基于环境因子的沙丁鱼太平洋群系亲体-补充量关系研究
2022-08-11章贤成汪金涛陈新军
唐 馨,章贤成,汪金涛,2,3,4,5,陈新军,2,3,4,5
(1.上海海洋大学海洋科学学院,上海 201306;2.农业农村部大洋渔业开发重点实验室,上海 201306;3.国家远洋渔业工程技术研究中心,上海 201306;4.大洋渔业资源可持续开发教育部重点实验室,上海 201306;5.农业农村部大洋渔业资源环境科学观测实验室,上海 201306)
沙丁鱼属鱼类(Sardina)为近海暖水性鱼类,通常栖息于中上层,适温在20~30℃左右,只有少数种类的适温较低,主要摄食浮游生物[1]。在太平洋群系中,1龄以上的沙丁鱼在黑潮周边越冬、产卵,夏秋季进行沿岸停留或小规模的向北方的索饵洄游[2](图1)。从卵的出现分布情况来看,沙丁鱼的产卵期为每年11月—次年6月,盛期为2—4月,沙丁鱼的最佳产卵温度在表层水温15℃到18℃之间[3]。近年来一些人类活动、大尺度气候和局部环境变化造成沙丁鱼资源补充量剧烈波动,亟需研究其亲体量与补充量关系及环境影响因素,了解资源量变化的动态过程,以期为其资源的可持续利用奠定基础[4]。
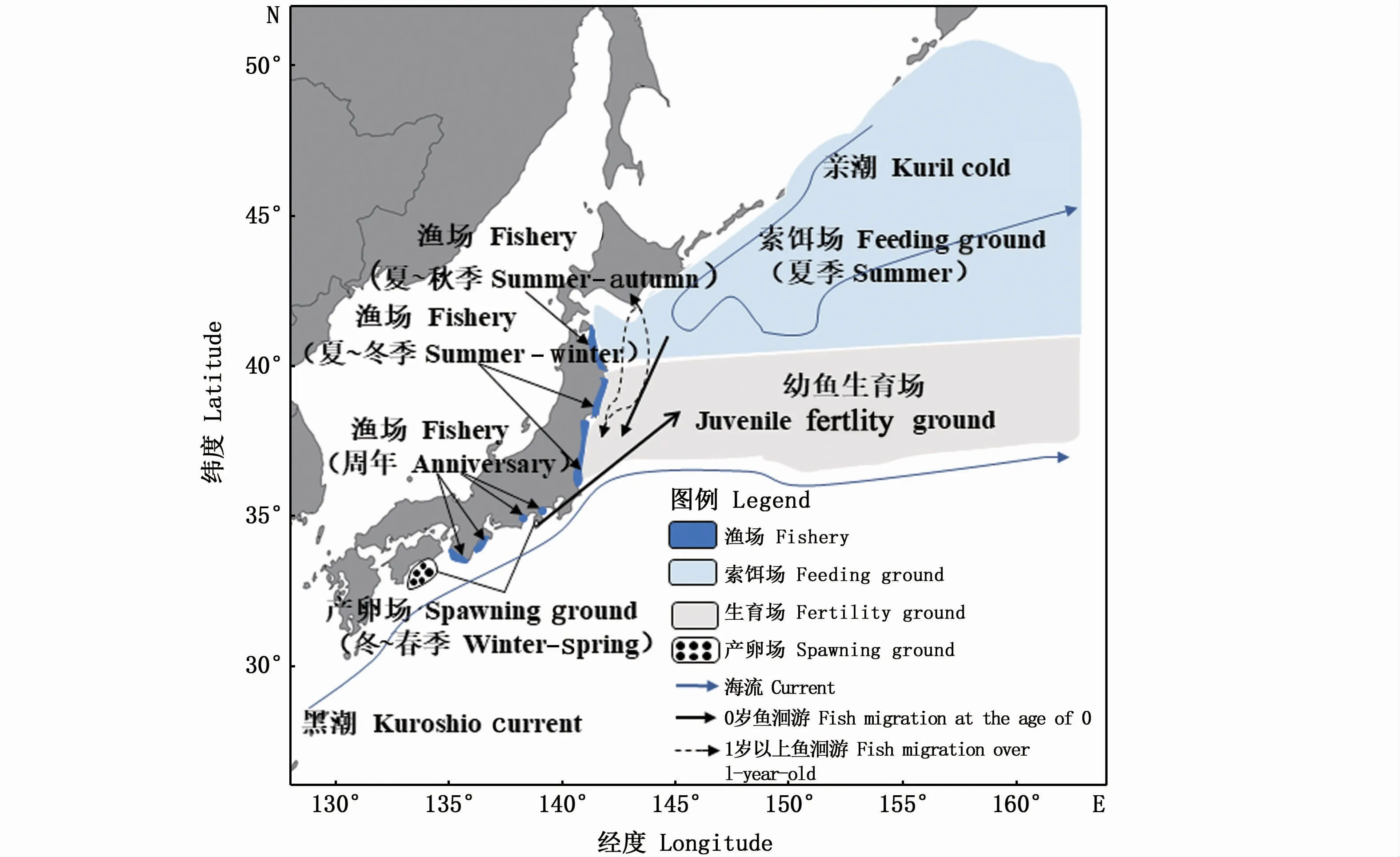
图1 太平洋群系沙丁鱼分布、洄游及生活史和渔场形成的示意图Fig.1 Schematic diagram of the distribution,migration,life history and fishing ground for Sardina in the Pacific Ocean
准确估计鱼类亲体-补充量关系(stock and recruitment relationship,SRR)是相关渔业资源管理的重要基础[5],但是SRR中的参数估计也是渔业资源评估中最困难的任务之一[6]。大洋中上层鱼类的亲体量与补充量之间的关系受环境影响较大,在研究这类鱼群时通常加入关键环境因子来探索二者之间的关系。Ricker模型是一种研究亲体量与补充量关系的模型,且该模型对环境有较显著的响应,因此有较多应用。如曹杰等[7]利用海水垂直平均温度(vertical average sea temperature,VAST)对印度洋黄鳍金枪鱼(Thunnus albacares)SRR的Ricker模型进行了优化,结果表明,VAST对其补充过程产生了一定的影响。林琴琴等[8]结合海表面温度(sea surface temperature,SST)和太平洋十年涛动(Pacific decadal oscillation,PDO)数据,利用Ricker模型对日本鲭(Scomber japonious)太平洋群系的亲体-补充量模型比较分析,发现两种环境因子的Ricker模型能更好的描述亲体-补充量关系。
众多研究表明,大洋中上层沙丁鱼的亲体量与补充量变化也显著地受其栖息地环境影响[9]。DEYLE等[10]演示了利用多元SSR检测环境变量对种群动态的影响,结果显示,SST对沙丁鱼的管理是一个重要的环境指标,SST或PDO可以提高补充量预测精度。YATSU等[11]应用Ricker模型研究表明,北太平洋海域的沙丁鱼补充量状况受其亲体量和SST的影响。上述研究结果都是在单一环境因子存在下得出,没有考虑双因子环境条件共同驱动对SRR的影响。为此,本研究利用SST与PDO双环境因子,探索太平洋群系沙丁鱼的SRR关系,以期为更好地预测、利用和保护沙丁鱼资源提供技术支撑。
