瓯江口刀鲚渔业生物学分析
2022-08-11陈伟峰蔡继晗张石天
陈伟峰,蔡继晗,叶 深,张石天,倪 勇,彭 欣,陈 坚
(1.浙江省海洋水产养殖研究所,浙江温州 325005;2.浙江省近岸水域生物资源开发与保护重点实验室,浙江温州 325005;3.温州市渔业技术推广站,浙江温州 325000;4.中国水产科学研究院东海水产研究所,上海 200090)
瓯江口地处西太平洋亚热带北部,其间散落着诸多岛屿,加上瓯江径流带来大量营养盐,是鱼类索饵、产卵、繁育的良好场所[1]。但随着经济社会发展对海域的侵占和长期高强度的捕捞行为,导致海域渔业资源种群结构发生巨大变化[2],以往常见的大型经济种类刀鲚(Coilia nasus)、带鱼(Trichiurus japonicus)和大黄鱼(Larimichthys crocea)等资源衰退明显,取而代之的是低龄化、低值化、小型化的鱼类,鱼类群落结构稳定性日益受损[3-4]。与此同时,随着社会经济的高速发展,瓯江口岸线被开发的力度越来越大。近岸码头林立,港口运输、沿海工业和滩涂围垦开发等人类活动影响,生态功能逐渐散失。
刀鲚隶属鲱形目,鳀科,鲚属,主要分布在中国、日本和朝鲜半岛[5]。近年来,关于刀鲚的研究主要集中在长江口海域,研究内容包括种群和资源变动[6]、生殖特征[7]、年龄鉴定[8]等。有关瓯江口刀鲚研究较少见,沈盎绿等[9]曾报道,瓯江口分布鱼类64种,鱼类种类以暖水种为主,夏季刀鲚在各调查站位出现率大于60%,鱼类多集群于瓯江口东侧区域。陈伟峰等[10]调查结果显示,瓯江口刀鲚多集中河口盐度较低区域。本文根据2017—2019年在瓯江口开展的12个航次渔业资源调查项目数据,对刀鲚种群参数进行估算,旨在丰富近海刀鲚基础研究数据,为更有效地保护刀鲚提供理论支持。
1 材料与方法
1.1 调查站位和调查方法
数据来自2017年2月至2019年11月的“瓯江口海域渔业资源本底调查”项目。共设调查站位11个(121°00′~121°54′E、27°45′~28°06′N),站位布设如图1所示。调查船为“浙乐渔休00105”底拖网船,调查船主功率为202 kW,吨位为128 t。调查网具为底拖网,水下网口面积约为6 m(宽)×1.5 m(高),网囊网目为2.5 cm,每站拖网时间为0.75 h,拖速控制在3~4 n mile左右,调查均于白天进行。

图1 瓯江口海域刀鲚资源调查站位Fig.1 Sampling stations of Coilia nasus in Oujiang River Estuary
1.2 数据处理方法
刀鲚资源密度计算公式为:
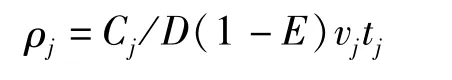
式中:ρj为j站位的资源密度(kg·km-2或103尾·km-2),Cj为j站位的渔获量(kg)或尾数,D为网口水平扩张宽度(km),vj为j站位拖网过程的平均船速(km·h-1),tj为j站位拖网时长(h),E为鱼类的逃逸率(0.5)[12]。
刀鲚体长和体质量的组距[13-14]。计算公式为:
体长组距(Snedecor)=Range/(Range/SD×4)
体质量组距(Sturges)=Range/(1+3.322 LgN)
式中:Range为体长(体质量)的全距,SD为体长(体质量)的标准差,N为总尾数。
刀鲚的体长与体质量的计算公式为:

式中:W为刀鲚的体质量(g),L为刀鲚的体长(mm),a为生长的条件因子,b为幂指数。
刀鲚生长方程计算公式为:

式中:l∞和W∞分别为刀鲚的极限体长和体质量,k为平均生长速率,t0是体长或体质量为零时的理论年龄。
t0采用Pauly的经验公式[15]:

资源开发率E的计算公式为:
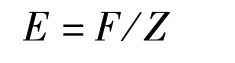
式中:Z为总死亡系数,F为捕捞死亡系数。
捕捞死亡系数的计算公式为:

式中:Z为总死亡系数,M为自然死亡系数。
总死亡系数Z通过体长变换渔获曲线(length-converted catch curve)[16]计算:

式中:Ni为第i体长组种渔获数量,dt为其从体长组下限生长到上限所需的时间,ti为第i体长组中值的年龄,回归方程中斜率的负值-b即为总死亡系数Z。
自然死亡系数M由Pauly的经验公式[15]计算获得:

