β-谷甾醇靶向CDC25B抑制肝癌细胞增殖
2022-08-11王凯李卫李志芳纪会琼郭志伟曹俊秋
王凯,李卫,李志芳,纪会琼,郭志伟,曹俊秋
(1.达州中医药职业学院公共基础部,四川达州 635000;2.达州市疾病预防控制中心附属医院,四川达州 635000;3.南充市中心医院影像科,四川南充 637000;4.四川大学华西医院肝胆外科,四川成都 610000)
肝癌是发生在肝脏的恶性实体瘤,肝癌发病早期无典型症状,晚期可表现为发热、乏力和肝脏区域疼痛等症状,肝癌早期治愈几率较高,晚期治疗方案复杂且患者预后差异较大[1]。肝癌发病率位居恶性肿瘤第4位,致死率位居第2位,男性发病率高于女性,肝癌发病具体作用机制和诱发因素尚未阐明,普遍认为肝癌发病与饮酒、食用霉变食物、病毒性肝炎、寄生虫和遗传等因素有关[2],肝癌基因组学研究[3]显示肝癌发病或进展和转移与多种基因突变、表观遗传修饰有关,肝细胞癌变后会重新激活端粒酶,导致癌变细胞增殖失控。细胞分裂周期25B(cell division cyclin 25B,CDC25B)是调控细胞周期的蛋白酪氨酸磷酸酶家族成员CDC25 的异构体之一,在细胞周期的特定阶段发挥作用[4],有研究[5-6]显示CDC25B可参与乳腺癌、黑色素瘤等疾病的调节过程。多种植物中的天然成分表现明显的抗肿瘤活性,其中包括紫杉醇、姜黄素、长春花素等,β-谷甾醇是广泛分布于植物中的甾醇类物质,我国杜仲、玄参和天冬等传统中草药的药用成分中也含有大量的植物甾醇类物质。已有研究表明[7]红豆杉中的β-谷甾醇和β-谷甾醇-葡萄糖苷对肝癌细胞表现毒性作用,但其具体作用机制尚不明确,故本研究通过肝癌数据库平台联合分子生物学技术探讨黄β-谷甾醇对肝癌细胞增殖的影响及其作用机制。
1 材料和方法
1.1 细胞和主要试剂
HepG2细胞系和Hep3B细胞系(中科院上海细胞库),6 周龄BALB∕C Nude 裸小鼠【成都达硕实验动物有限公司SCXK(川)2020-030】,动物实验遵守3R 原则并通过达州中医药职业学院实验动物伦理委员会批准(审批号20210309A);β-谷甾醇(货号CFN99916,纯度>95%,武汉中标科技有限公司),CCK-8试剂盒(货号CK04,深圳市纽邦生物技术有限公司),细胞凋亡和细胞周期试剂盒(货号A025、F10797,艾美捷科技有限公司),CDC25B 抗体(货号FNab01522,武汉菲恩生物科技有限公司),Ki67 抗体(货号AD80028,上海研卉生物科技有限公司),PVDF 膜(货号ISEQ00010,美国Milipore公司),蛋白提取试剂盒(货号SD-001,美国Invent Biotechnologies 公司),LipofectamineTM2000(货号11668019,美国Invitrogen公司)。
1.2 主要仪器
流式细胞仪(Aurora 3L-5L,美国Cytek Biosciences 公司),显微镜(X-Cite® 120PC Q,武汉提沃克科技有限公司),酶标仪(SpectraMax iD3,美国Molecular Devices 公司),冷冻离心机(Selectspin™17R,美国Select BioProducts 公司),细胞培养箱(Galaxy 48 R,德国Eppendorf公司)。
