北方汉族31个X染色体短串联重复片段突变及遗传多态性研究
2022-08-11徐妍刘岩郝世诚张金佩刘革新陶玉婷王显众袁丽
徐妍,刘岩,郝世诚,张金佩,刘革新,陶玉婷,王显众,袁丽
(中国政法大学证据科学教育部重点实验室,北京 100088)
女性含有两条X 染色体,除连锁基因外,在减数分裂时可以发生同源重组,随机遗传给子代一条X 染色体,与常染色体类似。男性体内只有一条X染色体,且除拟常染色区外,X 染色体与Y 染色体缺乏同源重组[1],因此减数分裂时男性个体的XSTR 以一种类似于Y-STR 的单倍型方式传递给其女儿。这种独特的遗传方式使X-STR 在含有女性的亲缘关系鉴定案件,如全同胞姐妹、同父异母半同胞姐妹、祖母和孙女、父女、母子或母女、三联体鉴定中[2-3]能够发挥独特的应用价值,并且可以利用X-STR 的基因频率或单倍型频率计算亲缘关系的似然率。当然,当联合应用多个X-STR 检验时需关注基因座之间是否存在连锁不平衡,以及突变等问题[4]。在一些家系重建案件中,检验更多的X-STR 来获得更多的遗传信息[5],以推导出重建人员更多确切的基因分型,往往能获得非常好的效果。既往报道的X-STR 基因座数量有限,而在复杂亲缘关系鉴定时往往需要检测更多的XSTR,这些基础群体数据缺乏,亟需有群体遗传信息来支撑案件的分析。基于以上背景,本文对北方汉族群体和家系样本检测31 个X-STR 基因座,开展了X-STR 遗传多态性、连锁不平衡检验和突变情况的研究。
1 材料与方法
1.1 样本
在获取知情同意条件下采集209 个中国北方汉族家系样本,具体为:205 个母亲-女儿-父亲三联体,2 个母亲-女儿二联体,2 个父亲-女儿二联体。所有家系样本已经常染色体STR检验,亲权指数均超过10 000,支持有亲子关系。家系样本用于观察31 个X-STR 基因座在414 次减数分裂过程中的突变率。选择所有家系中母亲∕父亲样本,分别组成207个女性∕男性无关个体群体,总共无关个体为414 人。本研究所设计样本实验均经中国政法大学证据科学研究院伦理委员会批准[批准号No.2020003],所有研究对象均签署了知情同意书。
1.2 X-STR分型检测