1 材料与方法
1.1 数据来源
沙丁鱼亲体量和补充量数据来源于2016年度太平洋群系资源评价数据集,时间跨度为1996—2015年。1996年之后的几年中,沙丁鱼亲体量的水平呈现先增后减的趋势,而在2000年到2008年,沙丁鱼亲体量逐年减少,一直处于低水平状态,但2008—2014年亲体量出现持续补充,同时捕捞强度有所下降,使其在2014年增加到103万t。此外,2015年极高的补充量使得资源量剧增至约186万t[12]。
沙丁鱼的产卵盛期为2—4月,本研究只考虑外部环境因子对沙丁鱼产卵期的同步效应,因此采用1996—2015年2—4月份的SST和PDO作为沙丁鱼SRR关系的关键影响因子。其中,SST数据下载于哥伦比亚大学气候数据库(http://iridl.Ldeo.columbia.edu/SOURCES/.IGOSS/.data_products.Html),海域为142°~165°E、35°~40°N,空间分辨率为1°×1°,时间分辨率为月。PDO数据来源于日本气象厅(http://www.jma.go.jp/jma/index.html),时间范围为1996—2015年,时间分辨率为月。
1.2 SRR模型
选用Ricker模型模拟沙丁鱼SRR关系,未加入和加入环境因素的公式分别为

式(1)、式(2)中:α,β是待估计参数,R为补充量,S为亲体量。式(2)是优化后的亲体-补充量模型,式中e∑ni=1γixi表示环境特征函数,xi为影响因子,n为影响因子的数量,γi为待估计环境系数。因为太平洋群系沙丁鱼SRR受其产卵场SST和PDO的影响,构建4种SRR模型,分别是未考虑环境因子的Ricker模型(Ricker)、考虑受产卵场SST的影响的Ricker模型(SST_Ricker)、考虑受PDO影响的Ricker模型(PDO_Ricker)、考虑同时受SST和PDO影响的Ricker模型(SST_PDO_Ricker)。
1.3 模型评价
利用赤池信息量准则(Akaike information criterion,AIC)检验模型差异,具有最小AIC值的模型为最优模型[13]。同时利用留一交叉验证法(leave-one-out cross validation)评价模型稳定性,即从N个观测数据中选择一个作为验证数据,然后使用剩下的观测数据拟合模型,并用最先被排除的那个观测值计算绝对残差,如此重复N次[14],本次观测数据为20年的数据,即N为20。绘制各个模型20次AIC和绝对残差的箱线图,选择最优模型。
2 结果与讨论
2.1 最优SRR模型
同时考虑SST和PDO影响的沙丁鱼太平洋群系SSR关系模型(SST_PDO_Ricker)的精度最高,其AIC均值为47.76,绝对残差的均值为35.01;留一交叉验证显示,SST_PDO_Ricker模型极少出现异常情况,最为稳定(图2)。因此选用SST_PDO_Ricker模型为最终模型。