1.3 生物信息学分析
通过SwissTarget 和TargetNet 数据平台预测β-谷甾醇的靶点基因37 个,GEO2R 分析GSE101728肝癌数据集中表达上调基因并与37 个靶点的交集基因5 个,然后通过下载TCGA_LIHC 数据矩阵分析得到5 个基因中只有CDC25B 与肝癌TNM 分期(T1、T2-4)、临床分期(I 期、Ⅱ-Ⅳ期)、癌症组织分化(G1-2 分化、G3-4 分化)、复发情况(复发、未复发)和患者生存预后(预后良好、预后不良)存在关系。
1.4 细胞培养和β-谷甾醇处理
将HepG2 细胞系和Hep3B 细胞系置于DMEM培养基(含10%胎牛血清)中培养的96 孔板培养(每孔3×105个),培养条件为37℃、5% CO2加湿培养箱,孵育12 h 后,HepG2 细胞系给予β-谷甾醇(0、5、10、15、20µmol∕L)处理48 h,Hep3B 细胞系给予β-谷甾醇(0、6、12、18、24 µmol∕L)处理48 h,加入10µL CCK-8 溶液,继续孵育2 h,在波长450 nm处检测光密度(OD),计算细胞活力相对活力=(实验组OD∕空白组OD)×100%。根据两种细胞系的细胞相对活力情况,Hep3B 细胞系以15 µmol∕L 作为β-谷甾醇处理浓度,HepG2 细胞系以18µmol∕L 作为β-谷甾醇处理浓度,分别作用于0、1、2、3、4、5 d时使用CCK-8 法检测在450 nm 处OD,并设置不用β-谷甾醇处理的细胞作为Control 组,并绘制生长曲线。
1.5 细胞集落形成实验检测细胞增殖情况
将HepG2 细胞和Hep3B 细胞接种至6 孔板(每孔1 000 个),孵育12 h 后,Hep3B 细胞加入β-谷甾醇至终浓度为18µmol∕L,HepG2 细胞加入β-谷甾醇至终浓度为15µmol∕L,继续培养2 周,使用多聚甲醛固定15 min、1%结晶紫染色30 min,冲洗,统计细胞集落形成数目,计算相对集落形成率=(实验组集落形成数目∕对照组集落形成数目)×100%
1.6 流式细胞法检测细胞周期分布和细胞凋亡情况
收集Hep3B 细胞系(15µmol∕L β-谷甾醇处理48 h)和HepG2 细胞系(18 µmol∕L β-谷甾醇处理48 h),2 000g离心5 min 收集细胞沉淀。一部分细胞沉淀中经磷酸缓冲液洗涤2 次,75%乙醇-20℃固定24 h,0.4µL PI 避光孵育0.5 h,上样流式细胞仪检测处于不同细胞周期的细胞分布情况。一部分细胞沉淀中加入300µL 1×Binding Buffer 使细胞重悬,然后加入5 µL Annexin V∕FITC、10 µL PI 避光孵育15 min上样流式细胞仪检测细胞凋亡率。
1.7 裸鼠移植瘤模型实验
