薯蓣皂苷对血管紧张素Ⅱ诱导的血管重构的影响及其机制
2022-08-13王建平周青山
王建平 周青山
武汉大学人民医院重症医学科 湖北 武汉 430060
高血压主要表现为动脉血压持续升高,是心肌梗死、冠状动脉粥样硬化性心脏病、脑卒中、心力衰竭等多种疾病的独立危险因素,其病程进展导致血管重构,引起心、脑、肾、视网膜等多脏器靶器官损伤[1,2]。高血压诱导的血管重构病理生理学改变主要有:血管内膜和中层增厚,外膜纤维化,血管张力和硬度增加;内皮细胞功能异常,血管平滑肌细胞增殖和迁移,血管炎症激活;细胞外基质稳态失衡[3⁃5]。改善高血压诱导的血管重构,可能为预防和治疗高血压及其并发症提供理论基础。
薯蓣皂苷是从中草药植物中提取,具有抗肿瘤、抗炎、抗心肌肥厚、护肾、护肝等广泛保护效应[6,7]。既往大量研究证实:薯蓣皂苷抑制多种恶性肿瘤细胞生长,缓解肝脏、肾脏和脑等组织器官缺血/再灌注损伤[8⁃10]。在心血管疾病方面,薯蓣皂苷可改善阿霉素导致的心脏毒性损伤,改善氧化应激诱导的心肌细胞凋亡[11]。而薯蓣皂苷在高血压诱导的血管重构中是否有保护作用尚不清楚。本研究将通过动物实验,阐明薯蓣皂苷在血管紧张素诱导的血管重构中作用及机制。
1 材料与方法
1.1 实验动物将40 只雄性C57BL/6 小鼠(清洁级,12 周龄左右)(购自湖北省实验动物中心)分为4组:生理盐水灌胃组(Saline 组)、薯蓣皂苷灌胃组(Dioscin 组)、血管紧张素Ⅱ泵注+生理盐水灌胃组(AngⅡ+Saline 组)、血管紧张素Ⅱ泵注+薯蓣皂苷灌胃组(AngⅡ+Dioscin),每组10 只小鼠。生理盐水或薯蓣皂苷[80 mg/(kg·d)](购自美国Selleck 公司)连续灌胃7 d,再予以血管紧张素Ⅱ(购自美国Sigma 公司)[1 500 ng/(kg·d)]皮下泵注14 d。处死前,利用全自动大小鼠无创血压测量仪(购自澳大利亚PowerLab 公司),测量小鼠清醒静息状态下尾动脉收缩压(mmHg),取3 次有效测量数据取平均值。2%戊巴比妥钠麻醉后处死小鼠,取胸主动脉、心脏、血浆、肾脏,液氮处理后,-80 ℃冰箱保存。
1.2 苏木素⁃伊红(HE)染色取小鼠胸主动脉后,4%多聚甲醛固定24 h,乙醇梯度脱水,二甲苯透明,浸蜡后包埋和切片。石蜡切片二甲苯脱蜡,乙醇梯度水化,透明。苏木紫染色10 min,洗涤后盐酸乙醇分化,自来水冲洗15 min,伊红染色3 min。脱水、透明、中性树脂封片,显微镜观察胸主动脉大体形态。