用Chelex-100 法提取基因组DNA。分别按照AGCU X19 STR 试剂盒和MicroreaderTM19X Direct ID System试剂盒说明书扩增各样本DNA。扩增产物经Applied Biosystems 3130 遗传分析仪检测、Gene-Mapper®ID-X v1.4软件分析,获得31个X-STR基因座的基因型,分别为DXS101、DXS981、DXS6789、DXS6795、DXS6800、DXS6803、DXS6807、DXS6809、DXS6810、DXS7132、DXS7133、DXS7423、DXS7424、DXS8378、DXS9902、DXS9907、DXS10074、DXS10075、DXS10079、DXS10101、DXS10103、DXS10134、DXS10135、DXS10148、DXS10159、DXS10162、DXS10164、GATA165B12、GATA172D05、GATA31E08和HPRTB。
1.3 统计学分析
对于209 个家系的X-STR 分型结果,依据遗传规律观察是否存在突变的基因座,并计算发生突变的基因座突变率。仅针对207 例女性X-STR基因座结果,使用Arlequin v3.5 软件进行31 个XSTR 遗传标记的Hardy-Weinberg 平衡检验及连锁不平衡检验。使用Arlequin v3.5 软件对男、女群体等位基因频率分布进行显著性差异分析。使用StatsX v2.0 软件统计各基因座的等位基因频率、观察杂合度(heterozygosity observed,Hobs)、期望杂合度(expected heterozygosity,Hexp)、基因差异度(gene diversity,GD)、女性个人识别率(discrimination power of female,DPF)、男性个人识别率(discrimination power of male,DPM)、多态性信息量(polymorphism information content,PIC)、平均父权排除概率(mean exclusion chance,MEC):父女或母子二联体X 染色体标记的平均父权排除概率(MEC_Desmarais_duo)、涉及女儿的三联体X 染色体标记的平均父权排除概率(MEC_Desmarais)。此外,将本研究的北方汉族群体分别与德国、日本、广东汉族、四川藏族群体[6-9]进行显著性差异分析。