取18 只BALB∕C Nude 裸小鼠均分为Control 组和β-谷甾醇组,分别于皮下注射HepG2 细胞(8×106个细胞),同时β-谷甾醇组裸鼠腹腔注射β-谷甾醇50 mg∕kg·d,Control组裸鼠注射等体积DMSO,每2 d 测量1 次肿瘤的体积(1∕2×肿瘤长轴×肿瘤短轴2),连续14 d,处死裸鼠称量移植瘤质量和测量移植瘤体积,使用免疫组化检测移植瘤中Ki67 蛋白表达水平。
1.8 免疫组化
参考文献报道的方法[8]进行免疫组化检测蛋白表达,将移植瘤固定于40 g∕L 多聚甲醛溶液中并进行常规石蜡包埋、切片,枸橼酸抗原修复液微波修复10 min,磷酸缓冲液冲洗2 次,加入3%过氧化物酶阻断液封闭30 min,加入Ki67 抗体(1:500)37 ℃反应45 min,加入生物素标记的二抗继续孵育30 min,磷酸盐缓冲液冲洗×2 次,DAB 显色5 min,蒸馏水冲洗,苏木素染色20 s,乙醇浓度梯度脱水、二甲苯透明,中性树胶封片,光学显微镜(×200)下观察Ki67 表达水平,呈棕褐色颗粒即为Ki67 表达阳性。
1.9 Western blot检测蛋白表达
提取细胞总蛋白,调整蛋白浓度一致,经SDSPAGE 凝胶电泳、电转膜至PVDF 膜,5%脱脂奶粉密封2 h,加入兔抗人CDC25B、β-actin 一抗(1:500)4 ℃孵育过夜,TBST 漂洗,加入HRP 标记的二抗(1:500)孵育1 h,TBST 漂洗,显色、定影,使用凝胶成像仪自带系统分析蛋白条带相对灰度值。
1.10 细胞转染
参照LipofectamineTM2000 说明书进行细胞转染,细胞转染前24 h 将细胞提前接种于24 孔板,当细胞生长至融合30%~50%时,将培养基更换为无血清的培养基,加入CDC25B 过表达质粒和空质粒,同时每孔细胞中加入200µL 转染液继续孵育6 h,然后将无血清培养基更换为含有血清的培养基,继续培养48 h 后,用G418 筛选出稳定过表达CDC25B 的HepG2 和Hep3B 细胞系。使用Western blot验证细胞中CDC25B蛋白表达水平。β-谷甾醇处理稳定过表达CDC25B 的HepG2 和Hep3B 细胞(作用浓度同1.5;记为β-谷甾醇+CDC25B 组),空载对照组且进行β-谷甾醇处理的细胞记为β-谷甾醇组,不做处理的细胞记为Control 组,使用CCK-8、细胞集落形成实验、流式细胞法检测细胞活力、细胞增殖、细胞周期分布和细胞凋亡情况。
1.11 统计学处理
实验数据运用SPSS 22.0软件、Graph pad 5.0软件,数据以()表示;二组计量资料的均数比较,如果每一组资料都呈正态分布并且方差齐性,组间比较采用t检验,反之用校正t检验或秩和检验;多组间计量资料的均数比较,如果每一组资料都呈正态分布并且方差齐性,组间比较采用方差分析,方差分析差异有统计学意义时采用Bonferroni 法进行两两比较;二组生存分析使用Log-rankt检验。α=0.05。
2 结果
2.1 CDC25B与β-谷甾醇和肝癌临床关系分析