1.3 Van Gieson 染色将石蜡切片脱蜡、水化,铁苏木紫染色10 min,洗涤后盐酸乙醇分化。加入Van Gieson 染液染色5 min,95%乙醇分化,脱水、透明,封片。显微镜观察胸主动脉弹力纤维状态。
1.4 组织免疫组化将小鼠胸主动脉石蜡包埋组织切片,二甲苯脱蜡,乙醇梯度脱水,切片放入枸橼酸⁃枸橼酸钠溶液(pH=7.4)微波炉加热,进行抗原修复,PBS 洗3 遍;3% 过氧化氢溶液孵育10~15 min,阻断组织内源性过氧化物,PBS 洗3 遍;5%山羊血清室温封闭1 h,PBS 洗3 遍;加入相应一抗(PCNA,稀释浓度1∶200;ICAM⁃1,稀释浓度1∶150),4 ℃封闭过夜,PBS 洗3 遍;辣根过氧化物酶标记的二抗室温下孵育1 h,二氨基苯胺(DAB)避光显色10 min。苏木紫复染细胞核,乙醇梯度脱水,二甲苯透明处理,中性树脂封片,普通显微镜拍摄,采用Image⁃pro Plus 软件分析图片,计算并统计PCNA 和ICAM⁃1 相对表达水平。
1.5 组织免疫荧光小鼠胸主动脉石蜡包埋组织切片,二甲苯脱蜡,乙醇梯度脱水,切片放入枸橼酸⁃枸橼酸钠溶液(pH=7.4)微波炉加热,进行抗原修复,PBS 洗3 遍;3%过氧化氢溶液孵育10~15 min,阻断组织内源性过氧化物,PBS 洗3 遍;5%山羊血清室温封闭1 h,PBS 洗3 遍;加入相应一抗(α⁃SMA,稀释浓度1∶200;VCAM⁃1,稀释浓度1∶150),4 ℃封闭过夜,PBS 洗3 遍;滴加藻红蛋白(PE)标记的荧光二抗,室温避光孵育1 h,PBS 洗3遍;二脒基⁃2⁃苯基吲哚二盐酸盐(DAPI)复染细胞核,脱水、透明、封片,倒置荧光显微镜观察,Image⁃pro Plus 软件分析,计算并统计主动脉组织VCAM⁃1 和α⁃SMA 相对表达水平。
1.6 免疫印迹实验全组织蛋白提取试剂盒提取组织蛋白,BCA 法测定蛋白浓度,上样后10%SDS⁃PAGE 胶电泳(80 V,0.5 h;110 V,1 h),将蛋白电泳转膜至PVDF 膜,5% BSA 封闭2 h,TBST 洗膜3遍;相应一抗(稀释浓度1∶1 000)4 ℃孵育过夜。TBST 洗3 次后,辣根过氧化物酶标记的二抗室温孵育1.5 h,TBST 洗3 遍,增强型化学发光试剂曝光,Tanon 全自动化学发光图像分析系统获取蛋白条 带,以GAPDH 为 内 参,Gel⁃Pro Analyzer 4.0 软件分析目的条带灰度值。