式中:l∞为极限体长,k为平均生长速率,T为平均水温(℃)。2017—2019年瓯江口海域平均水温为18.9℃。
利用Beverton Holt模型预测相对单位补充渔获量(Y′/R)和相对单位补充生物量(B′/R)[17-18];本文采用了FiSAT II软件中的刀刃式选择假设模型。其中,E10为资源量减少10%时的开发率;E50为资源量下降到原始水平50%时的开发率;Emax表示获得最大渔获量时的开发率,其计算公式为:

式中:E为资源开发率,k为平均生长速率,lc为开捕体长,l∞为极限体长,M为自然死亡系数,Z为总死亡系数,F为捕捞死亡系数。
2 结果与分析
2.1 刀鲚资源季节变化
2017—2019年12个航次拖网调查共采集刀鲚1 008尾。调查海域各季节刀鲚平均资源量指数如图2所示,2017年的平均生物量为16.06 kg·km-2,平均丰度为0.82×103尾·km-2;2018年平均生物量为4.22 kg·km-2,平均丰度为0.13×103尾·km-2;2019年平均生物量为14.62 kg·km-2,平均丰度为0.85尾·km-2。按月份看,平均生物量最大值出现2017年11月,为29.25 kg·km-2,最小值出现在同年8月,为0.39 kg·km-2;丰度指数平均值最高出现在2019年2月,为1.61×103尾·km-2,最低值出现在2017年8月,为0.01×103尾·km-2。

图2 刀鲚生物量/丰度季节变化Fig.2 Seasonal variation of biomass and abundance of Coilia nasus
2.2 体长、体质量分布
瓯江口刀鲚的体长范围为15~357 mm,平均体长为186 mm。由Snedecor分组法计算的组距值为15.88 mm,因此将定,以15 mm为体长统计组距,绘制各组距的频率分布图(图3)。将组内数量占比大于5%的定位优势组,2017—2019年刀鲚的优势体长为90~270 m,占84.36%;刀鲚体质量范围为0.08~195.00g,优势体质量为0~60 g,占86.61%。

图3 刀鲚体长、体质量组成Fig.3 Frequency distribution of body length and body weight of Coilia nasus
体长与体质量的关系由幂函数拟合,其表达式为:W=3×10-6L3.0178(R2=0.926 4)(图4)

图4 刀鲚体长与体质量的关系Fig.4 Relationship between body length and weight of Coilia nasus
2.3 生长方程及死亡参数
基于刀鲚体长频数数据,求得Von Bertalanffy生长方程的生长参数l∞=362.25 mm,Vk=0.440,W∞=158.38 g,t0=-0.31a,Von Bertalanffy体长体质量生长方程分别为:lt=362.25[1-e-0.44(t+0.31)];Wt=158.38[1 -e-0.44(t+0.31)]3.0178(图5)。

图5 刀鲚Von Bertalanffy生长曲线Fig.5 Von Bertalanffy growth curve for Coilia nasus
根据体长变换渔获率曲线法(lengthconverted catch curve)计算总死亡系数Z,拟合的方程ln(N/dt)=6.916-1.33t(R2=0.931),-b=1.33即总死亡系数Z。根据Pauly的经验公式计算得出M=0.43,则捕捞死亡系数F=Z-M=0.90,又由资源开发率E=F/Z得出2017—2019年瓯江口刀鲚的资源开发率为0.68,体质量生长拐点年龄tλ值为2.20龄,相对应的体长为242.0 mm,体质量为47.00 g。对图6中的点计算观测值和预期值之比(图5)得出瓯江口刀鲚开捕体长为229.4 mm。

图6 根据体长变换渔获曲线估算总死亡系数Fig.6 Length-converted catch curve of Coilia nasus
2.4 刀鲚资源利用现状
根据相对单位补充渔获量(Y′/R)与开发率(E)的二维分析图显示了瓯江口刀鲚资源的当前开发程度(图7),E10=0.720,E50=0.401,Emax=0.803。以目前的开捕体长Lc=229.4 mm、E>0.68时,Y′/R随开发率的增大先增大后减小;当Lc=229.4 mm、E<0.68时,Y′/R随开发率的增大而增大。当E从当前的0.68提高至Emax(0.803)时,对应的Y′/R达到最大(Y′/R≈0.103),若降至E50时,对应的Y′/R则下降到0.085。从相对单位补充生物量(B′/R)的维持角度来看,当前刀鲚利用率为0.68,相对单位补充生物量(B′/R)约有20%得到保持;当Emax=0.803时,只有11.8%的相对单位补充生物量(B′/R)得到保持,而当E50=0.401时,约有50%的相对单位补充生物量(B′/R)能得到保持。

