋白质等电点大于(碱性蛋白质)和小于(酸性蛋白质)7的成员数量分别为90和56个,说明大多数为碱性蛋白质。脂肪系数均小于100,亲水系数均为负值,不稳定系数小于40的蛋白序列仅5个(SbMYB55、SbMYB81、SbMYB109、SbMYB117、SbMYB137),表明SbMYB蛋白多为亲水性不稳定蛋白。WoLF PSORT预测结果显示,144个SbMYB定位在细胞核中,SbMYB143、SbMYB120分别定位在线粒体和叶绿体中。见表1。
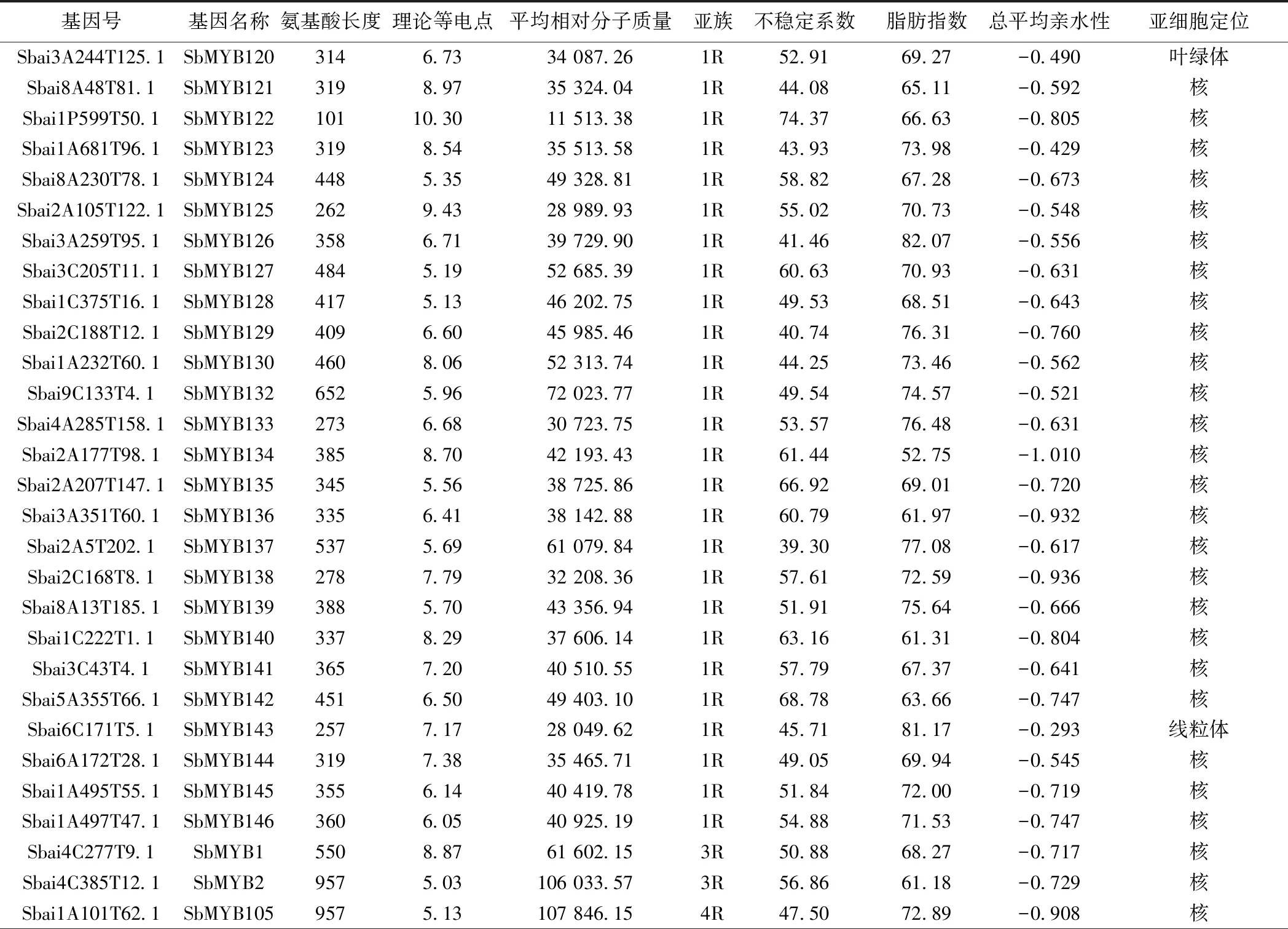
表1 黄芩MYB转录因子家族信息
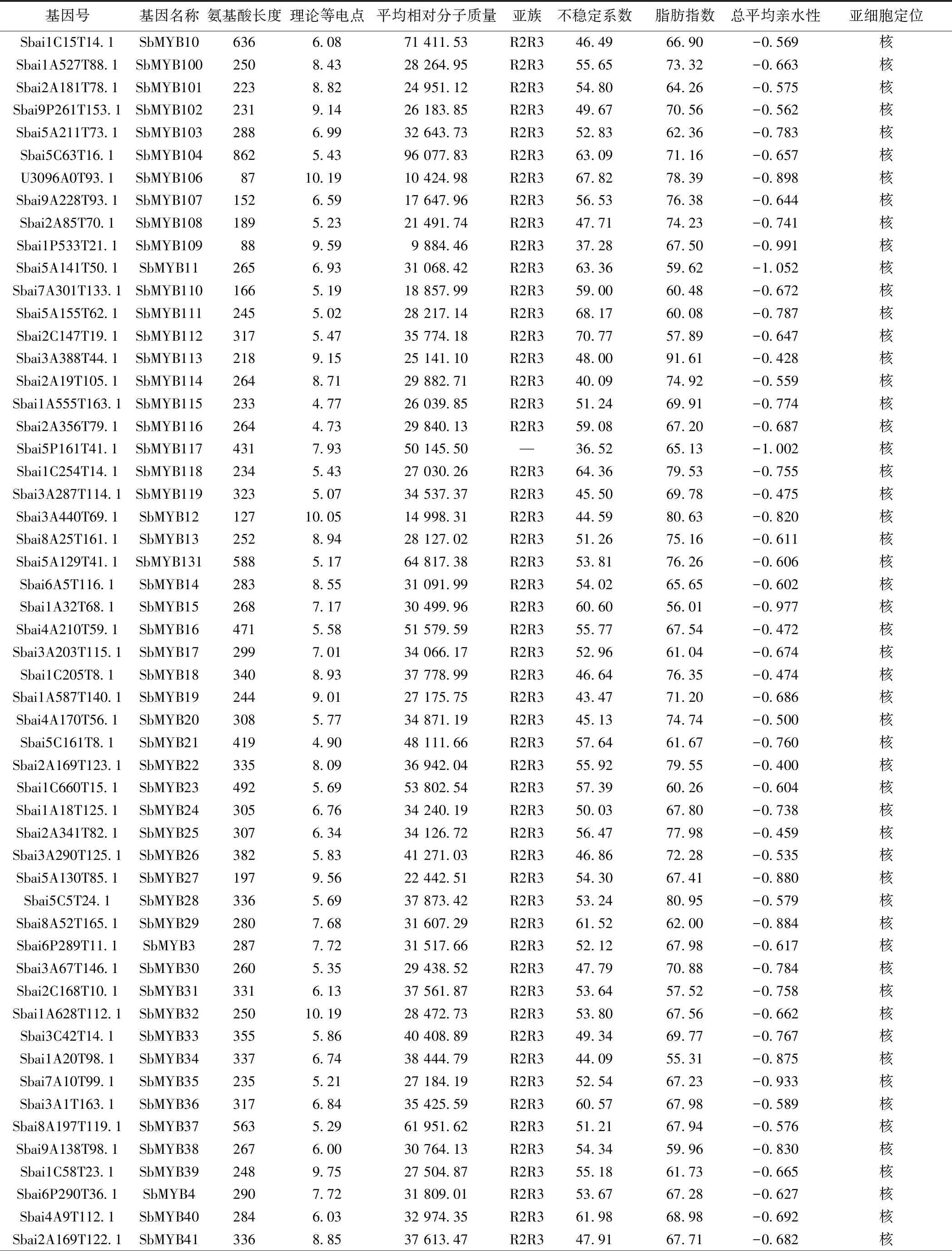
续表1 黄芩MYB转录因子家族信息
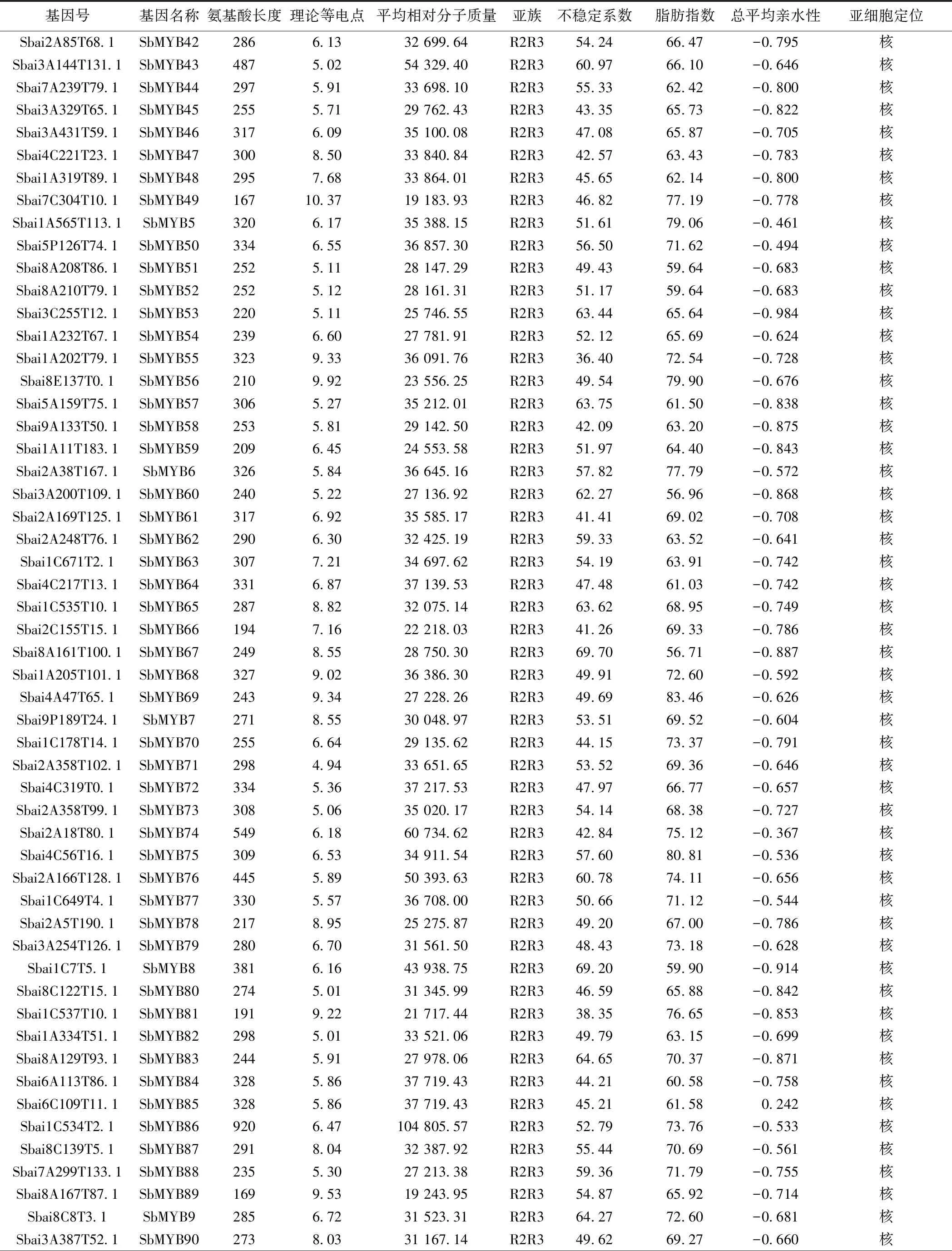
续表1 黄芩MYB转录因子家族信息
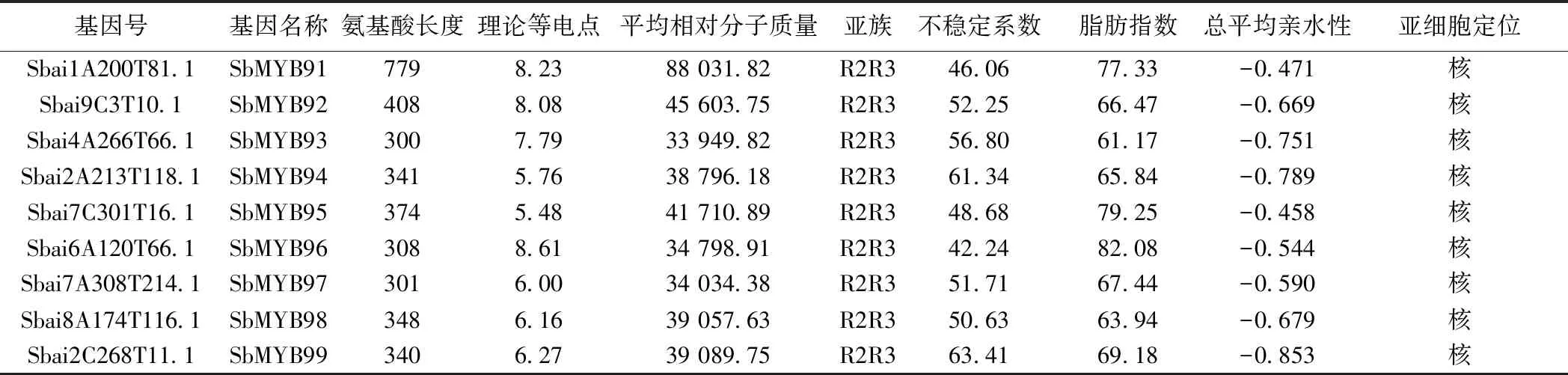
续表1 黄芩MYB转录因子家族信息
2.2 黄芩MYB转录因子家族的系统进化分析 最大似然法构建拟南芥和黄芩MYB蛋白全长序列系统发育树,自动选择最佳模型为VT+R6,进化树分枝上的圆圈代表支持率(%),圆圈越大,相应的支持率也增大。