2 结果
2.1 31个X-STR基因座的突变情况
根据遗传规律观察209个家系的X-STR 分型,经分析在19个基因座(DXS981、DXS10148、DXS7132、HPRTB、DXS9907、DXS10162、DXS7424、DXS10159、DXS10075、GATA172D05、DXS10135、DXS9902、DXS6810、DXS10164、DXS10101、DXS10079、DXS10103、DXS6789、GATA165B12)上发现29 个新发突变。总家系平均突变率为0.002 3。在这19 个X-STR 基因座上突变率最高的基因座 是DXS10135,为0.012 1;其次是HPRTB,为0.007 2;DXS7132、DXS10159、DXS981、DXS10162基因座突变率均为0.004 8(表1)。
2.2 31个X-STR基因座的遗传多态性
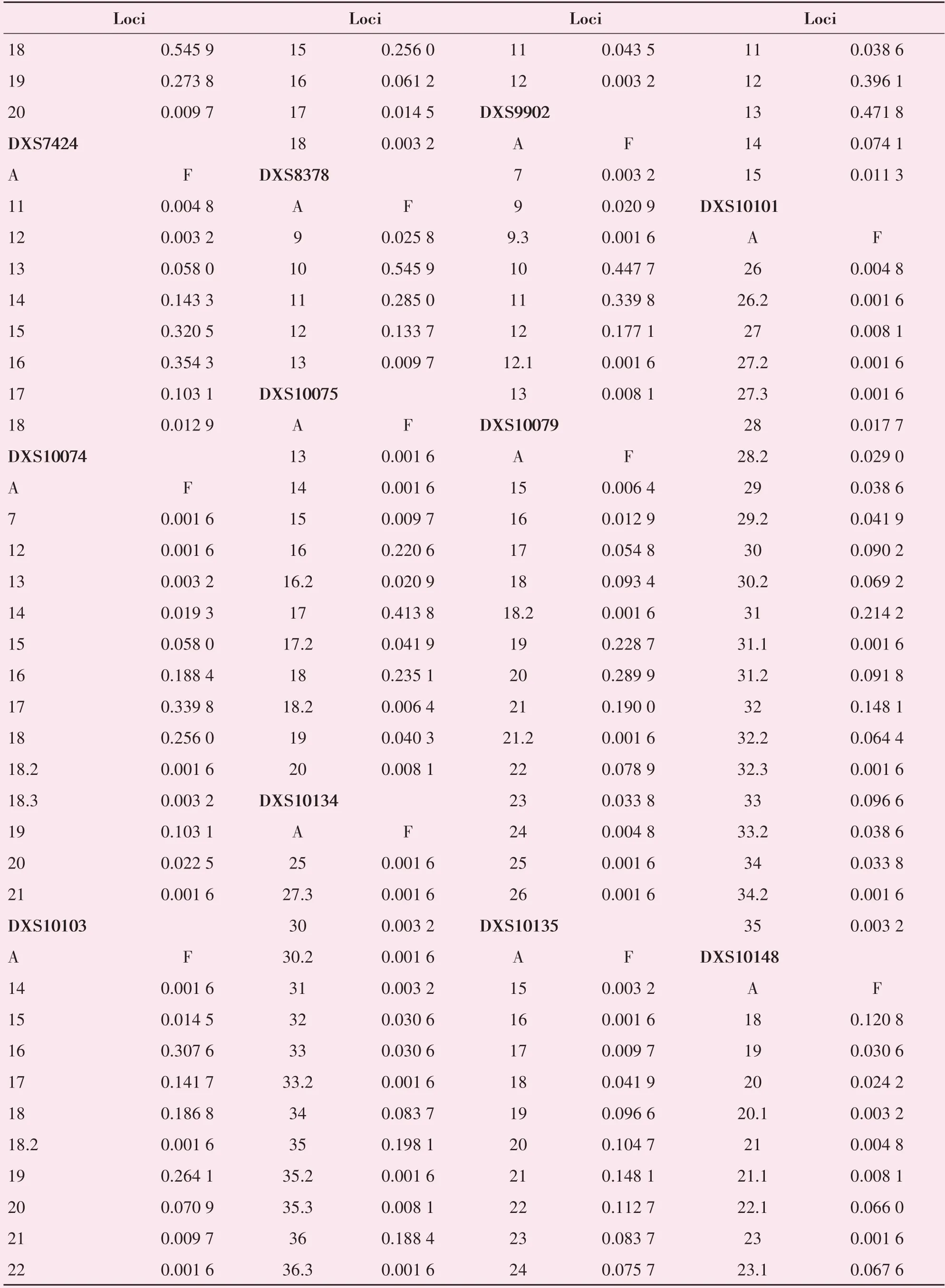
207 例女性样本的31 个X-STR 基因座基因型分布P 值均大于0.05,均符合Hardy-Weinberg 平衡。414 例无关个体的31 个X-STR 基因座共检测出了344 个不同的等位基因。对男、女群体等位基因分布频率进行显著性差异分析后的P值均大于0.05,表明31 个X-STR 等位基因频率分布在男、女两个群体中不存在显著差异,同时女性群体符合Hardy-Weinberg 平衡,故将女性、男性样本合并计算每个基因座的等位基因频率。表2 列出了合并后的基因座等位基因频率。等位基因的频率分布在0.001 6-0.810 0 之间,其中DXS6800 的等位基因16频率最高,为0.810 0。