1.7 细胞实验培养小鼠主动脉血管平滑肌细胞(MOVAS),细胞融合度达到50%,加入生理盐水或薯蓣皂苷(200 ng/mL),孵育2 h。加入AngⅡ(1 μmol/L),继续孵育24 h。提取细胞蛋白,进行Western Blot 检测。
1.8 统计学分析采用Graphpad Prsim5.0 进行统计,数据以均值±标准误表示,组间差异采用单因素方差分析(ANOVA),以P<0.05 为差异有统计学意义。
2 结果
2.1 薯蓣皂苷缓解血管紧张素Ⅱ诱导的血管重构见图1。血管紧张素Ⅱ皮下泵注2 周成功诱导小鼠高血压形成。与saline 组相比,AngⅡ+Saline组小鼠血压显著升高[分别为(154±6.01)mmHg和(113.36±6.68)mmHg,P<0.05)]。薯蓣皂苷灌 胃 干 预 后,与Ang Ⅱ+Saline 组 相 比,Ang Ⅱ+Disocin 组小鼠血压下降[分别为(136.02±8.9)mmHg 和(154±6.01)mmHg,P<0.05)](图1C)。形态学检测发现:血管紧张素Ⅱ皮下泵注成功诱导血管重构,AngⅡ+Saline 组小鼠胸主动脉HE 染色可见血管中层增厚,EVG 染色可见部分弹力纤维断裂(图1A 和1B)。与AngⅡ+Saline 组相比,AngⅡ+Dioscin 组小鼠胸主动脉血管重构明显缓解,血管中层厚度降低(P<0.05),弹力纤维走行规整,未见断裂(图1A 和1B)。
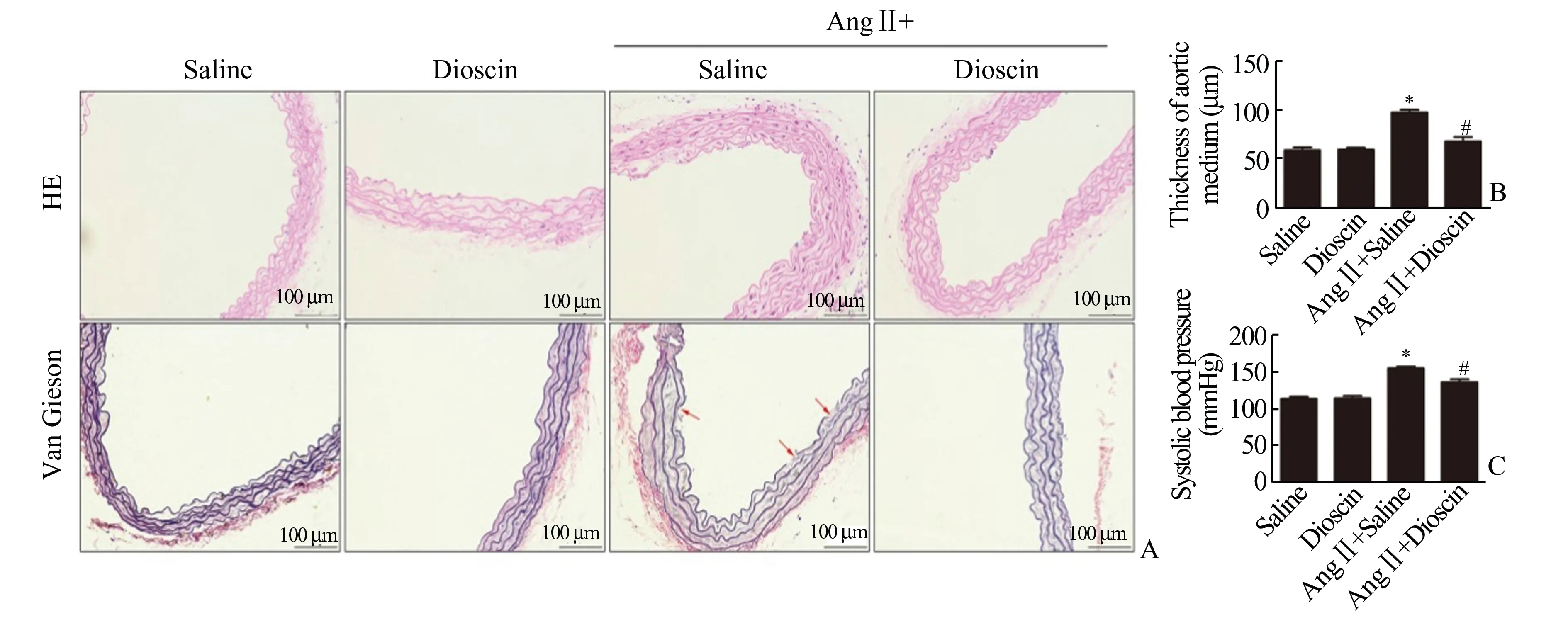
图1 薯蓣皂苷缓解血管紧张素Ⅱ诱导的血管重构
2.2 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管内皮细胞功能异常免疫组化和组织免疫荧光染色结果表明,与Saline 组相比,AngⅡ+Saline 组小鼠主动脉组织ICAM⁃1 和VCAM⁃1 表达上调(P<0.05)(图2),这提示血管紧张素Ⅱ刺激后,血管内皮细胞功 能 异 常。与Ang Ⅱ+Saline 组 相 比,Ang Ⅱ+Dioscin 组小鼠主动脉组织ICAM⁃1 和VCAM⁃1 表达下调(P<0.05)(图2)。