SwissTarget 和TargetNet 预测β-谷甾醇的靶点,共有37 个靶点。37 个靶点与GEO2R 分析GSE101728 肝癌数据集所得上调基因的交集基因有5个,其中包括CDC25B基因(图1A)。TCGA_LIHC 表达数据和临床信息比对发现只有CDC25B 的表达水平与肝癌患者总生存期相关,因此选择CDC25B 进一步研究。TCGA_LIHC 表达数据和临床对应信息分析显示CDC25B 在肝癌组织表达水平显著高于其癌旁组织(t=90.414,P<0.001;图1B),肝癌不同TNM 分期(T2-4)的CDC25B 表达水平显著高于TNM 分期(T1)(F=3635.158,P<0.001;P<0.001;图1C),肝癌临床分期(Ⅱ-Ⅳ)的CDC25B表达水平显著高于临床分期(I)(F=3136.663,P<0.001;P<0.001;图1D),肝癌G3-4 患者的CDC25B表达水平显著高于G1-2 患者(t=38.728,P<0.001;图1E),肝癌复发患者的CDC25B 表达水平显著高于未复发肝癌患者(t=29.710,P<0.001;图1F),预后不良肝癌患者的CDC25B 表达水平显著高于预后良好肝癌患者(t=27.006,P<0.001;图1G),以CDC25B 表达水平的中位数将患者分为CDC25B 低表达组和CDC25B 高表达组,CDC25B 高表达组肝癌患者的总生存率显著低于CDC25B 低表达组(Log-rankt=7.509,P=0.006;图1H)。

图1 CDC25B与β-谷甾醇和肝癌临床关系Fig.1 Clinical relationship among CDC25B,β-sitosterol and HCC
2.2 β-谷甾醇对肝癌细胞增殖和凋亡的影响
Hep3B 和HepG2 的细胞活力随着β-谷甾醇剂量增加而逐渐降低(F=108.400,P<0.001,图2A;F=95.180,P<0.001,图2B),Hep3B(β-谷甾醇15µmol∕L)和HepG2(β-谷甾醇18µmol∕L)的细胞生存率约为50%,因此选择此浓度做后续实验(图2A~D);Hep3B(β-谷甾醇15 µmol∕L)和HepG2(β-谷甾醇18µmol∕L)的相对克隆数目显著低于Control 组(t=16.515,P<0.001;t=22.517,P<0.001),S 期细胞百分比均显著低于Control 组(t=4.906,P=0.008;t=4.359,P=0.0125),G0∕G1期细胞百分比高于Control组(t=4.109,P=0.015;t=3.618,P=0.022),细胞凋亡率均高于Control 组(t=23.674,P<0.001;t=20.577,P<0.001;图2E~H)。

图2 β-谷甾醇对肝癌细胞增殖和凋亡的影响Fig.2 Effects of β-sitosterol on the proliferation and apoptosis of HCC cells
2.3 β-谷甾醇对体内移植瘤生长的影响