表2 31个X-STR基因座在中国北方汉族群体中的等位基因频率Table 2 Allele frequencies of 31 X-STR loci in Northern Han of Chinese(n=414)
续表

续表
中国北方汉族群体31个X-STR基因座的法医学参数见表3。31 个X-STR 基因座的PIC 在0.312 7-0.912 4之间,DPM在0.330 5-0.918 1之间,DPF在0.534 0-0.987 6之间,MEC_Desmarais 在0.312 7-0.912 4,MEC_Desmarais_duo 在0.193 5-0.844 6之间。根据ChrX-STR.org 2.0数据库(http:∕∕www.chrx-str.org∕)中的公式,31个X-STR的女性群体累积个人识别概率为1-1.578 4×10-31,男性群体累积个人识别概率为1-4.566 4×10-19。累积的平均父权排除概率,三联体为1-2.281 9×10-17,二联体为1-2.509 0×10-12。

表3 31个X-STR基因座在中国北方汉族群体中的法医学参数Table 3 Forensic parameters of 31 X-STR loci in Northern Han of Chinese
2.3 31个X-STR基因座的连锁不平衡分析结果
在31 个X-STR 基因座配对组成的465 个配对基因座中,一共30 对基因座连锁不平衡检验P<0.05,见表4。经Bonferroni校正(0.05∕465=0.000107 526 9),仍有1对基因座(DXS10103-DXS10101)具有统计学意义,存在连锁不平衡。DXS10103-DXS10101 单倍型共有60 种,单倍型频率在0.004 8-0.087 0 之间(表5),其中单倍型16-31的频率最高,为0.087 0。

表4 连锁不平衡检验中P值小于0.05的基因座对Table 4 Locus pairs with P value less than 0.05 in linkage disequilibrium test
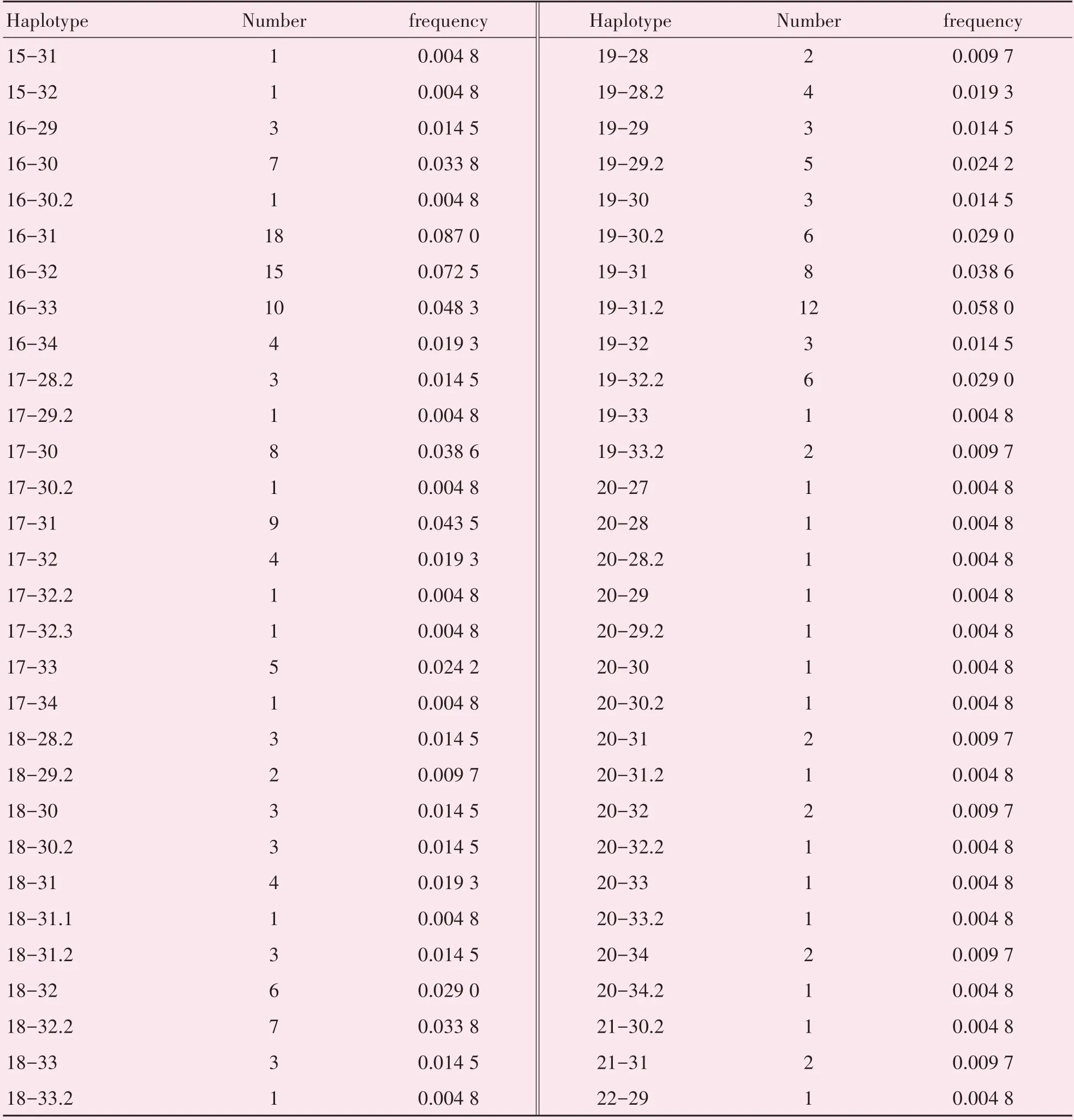