图2 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管内皮细胞功能异常
2.3 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管平滑肌细胞表型转化通过免疫组化和组织免疫荧光实验,我们发现:与Saline 组相比,AngⅡ+Saline 组小鼠主动脉组织α⁃SMA 表达水平显著下降(P<0.05),PCNA 表达上调(P<0.05)。与AngⅡ+Sa⁃line 组相比,AngⅡ+Dioscin 组小鼠主动脉α⁃SMA表达上调(P<0.05),PCNA 表达下调(P<0.05)(图3A)。Western Blot 结果显示:与Saline 组相比,AngⅡ+saline 组小鼠主动脉发生显著的血管平滑肌细胞表型转化,收缩型标记分子α⁃SMA 和cal⁃ponin1 表达下调(P<0.05),合成型标记分子Vi⁃mentin 和PCNA 表达上调(P<0.05)。与AngⅡ+Saline 组相比,AngⅡ+Dioscin 组主动脉血管平滑肌细胞表型转化缓解,α⁃SMA 和calponin1 表达水平上调(P<0.05)(图3B、3C、3D),Vimentin 和PCNA表达水平下降(P<0.05)(图3B、3E、3F)。
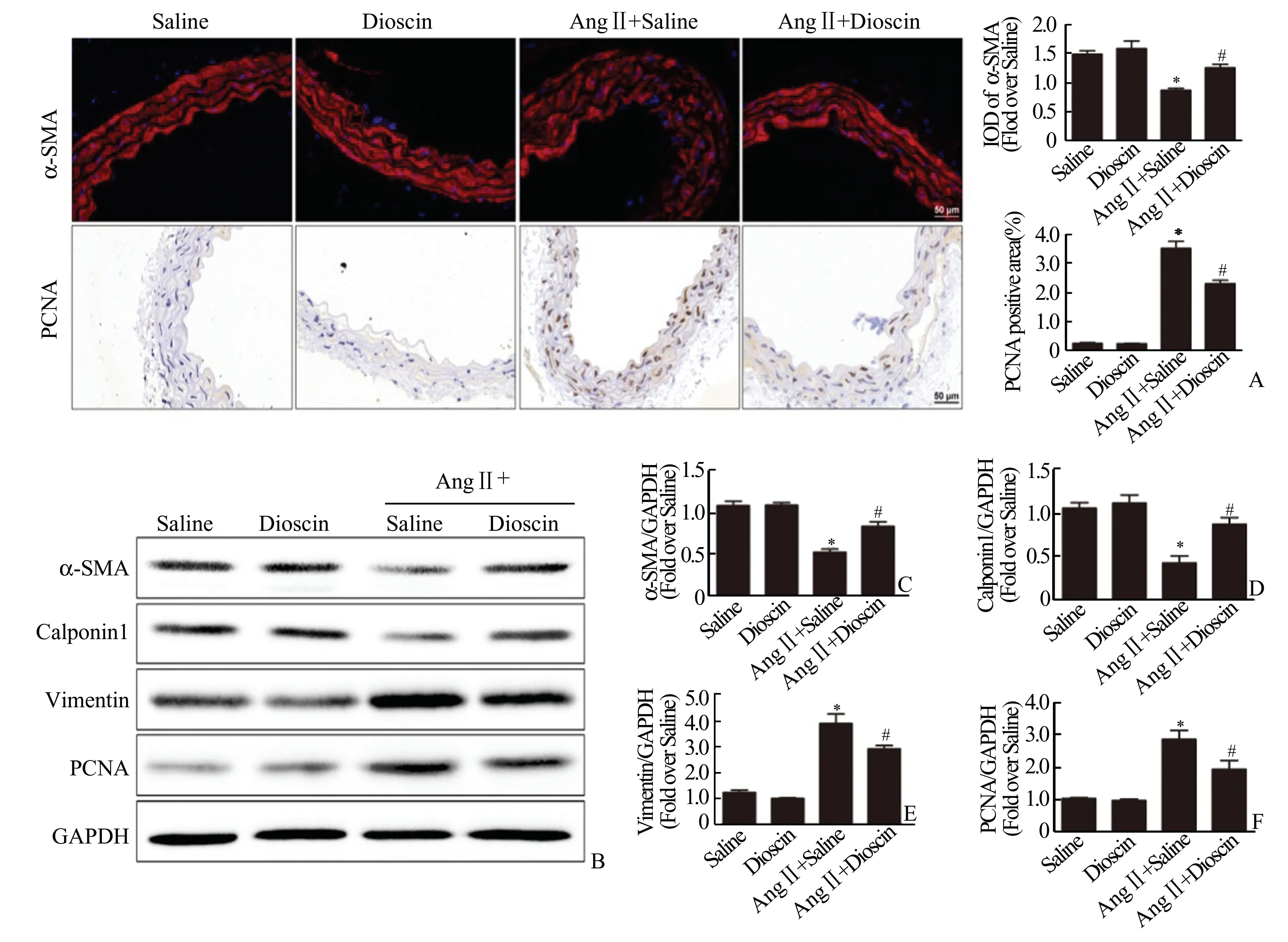
图3 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管平滑肌细胞表型转化
2.4 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管炎症通路激活Western Blot显示:与Saline 组相比,AngⅡ+Saline 组小鼠主动脉组织磷酸化STAT1 和磷酸化IκBα 表达显著上调(P<0.05),提示血管重构过程中炎症通路激活。与AngⅡ+Saline 组相比,AngⅡ+Dioscin 组小鼠主动脉组织磷酸化STAT1 和磷酸化IκBα 表达下调(P<0.05)(图4)。