结果显示,根据与拟南芥的系统发育关系,黄芩146个SbMYB转录因子可以划分为4个亚族(1R-MYB,R2R3-MYB,3R-MYB,4R-MYB)和一个未分族成员(SbMYB117)。其中,R2R3-MYB亚族成员数量最多,有116个,占总SbMYB转录因子的79.45%;1R-MYB有26个,占总SbMYB的17.81%,是黄芩SbMYB家族中的第二大亚族;3R-MYB、4R-MYB亚族成员数量较少,分别有2个和1个。见图1。
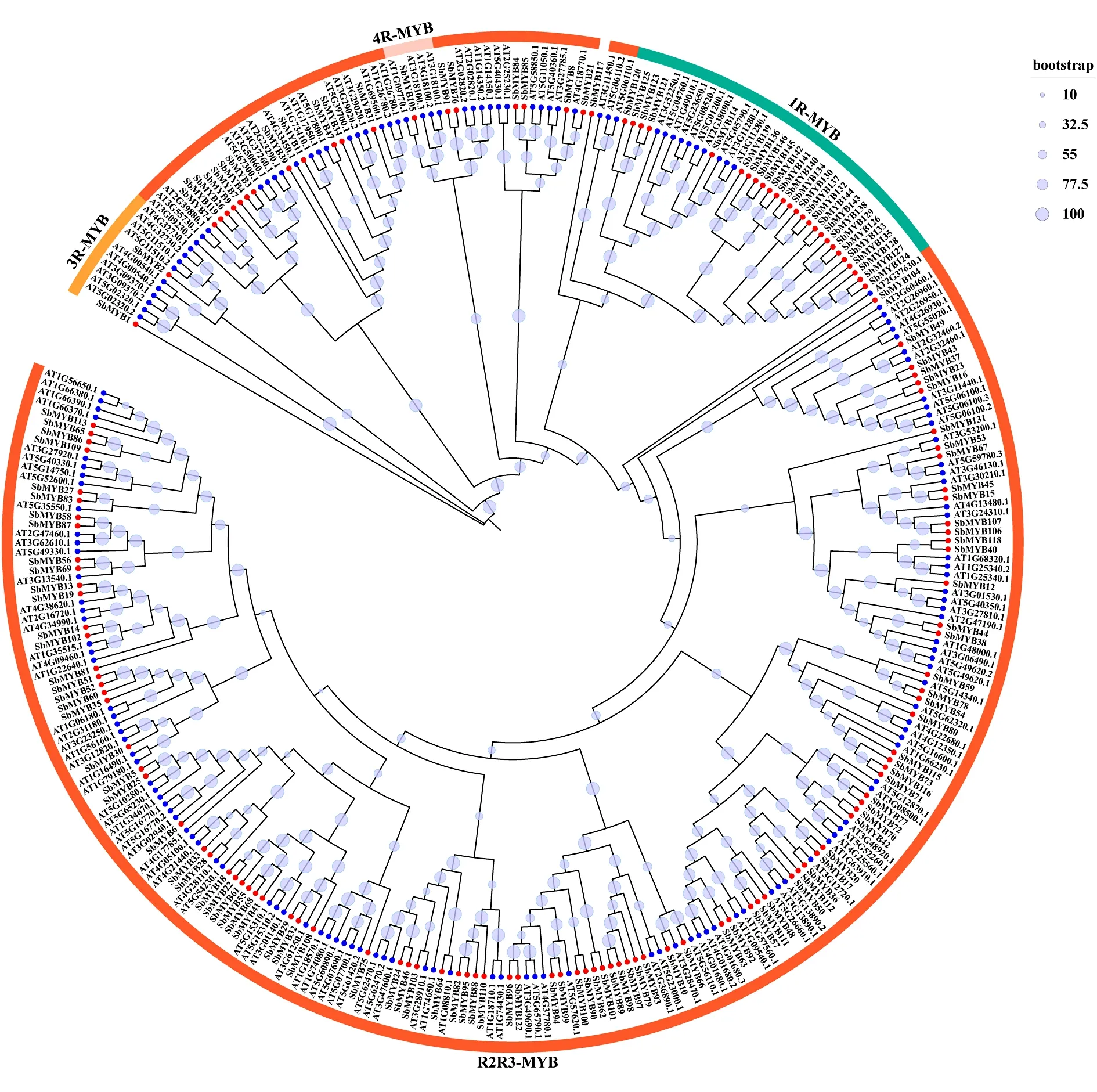
图1 黄芩和拟南芥MYB转录因子家族系统进化树
2.3 黄芩MYB转录因子结构分析及其染色体定位 根据基因组注释信息,绘制了SbMYB转录因子的外显子和内含子模式图。SbMYB外显子数目2~19个,表明不同的SbMYB成员在基因结构上存在多样性和显著差异。同一SbMYB亚族内的外显子数量较为保守,R2R3-MYB亚族成员大都含有2~3个外显子,外显子长度和位置也较为相近;1R-MYB、3R-MYB、4R-MYB亚族成员的外显子数目明显多于R2R3-MYB,大部分含有4~6个外显子。不同亚族之间外显子的位置和长度存在差异。见图2A。