图7 瓯江口刀鲚相对单位补充渔获量(Y′/R)、相对单位补充量生物量(B′/R)和开发率(E)曲线图Fig.7 Curve of relative biomass/recruit,relative yield/recruit and exploitation ratio of Coilia nasus in Oujiang River Estuary
3 讨论
3.1 资源变化
颜文超等[19]报道了2013年5月和10月瓯江口各站位刀鲚的出现频率分别为20%和50%;刘志坚等[20]于2015年对瓯江口海域展开了底拖网渔业资源调查,其结果显示,刀鲚、凤鲚(Coilia mystus)为瓯江口春夏季的优势种。本文统计分析2017—2019年共12个航次渔业资源调查数据发现,各站位刀鲚的出现频率为42.5%,2017年和2018年刀鲚为常见种,2019年为优势种,资源变动无明显规律。2017—2018年5月、8月的平均生物量和丰度均较低,分别为5.51 kg·km-2和150尾·km-2,DT09和DT11站位未捕获刀鲚。究其原因,瓯江口刀鲚3—4月便开始溯河进入瓯江进行生殖洄游[21],至10月以后瓯江内水温下降则降海进行越冬;2019年2月份在DT09站位单网捕获刀鲚98尾,体长范围为88~280 mm,体质量范围为3.20~76.25 g,亦显示出该海域在冬季为刀鲚的越冬场所。通过对刀鲚出现站位的温盐进行分析,发现瓯江口海域的刀鲚仅出现在表层盐度31以下的站位,而温度则主要出现在表层水温25℃以下的站位。
3.2 生长参数
20世纪80年代以来,由于过度捕捞和环境污染等影响,海洋渔业资源量及群落结构同以往相比均发生了较大改变,包括刀鲚在内的传统经济鱼类资源衰退明显[22]。本研究结果显示,瓯江口海域刀鲚的体长和体质量范围分别为12~357 mm和0.08~195.00 g,明显小于长江刀鲚[7,24],与钱塘江口刀鲚较为接近[24];本研究结果显示,刀鲚的极限体长、体质量分别为362.25mm和158.38 g,k值为0.440(表1)。2017—2019年瓯江口刀鲚群体的k值小于靖江(k=0.55)、淀山湖(k=0.54)和长江口群体(k=0.51),但高于太湖(k=0.35)和鄱阳湖(k=0.24)群体;相对应的拐点年龄则呈现出鄱阳湖(tλ=4.00)和太湖(tλ=3.10)群体>瓯江口(tλ=2.20)群体>靖江(tλ=1.54)和淀山湖(tλ=1.37)群体[24,26-29]。整体上看,可以发现刀鲚所处的环境在离海域越远的情况下其平均生长速率(k)越小,而其拐点年龄(tλ)越大;这可能与海域中的饵料更丰富、更适宜刀鲚生长有关,亦可能与刀鲚洄游距离大导致消耗的能量更大有关。

表1 不同水域刀鲚生长参数比较Tab.1 Growth parameters of Coilia nasus from different areas
3.3 资源管理
詹秉义[11]研究认为,当总死亡系数与平均生长速率的比值(Z/k)≤3时,鱼类的自然死亡系数相对捕捞死亡系数的占比较高,即自然死亡是群体死亡的主要原因;而Z/k>3时,捕捞死亡系数相对自然死亡系数占比较高,即捕捞死亡是群体死亡的主要原因。本研究中,Z/k值为3.02,表明瓯江口海域刀鲚的捕捞死亡较自然死亡压力大。GULLAND[30]、PAULY和MORGAN[31]提出的鱼类最适开发率分别为E=0.5和E=0.286,METHANNA[32]认为,在相对单位补充渔获量与开发率关系曲线中,开发率E小于最大开发率Emax时,种群资源将可持续性发展,当开发率E大于Emax时,种群资源将会受到破坏。本研究估算出刀鲚的开发率为0.68,大于鱼类最适开发率,但未超过最大开发率,即瓯江口刀鲚资源已充分利用,但若适量减少捕捞压力可更好的保护瓯江口的刀鲚资源量。而通过浙江省海洋水产研究所的调查发现①浙江省海洋水产研究所调查结果,尚未公开发表,瓯江中下游的西洲岛-楠溪江口水域为刀鲚的天然产卵场,为更好地保护和利用刀鲚资源,建议在刀鲚的繁殖期对西洲岛-楠溪江口段水域设立刀鲚产卵场保护区。