图4 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管炎症通路激活
2.5 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管平滑肌细胞表型转化和炎症通路激活 细胞实验Western Blot 显示:与Saline 组相比,AngⅡ+Saline组血管平滑肌细胞收缩型标记分子α⁃SMA 表达下调(P<0.05),合成型标记分子Vimentin 表达上调(P<0.05),提示血管紧张素诱导血管平滑肌细胞发生表型转化;炎症通路激活相关分子磷酸化STAT1 和磷酸化IκBα 表达显著上调(P<0.05)。与AngⅡ+Saline 组 相 比,AngⅡ+Dioscin 组 小 鼠血管平滑肌细胞表型转化缓解,磷酸化STAT1 和磷酸化IκBα 表达下调(P<0.05)(图5)。
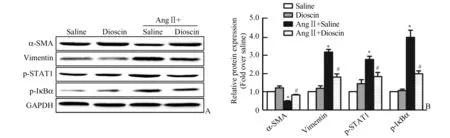
图5 薯蓣皂苷抑制血管紧张素Ⅱ诱导的血管平滑肌细胞表型转化和炎症通路激活
3 讨论
高血压是由基因、环境因素、神经⁃体液因子失衡、肾素⁃醛固酮系统过度激活、胰岛素抵抗等众多因素导致的全身阻力血管压力持续升高(收缩压≥140 mmHg 或舒张压≥90 mmHg)。长期高血压导致血管重构,血管重构主要病理生理学改变包括:血管内皮功能障碍,血管平滑肌细胞增殖、迁移和表型转化增强,血管外膜纤维化,血管炎症激活,胶原沉积,血管壁增厚,动脉硬化,收缩/舒张功能异常,引起血管功能和结构异常,最终导致血流动力学紊乱,心脏、肾脏、视网膜、脑等多脏器靶器官受损[4,12]。控制血压和改善血管重构是高血压及相应并发症治疗的重要措施。
血管内皮细胞功能异常是血管重构发生发展的重要环节。血管内皮细胞合成释放多种细胞因子,参与血管张力维持、血管新生、炎症反应、血管通透性等多种病理生理学效应的调节[13,14]。高血压中,血管内皮细胞舒血管因子如内皮细胞来源超极化因子(EDH)、内皮源性一氧化氮等合成减少,缩血管因子如内皮素⁃1、血栓素A2 和超氧化物等释放增加,血管持续收缩[15]。此外,高血压发生发展过程中,血管内皮细胞氧化应激和炎症反应激活,内皮细胞表面的细胞间黏附因子(血管细胞黏附分子⁃1、细胞间黏附分子⁃1、内皮细胞选择素等)表达上调,内皮细胞分布和排列完整性破坏,血管通透性增加,易于炎症细胞吸附、募集和迁移至血管壁[13,16]。薯蓣皂苷抑制TNF⁃α 诱导的血管内皮细胞VCAM⁃1、ICAM⁃1 和内皮脂肪酶的表达[17]。本研究发现,薯蓣皂苷干预可缓解血管紧张素Ⅱ诱导的主动脉组织VCAM⁃1 和ICAM⁃1 表达上调,改善内皮细胞功能。