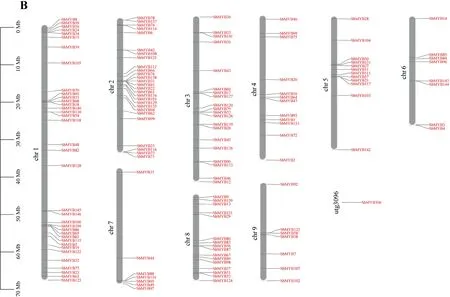
图2 SbMYB的基因结构(A)与染色体分布(B)
依据从黄芩基因组注释文件中获取的SbMYB位置信息,绘制SbMYB染色体定位图。145个SbMYB转录因子随机不均匀分布在9条染色体上(chr1~chr9),其中大多数SbMYB定位在chr1和chr2上(36个和27个),chr6(8个)、chr7(7个)和chr9(7个)上分布较少,SbMYB106未定位到染色体上,而定位在未挂载到染色体的contig上,这与基因组的染色体组装有关。见图2B。
2.4 黄芩MYB转录因子保守结构域与基序分析 本研究对SbMYB进行保守结构域分析,共预测到4个MYB转录因子保守结构域,命名为MYB Domian1-4。所有的SbMYB至少含有1个MYB保守结构域。1R-MYB大都仅含1个MYB Domian1,部分成员还含有MYB Domian3,且MYB Domian3仅在1R-MYB中存在。R2R3-MYB中,最保守的结构域组成含有2个MYB Domian1,很少部分成员仅含有1个MYB Domian2。3R-MYB均含有3个MYB Domian,但组成不同,SbMYB2含有1个MYB Domian2。见图3A。