图2 不同环境影响下沙丁鱼太平洋群系亲体-补充量关系模型AIC值(a)与绝对残差值(b)Fig.2 AIC(a)and residual values(b)for Pacific Sardina of four SRR models
2.2 环境因子对沙丁鱼SRR关系的影响
通过最优模型的环境系数(γPDO=0.50,γSST=1.03)发现,沙丁鱼太平洋群系产卵期的SST和PDO对其补充量均有正影响,且SST的影响作用要大于PDO(P<0.05,表1),即2—4月份产卵场平均温度和平均PDO越高,当年的沙丁鱼太平洋群系的补充量越高。

表1 不同模型的参数值Tab.1 Parameter values of different models
1996—2015年这20年间沙丁鱼太平洋群系产卵场的平均SST和平均PDO能够明显反映其补充量的变化规律。总体来说,沙丁鱼补充量分布较为分散,其资源量进入20世纪90年代后有所减少,此时SST值均在平均值以下,尽管PDO处在高水平,但波动明显不够稳定,造成在1994年以后补充量进一步减少,即从2002年到2006年一直保持在10万t左右的低水平。在2007—2014年,SST高于平均值并持续上升,有利于沙丁鱼补充量的发生。此外,2015年极高PDO使得补充量有了更大地提高(图3)。

图3 环境因子与沙丁鱼亲体-补充量之间的关系Fig.3 Relationship between environmental factors and Sardina’s parental volume-recruitment
当PDO出现暖期时,北美大陆附近海面的水温就会异常升高,而北太平洋洋面温度却异常下降,导致沙丁鱼补充量的增加,当PDO出现冷期时,情况正好相反[15]。相比PDO,沙丁鱼补充量受SST的影响程度更大,当海表温度相对较高处于平均温度之上时,其补充量会有明显升高。所以,在太平洋,随着海表温度的升高,沙丁鱼的数量往往会增加,而温度上升主要对鱼群的产卵有一定影响,并且产卵场还会随着其资源水平的变化而变化。在20世纪30— 40年代,由于气候温暖,使得加利福尼亚洋流中的沙丁鱼数量激增,而在50—70年代,气候寒冷,它们的数量不断减少。在1999年,东北太平洋进入了一个新的“寒冷”状态,此状态不利于沙丁鱼的产卵和补充,在海洋温度较高的年份,0岁沙丁鱼的数量明显增加,并且与小型沙丁鱼相比,大型和中型沙丁鱼更容易在较低的表面盐度和温度下被捕获。对于浮游鱼类而言,温度与卵的丰度和中小生活史阶段显著相关,各阶段渔获量与温度均呈正相关关系[9]。
除了SST和PDO对太平洋群系的沙丁鱼补充量有影响外,还有一些其他的环境因子可能也对其有影响。此外,相同环境因子的强弱变化可能也会对其产生影响,所以关于SRR影响的研究还有待进一步深入。了解气候变化在年际和年代际时间尺度上对鱼类补充量的影响是了解鱼类种群动态和海洋生态系统的基础[16-17],为了更加准确地掌握环境因子对太平洋群系沙丁鱼资源的影响,也应对该群系沙丁鱼的一些行为以及综合其他的一些因子,例如食物来源等进行更加深入的研究,以实现对其资源的评估与利用。