β -谷甾醇组移植瘤的体积显著低于Control 组(F=214.114,P<0.001,图 3A;t=2.240,P=0.040,图3B),移植瘤质量低于Control 组(t=2.475,P=0.025;图3C),Ki67 相对表达水平均显著低于Control 组(t=26.163,P<0.001;图3D)。
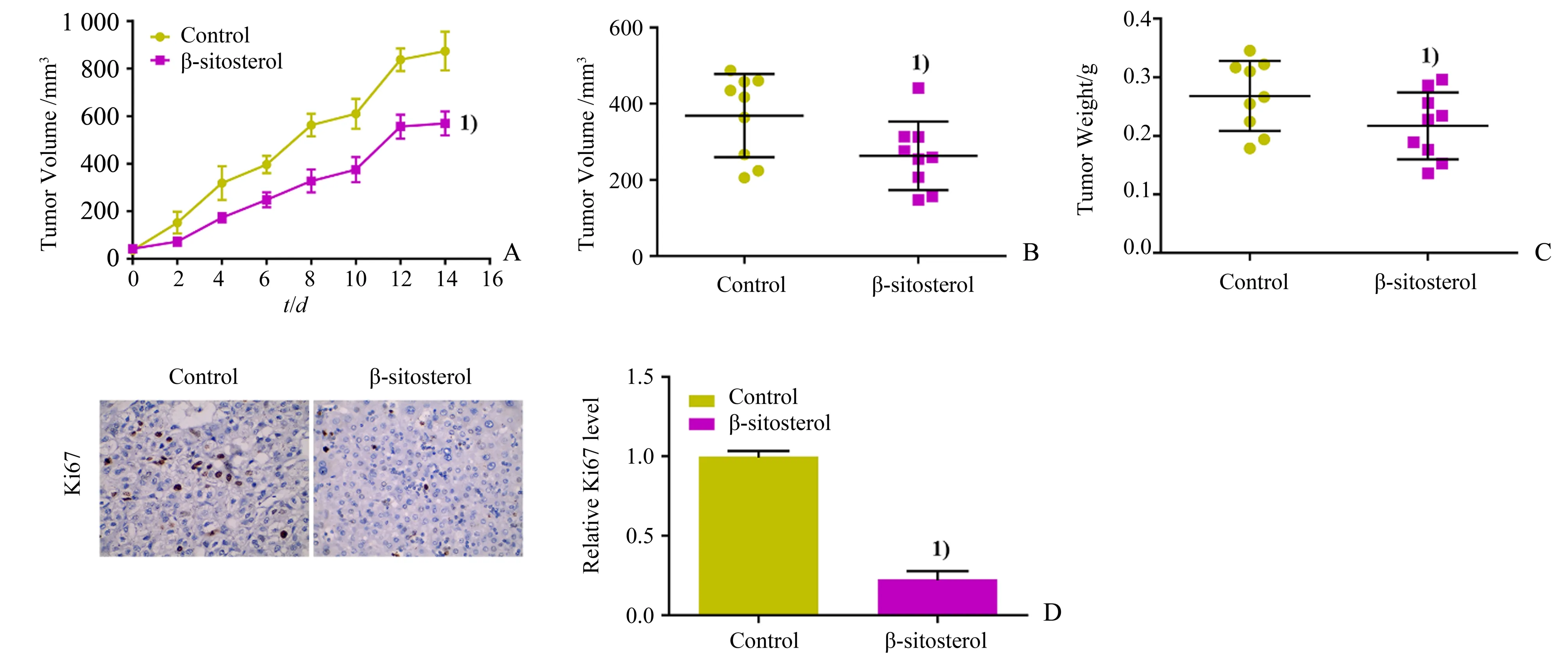
图3 β-谷甾醇对体内移植瘤生长的影响Fig.3 Effects of β-sitosterol on the growth of xenografted tumors in vivo
2.4 β-谷甾醇对肝癌细胞CDC25B 蛋白表达的影响
Hep3B 和HepG2 细胞中的CDC25B 蛋白水平随着β-谷甾醇浓度增加而逐渐降低,Hep3B(β-谷甾醇15µmol∕L)和HepG2(β-谷甾醇18µmol∕L)中的CDC25B 蛋白水平均随着β-谷甾醇作用时间延长而逐渐降低(图4)。

图4 β-谷甾醇对肝癌细胞CDC25B蛋白表达的影响Fig.4 Effects of β-sitosterol on the expression of CDC25B protein in HCC cells
2.5 β-谷甾醇通过CDC25B抑制肝癌细胞增殖
β-谷甾醇作用下Hep3B和HepG2细胞CDC25B蛋白表达降低(t=25.351,P<0.001;t=31.859,P<0.001;图5A),给予CDC25稳定过表达细胞系可以恢复细胞中CDC25蛋白表达(F=149.686,P<0.001;F=182.844,P<0.001;图5B)。Hep3B细胞的β-谷甾醇+CDC25B组的细胞生长、相对克隆数目、S期细胞百分比均显著高于β-谷甾醇组(F=176.247,P<0.001;F=168.153,P<0.001;F=25.052,P=0.001),而G0∕G1期细胞百分比、细胞凋亡率均显著低于β-谷甾醇组(F=15.490,P=0.004;F=222.843,P<0.001)(图5C、E、F、H)。HepG2 细胞的β-谷甾醇+CDC25B 组的细胞生长、相对克隆数目、S期细胞百分比均显著高于β-谷甾醇组(F=186.723,P<0.001;F=253.500,P<0.001;F=18.138,P=0.003),而G0∕G1 期细胞百分比、细胞凋亡率均显著低于β-谷甾醇组(F=17.138,P=0.003;F=413.382,P<0.001;图5D、E、G、H)。
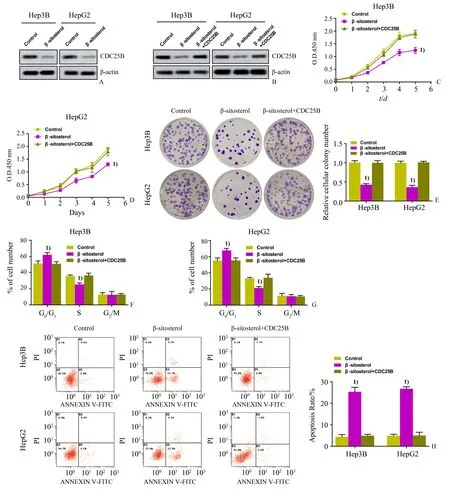
图5 β-谷甾醇通过CDC25B抑制肝癌细胞增殖Fig.5 β-Sitosterol inhibited the proliferation of HCC cells by CDC25B
3 讨论