同一基因家族的成员,其基序组成通常较为保守,且进化关系较近的成员间具有相似的基序组成。我们在SbMYB转录因子中共预测到15个保守基序(Motif),命名为Motif1-15。不同亚族之间SbMYB基序组成存在差异,同一亚族SbMYB成员具有相似数目和类型的基序组成。1R-MYB亚族的保守基序为Motif7,它可能与Motif5或Motif13共同组成MYB Domian3;R2R3-MYB亚族中,Motif1、Motif2、Motif3和Motif4是其最保守的基序,依照其在基因位置上与MYB结构域的对应关系,Motif1-4可能是MYB Domian1的代表基序;3R-MYB亚族的保守基序为Motif1、Motif2、Motif3和Motif11;4R-MYB亚族仅预测到Motif3。见图3B。
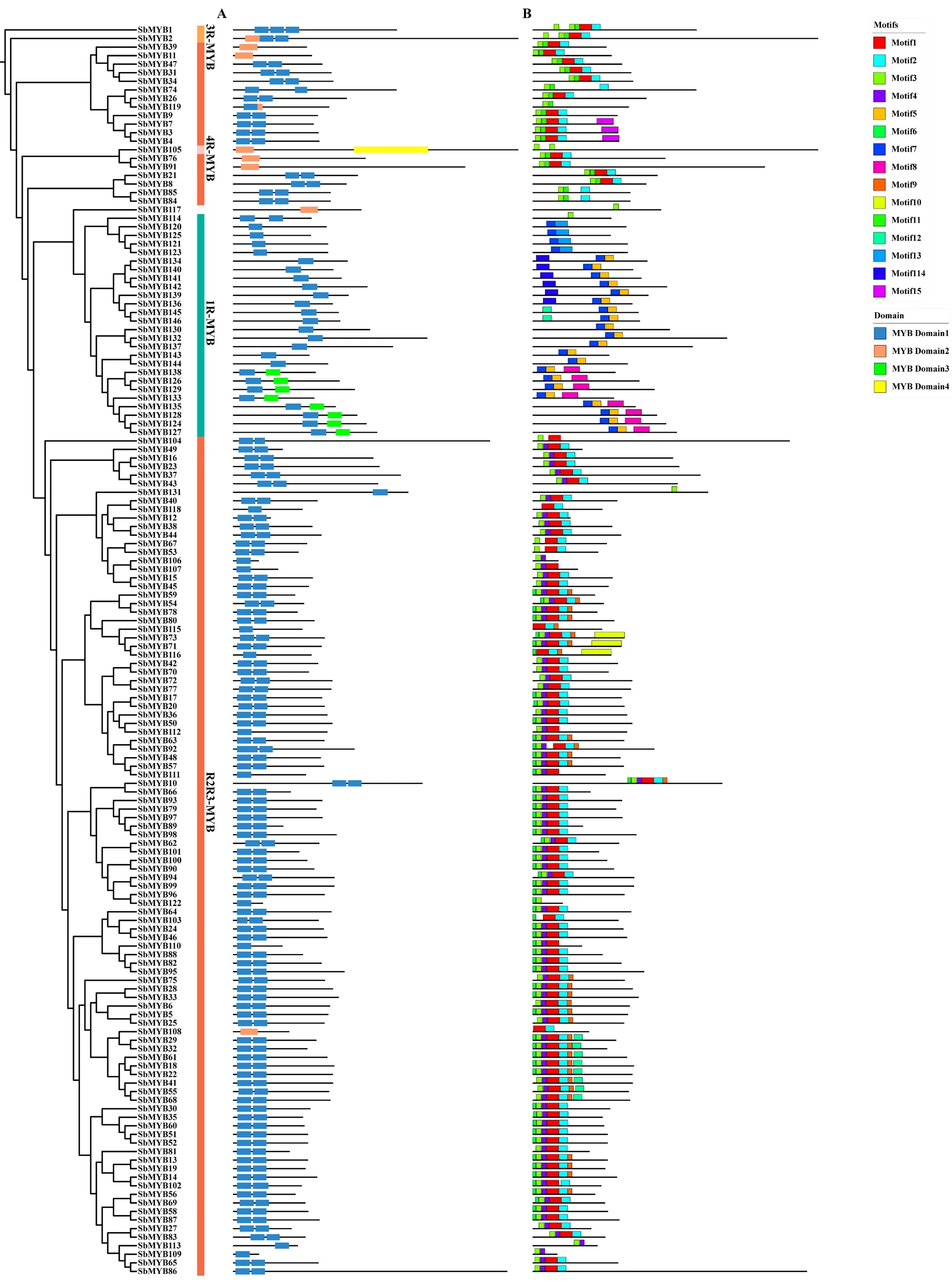
图3 黄芩MYB基因保守结构域(A)与基序(B)分析
2.5 黄芩MYB转录因子的基因表达模式分析 基于黄芩根、茎、叶、花的转录组数据,进一步分析SbMYB在不同器官中的表达模式,结果表明,SbMYB在不同器官中具有不同的表达模式。根、茎、叶中表达模式相似,与花中的表达模式差异较大。