血管平滑肌细胞表型转化在血管重构中起重要作用。血管紧张素Ⅱ诱导的高血压发生发展过程中,血管平滑肌细胞增殖和迁移能力增强,同时发生显著的表型转化,由收缩表型向合成表型转化,收缩表型标记分子α⁃SMA、calponin1、smoothe⁃lin 和desmin 等表达下调,合成表型标记分子Vimentin、OPN 和PCNA 等表达上调[4,18]。既往研究发现:薯蓣皂苷可通过抑制MAPK⁃FoxM1 通路,缓解颈动脉损伤后内膜增生[19]。我们动物实验和细胞实验研究发现:薯蓣皂苷可改善血管紧张素Ⅱ诱导的血管平滑肌细胞表型转化。
血管炎症反应激活在血管重构中有着重要作用。血管重构中,血管内皮细胞功能异常,血管通透性增强,炎症细胞吸附和迁移进入血管壁,刺激血管内皮细胞、血管平滑肌细胞和血管外膜成纤维细胞炎症反应激活,释放大量炎症因子,进一步加重内皮损伤、血管平滑肌细胞增殖迁移和表型转化、外膜纤维化、炎症细胞浸润等病理生理学改变[16,20]。大剂量血管紧张素Ⅱ干预甚至诱导动脉瘤/主动脉夹层形成[21]。既往研究发现:薯蓣皂苷抑制血管内皮细胞和巨噬细胞TNF⁃α、IL⁃1β 和IL⁃6 分泌;同时,薯蓣皂苷抑制TLR4 通路,降低NF⁃κB 转录活性,改善脑缺血再灌注损伤[7]。我们研究发现:薯蓣皂苷干预可降低血管紧张素Ⅱ诱导的主动脉组织磷酸化STAT1 和磷酸化IκBα 表达水平,缓解主动脉组织炎症反应。薯蓣皂苷可能抑制血管内皮细胞炎症反应,改善内皮功能,抑制炎症细胞黏附;抑制血管平滑肌细胞炎症反应,减少炎症因子释放,缓解血管平滑肌细胞表型转化,维持血管弹性。薯蓣皂苷通过缓解血管内皮细胞和血管平滑肌细胞炎症激活和改善平滑肌细胞表型转化,改善血管紧张素Ⅱ诱导的血管重构。
综上所述,本研究发现并初步证实薯蓣皂苷改善血管内皮功能,抑制血管平滑肌表型转化和血管炎症通路激活,改善血管紧张素Ⅱ诱导的血管重构,为高血压及其并发症防治提供新的理论依据。本研究不足之处在于,薯蓣皂苷抗血管重构具体靶点尚不清楚,需进一步研究。