肝癌的临床治疗方案确定主要依赖于患者的肝功能、肿瘤侵犯程度和转移情况,目前普遍认为手术切除治疗或肝移植治疗是可能根治肝癌的重要方式,但术后复发和手术切除率低仍旧是肝癌的治疗难点[9]。非手术治疗在控制肝癌病情、辅助术前和术后治疗具有重要意义,其中包括消融治疗、化疗、放射治疗和分子靶向治疗等[10]。β-谷甾醇是植物中广泛存在的多酚羟酸类化合物,同时也是构成真核生物细胞膜的主要成分之一,目前已经鉴定植物甾醇约200 种,β-谷甾醇在植物油、种子和果蔬中含量较高,具有抗氧化[11]、抗衰老[12]、降低胆固醇[13]和神经保护作用[14]。国外有研究[15]表明β-谷甾醇介导的银纳米粒子在人结肠癌HT-29 细胞中呈现出细胞毒性作用,可促进结肠癌细胞发生凋亡。本研究通过分析β-谷甾醇不同作用浓度和不同作用时间的Hep3B、HepG2 细胞活性和细胞增殖趋势发现β-谷甾醇可以显著抑制肝癌Hep3B 和HepG2细胞增殖,此结果在裸鼠移植瘤实验中也得到证实。
肿瘤增殖抑制受到多方面因素影响,其中包括细胞增殖和细胞凋亡[16-17],本研究结果显示β-谷甾醇作用下的Hep3B、HepG2 细胞相对克隆数目和S期细胞百分比显著降低,而细胞凋亡率和G0∕G1期细胞百分比显著升高,同时体内移植瘤中的Ki67表达水平显著降低,说明β-谷甾醇对肝癌细胞抑制作用可能是通过抑制癌细胞增殖和促进癌细胞凋亡实现。细胞增殖速率与细胞周期正常运转具有密切关系,一个完整的细胞周期包括有丝分裂和分裂间期,分裂间期包括G1 期、S 期和G2 期,任何一个时期出现问题均会影响细胞周期的正常运转[18]。G1 期主要是RNA 和蛋白质的合成,S 期是DNA 的复制[19],Ki67 是与核糖体RNA 转录有关的核蛋白,与细胞的有丝分裂具有密切关系,Ki67 常用于临床肿瘤恶性程度的判断,其表达水平越高表示细胞增殖越活跃[20]。本研究结果说明β-谷甾醇可阻碍使肝癌细胞周期运行,对细胞增殖有抑制作用,关于β-谷甾醇对肝癌细胞凋亡的促进作用可能与通过核因子相关因子2、氧化应激激活人肝癌细胞线粒体凋亡有关[21]。
本研究通过数据库分析显示CDC25B 是β-谷甾醇靶点之一,还发现CDC25B 在肝癌组织、高临床分期、复发和不良预后患者中呈高表达,并且还会影响患者的生存时间。CDC25B 为半衰期磷酸化蛋白,与细胞分裂有关的纺锤体前期的形成关系密切,是细胞早期有丝分裂的启动因子,目前普遍认为CDC25B 过量表达会促进肿瘤增殖和细胞恶性转化[22-23]。本研究结果显示β-谷甾醇可降低Hep3B、HepG2 细胞中CDC25B 表达水平,推测β-谷甾醇对肝癌增殖的抑制作用可能是通过降低CDC25B 表达实现,本研究通过构建CDC25B 稳定过表达细胞系并给予β-谷甾醇处理,发现CDC25B表达水平升高可以逆转β-谷甾醇对肝癌细胞增殖抑制作用和促进细胞凋亡作用,说明β-谷甾醇对肝癌细胞的抑癌作用可能与抑制CDC25B 表达有关。
综上所述,β-谷甾醇对肝癌细胞的凋亡有促进作用,从体外细胞实验和体内移植瘤实验证实β-谷甾醇可抑制肝癌细胞的增殖,其作用机制可能是通过抑制CDC25B 蛋白表达使细胞周期运转发生阻碍,从而抑制肝癌细胞增殖,但β-谷甾醇下调CDC25B 表达的具体机制尚需进一步探讨。本研究主要发现β-谷甾醇抑制肝癌细胞增殖的可能作用机制,而关于其对肝癌细胞凋亡的作用机制尚不清楚。