表5 DXS10103-DXS10101单倍型频率Table 5 Haplotype frequency of DXS10103-DXS10101
2.4 北方汉族群体与其他四个群体之间X-STR基因座等位基因频率分布的比较
本研究的北方汉族群体分别与德国、日本、广东汉族、四川藏族群体比较P值见表6。北方汉族群体与德国群体在可比较的13 个基因座上均存在明显差异,与日本群体在可提供数据比较的14 个基因座中有8 个基因座存在差异,与广东汉族群体在11个基因座中的3个基因座上存在差异、与四川藏族群体在可比较的19 个基因座中的6 个基因座上存在差异。
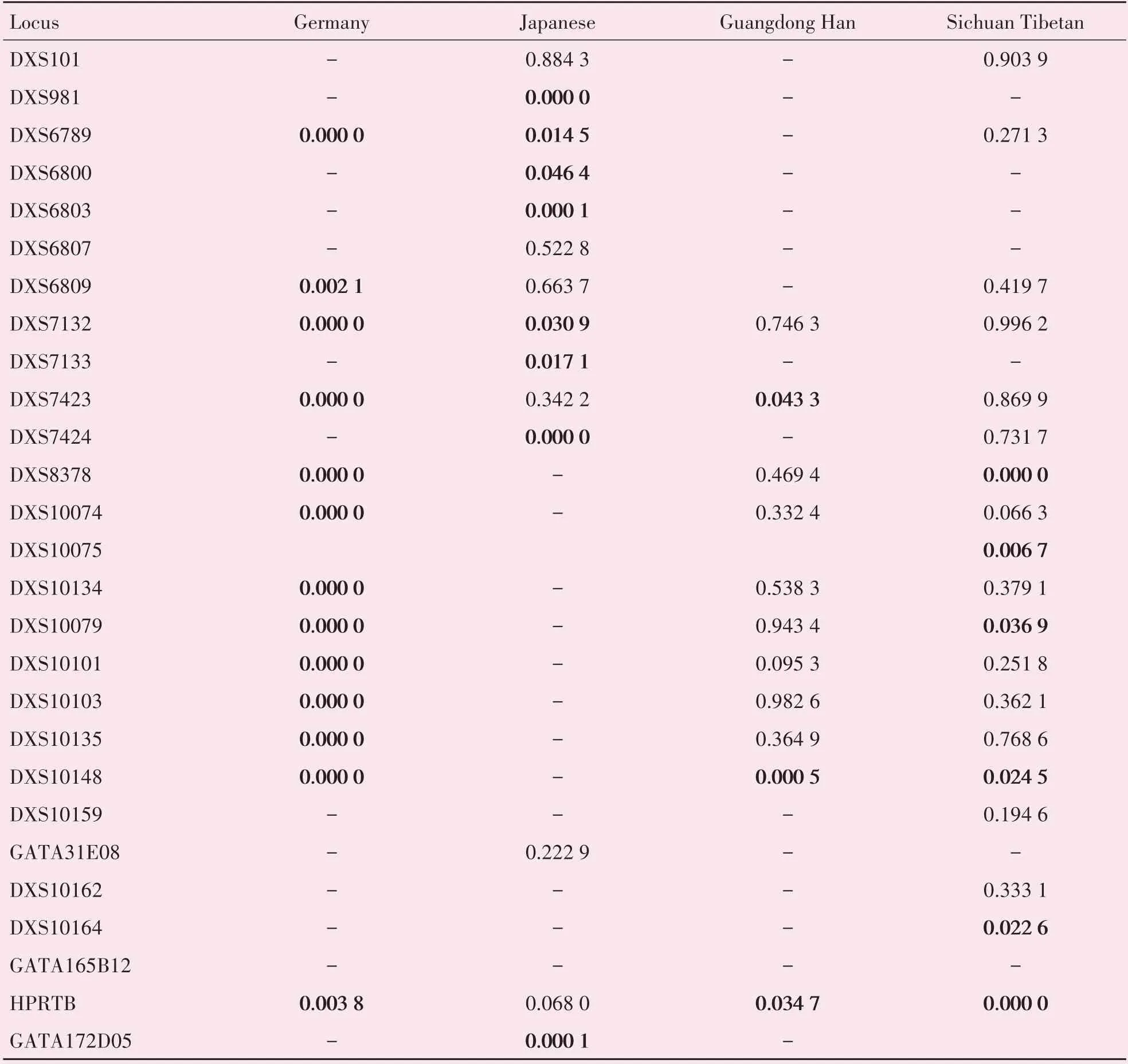
表6 北方汉族群体与其他四个群体之间X-STR基因座等位基因频率分布的差异检验结果Table 6 Difference test results of allele frequency distribution of X-STR loci between northern Han population and other four populations
3 讨论
经家系分析,31个X-STR在414次减数分裂中发生了29 个突变,且均为一步突变,未观察到有一个家系出现2 个突变。除4 个突变来源不明外,有22 个突变来源于父亲,有3 个突变来源于母亲,父源性突变与母源性突变的观察值比例约为7.3:1,表明了父源性突变相比较于母源性突变来说更容易发生。这种突变机率的差异可能来源于生殖细胞分化形成卵子和精子过程中细胞分裂次数的差异[10]。对于女性来说,分裂形成卵子的卵原细胞在胚胎发育结束时几乎已经确定,而对于男性来说,精原细胞能不断地进行有丝分裂,增加细胞数量并分化为精母细胞。因此,相比于卵子,精子的形成经历了更多的细胞分裂,这有可能是父源性突变率更高的原因之一。除突变来源外,突变率的大小还可能与遗传标记重复序列的结构特征、种群、样本量大小等因素有关[11-12]。本次研究的北方汉族人群中,31个X-STR 基因座平均突变率为0.002 3,与常染色体STR 和Y-STR 的平均突变率相近[13-16]。