见图4A。对SbMYB与黄酮类化合物合成结构基因之间的表达模式分析显示,黄酮类化合物合成相关基因大都与R2R3-MYB聚类在同一枝,表明R2R3-MYB亚族更可能参与黄酮类化合物的调控。此外Sb4CLL10、Sb4CLL7-1、SbFNSII1分别与SbMYB120、SbMYB127、SbMYB126聚在同一枝,说明黄酮类化合物合成相关基因可能也受到1R-MYB的调控。见图4B。
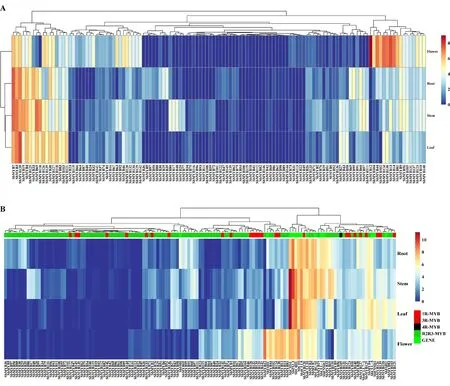
图4 不同组织中的基因表达分析
2.6 黄芩MYB转录因子与黄酮类化合物合成相关结构基因调控网络 通过在转录组水平上对SbMYB与15个黄酮类化合物合成相关结构基因间的相关性进行分析,构建共表达网络,结果表明,15个黄酮类化合物合成相关基因均与43个SbMYB存在强相关性(|r|>0.8),说明它们之间可能存在调控关系。大多数SbMYB(38个)具有正调控作用,少部分SbMYB(9个)具有负调控作用。根据节点大小可以看出,SbCHI可能受SbMYB强调控作用,且大多为正调控。见图5A。PPI网络分析结果表明,有8个黄酮类化合物合成相关基因与SbMYB可能存在相互作用关系;部分SbMYB之间也有潜在的互作关系(如:SbMYB100和SbMYB117),说明SbMYB之间可能形成蛋白复合体共同调节下游基因的表达。见图5B。
3 讨论
药用植物活性成分是发挥其药效的重要基础,主要来源于萜类、黄酮类、生物碱类等次生代谢产物的生物合成。由于植物次生代谢产物含量少、复杂多样、分离困难等因素限制了药用植物的发展和应用。研究表明,转录因子是植物代谢合成过程中的重要调控元件[20],通过与序列特异性DNA结合和PPI调节特定基因表达,可以作为基因表达的激活剂或抑制剂,参与次生代谢物的合成和积累,获取高价值次生代谢产物[21-22]。
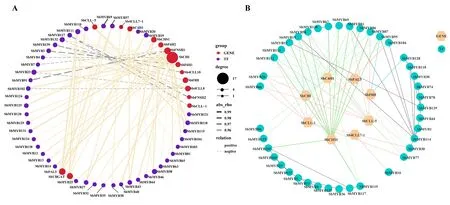
图5 SbMYB转录因子与黄酮类化合物合成相关基因间的调控网络
随着黄芩基因组的公布,黄芩的WRKY转录因子家族已被系统鉴定与分析[23]。MYB是植物中家族最庞大的转录因子家族之一,以R2R3-MYB亚族数量最多,R2R3-MYB也是报道具有最多生物学功能的亚族[24]。本研究基于黄芩的基因组数据,注释出146个MYB转录因子,其中R2R3-MYB亚族有116个,1R-MYB、3R-MYB、4R-MYB亚族成员分别有26个、2个和1个,这与拟南芥、水稻、葡萄等物种中各亚族MYB的数量分布情况相似[5-6,25];保守结构域和基序分析表明,不同亚族间具有不同的结构域和基序组成,这可能与MYB转录因子功能和调控作用方式有关。