在北方汉族群体,DXS7132、HPRTB、DXS10159、DXS981、DXS10162 和DXS10135 基因座的突变率明显高于平均突变率。对31个X-STR 突变情况的研究,提示我们在检测X-STR 时,特别是DXS7132、HPRTB、DXS10159、DXS981、DXS10162 和DXS10135 这几个突变率较高的基因座,需要保持谨慎,不要因为个别基因座不符合遗传规律而排除具有某种亲缘关系,尤其是矛盾等位基因来自于父源。
在31 个X-STR 基因座中,共28 个基因座表现出了高度的多态性[17],即PIC>0.5,多态性较差的基因座有DXS7423、DXS7133、DXS6800。31 个XSTR 的女性群体累积个人识别概率为1-1.578 4×10-31,男性群体累积个人识别概率为1-4.566 4×10-19,可以作为常染色体STR 的有力补充来满足个人识别的需求。31 个X-STR 三联体累积的平均父权排除概率为1-2.281 9×10-17,二联体累积的平均父权排除概率为1-2.509 0×10-12,可以有效地辅助常染色体STR进行复杂亲缘关系的鉴定。
本次研究的北方汉族女性群体31个X-STR 基因座的基因型结果经连锁不平衡检验,在Bonferroni 校正(P<0.05∕465=0.000 107 526 9)后,仍有1 对基因座(DXS10103-DXS10101)具有显著性差异,存在连锁不平衡现象。因此,在案件涉及北方汉族的,统计分析中DXS10103-DXS10101按照单倍型频率来计算。同时,本次连锁不平衡检验也反映出,虽然在X 染色体上报道了4 个连锁群,但是实际检验出来的X-STR连锁不平衡并没有理论上的多,即使是位于同一个连锁群内,也有可能不存在连锁不平衡的情况,这在其他研究中也有类似的报道[18]。
不同群体之间的显著性差异研究可以发现,北方汉族群体与德国群体存在明显差异,与日本群体、四川藏族群体、广东汉族群体均存在一定程度上的差异。人种差异越大、地域分布越广泛,群体之间的差异也越大。这可能源于更为相近的地理距离或更为接近的种群在遗传上更具有同源性。因此在实践中应用X-STR时选择相对应的群体是至关重要的。