如:R2R3-MYB和3R-MYB通过R2、R3 MYB结构域特异结合DNA序列,而1R-MYB一般仅含1个MYB保守结构域,可能以不同的方式结合到DNA序列上,如与端粒结合,在维持染色体稳定性中具有重要作用[26-27]。
已有较多研究表明,MYB转录因子参与调控植物黄酮类次生代谢产物合成,PpMYB10.1是红皮桃(Prunus persica)花青素积累的主要调节因子,并激活PpUFGT转录[28];拟南芥AtMYB12和葡萄(Vitis vinifera)VvMYBF1被鉴定为黄酮醇特异性激活剂,AtMYB12转化至转基因番茄(Solanum lycopersicum)中可以产生大量的黄酮醇[29-31]。黄芩中黄酮类成分的转录调控研究仍比较薄弱,Yuan等[13-14]研究人员前期从黄芩cDNA中筛选出19个SbMYB,并通过转基因烟草实验证明了SbMYB2、SbMYB7和SbMYB8可影响转基因烟草中黄酮类化合物相关基因(NtPAL1、NtPAL2、NtC4H和NtUFGT)的转录水平从而影响黄酮类化合物的积累。目前黄芩中黄芩素、野黄芩素、汉黄芩素及其糖苷类黄酮成分的生物合成途径及关键酶已经解析,本研究通过Cor函数计算SbMYB与黄酮类化合物生物合成结构基因间的相关性,并预测它们之间潜在的PPI关系,构建了基因调控网络,从而预测到SbMYB转录因子与黄酮类化合物合成通路上结构基因间潜在的调控关系。可能参与黄酮类化合物生物合成的43个SbMYB转录因子中,38个SbMYB可能参与正调控,9个SbMYB可能参与负调控。
通过调节转录因子对代谢途径上关键酶活性的调控作用,从整体角度调控代谢途径以获得高产量目标产物具有重要的科学价值。然而转录因子如何调控植物次级代谢产物的合成与积累是一个复杂的研究体系,如苹果(Malusdomestica)中MdMYB308L可以与MdbHLH33相互作用,增强其与MdCBF2和MdDFR启动子结合,正向调控苹果花青素的积累,苹果的泛素连接酶(MdMIEL1)还可以与MdMYB308L互作,促进MdMYB308L降解从而负调控花青素积累[32],因此,揭示其复杂调控网络与调控模式是未来的一项重要任务。本研究为黄芩中参与黄酮类成分生物合成调控的MYB转录因子筛选提供重要参考,在后续研究中,要结合先进的分子生物学技术,强化对重要SbMYB转录因子的机制解析。
