我国羊肚菌栽培历程及相关基础研究进展
2022-08-10刘伟何培新时晓菲许雯珺张亚于富强
刘伟 何培新 时晓菲 许雯珺 张亚 于富强*
我国羊肚菌栽培历程及相关基础研究进展
刘伟1何培新2时晓菲1许雯珺3张亚4于富强1*
(1. 中国科学院昆明植物研究所,云南 昆明 650201;2. 郑州轻工业大学食品与生物工程学院,河南 郑州 450001;3. 日本信州大学,长野 399-4598;4. 四川菌益侬农业科技有限公司,四川 成都 610000)
羊肚菌早期的驯化栽培因受制于基础研究不足而稳定性差。近年来,得益于羊肚菌人工栽培的快速发展,我国在羊肚菌种质鉴定与选育、交配型基因与生活史、菌种老化与质控体系、营养代谢与组学等研究领域取得了系列进展,并在一定程度上支撑了羊肚菌栽培区域和面积的快速扩张。然而,目前羊肚菌栽培仍然存在着优良种质匮乏、整体产量不稳定、病害与连作障碍凸显等问题,相关基础研究亟待开展与加强。
羊肚菌;栽培模式;种质资源;交配型;菌种老化;营养代谢;组学
羊肚菌(spp.)又称“真羊肚菌”(true morel),与之相对应的“假羊肚菌”(false morel)则指与羊肚菌形态类似的其他子囊菌,如马鞍菌(spp.)、钟菌(spp.)和鹿花菌(spp.)等[1]。羊肚菌富含氨基酸、腺苷等呈味物质,味道鲜美、口感独特,深受美食者的喜爱[2]。在欧洲,羊肚菌被认为是仅次于块菌(松露)(spp.)的美味食用菌;在北美,羊肚菌是蘑菇爱好者们认为最具魅力、被广泛认可的野生食用菌[3,4]。羊肚菌还具有一定的药用价值。明代《本草纲目》中就有羊肚菌“甘寒无毒,益肠胃,化痰利气,补脑提神”的记载[1,2]。现代医学研究表明,羊肚菌含有的活性成分,具有调节机体免疫力、抗氧化、抗菌、抗肿瘤、降血脂和保肝、护肝等功效[5]。
人类开展羊肚菌驯化栽培已有百余年历史[6]。国际上公认的羊肚菌驯化栽培技术突破是上世纪80年代美国科学家Ower等所做出的[7-10]。随后,美国企业应用该技术进行室内栽培生产,有过一些高产报道,但产出长期不稳定,于2006年前后全线停产[1,6]。本世纪初,以色列科学家Masaphy依照Ower的技术方案,进行羊肚菌室内栽培和发育形态学研究[11,12],但未有商业推广。2015年前后,曾经的美国羊肚菌栽培被Gourmet Mushrooms公司重新启用,但依然没有解决羊肚菌栽培中的稳定性问题。
我国的羊肚菌驯化栽培以2012年为界,2012年以前,以不同形式的仿生栽培为主,多数未进行持续研究和应用推广[1,14]。2012年前后,大田生态栽培模式逐渐成型,并开启了规模化示范推广之路[13],自此,技术不断完善,栽培区域也逐渐扩展至全国各地[13-15]。然而与羊肚菌栽培快速发展相比,相关基础理论研究却十分有限,这在很大程度上加剧了规模化生产的风险。值此羊肚菌产业发展十周年之际,回顾总结我国羊肚菌栽培研究和生产实践中所取得的成绩,分析存在的主要问题和不足,并据此提出下一步研发的重点领域和方向,为我国羊肚菌产业可持续发展提供参考。
1 羊肚菌栽培发展史与国内产业现状
1.1 羊肚菌栽培模式演变与内在机理
室内栽培。该模式源自于Ower,其栽培思路是首先通过“层级菌种”生产工艺培育菌核(图1-A),使用成熟的菌核作为“种子”,播种、浸水,进而诱导出菇[8-10],该技术也先后被一些公司用于羊肚菌的集约化栽培,但因出菇稳定性差而停滞不前[6,14]。目前,在大田生态栽培成熟模式和经验的基础上,我国有多家科研单位和企业投身到羊肚菌室内工厂化栽培的研发中。
仿生栽培。2010年以前,我国的羊肚菌栽培以纯仿生栽培、段木仿生栽培和林下仿生栽培为主,以野外播种、模拟并靠自然出菇为主要特征。其中以云南西北的段木仿生栽培最具代表性,其要点在于利用圆叶杨段木的树皮作为羊肚菌的营养源(图1-B),完成营养转化后出菇[16],虽单产不低,但因消耗木材量过大、不利于推广而逐渐退出市场。该模式仍然为后期的羊肚菌的营养代谢研究提供了有益参考[1]。

图1 羊肚菌三种栽培模式示意图(许雯珺绘制)
大田栽培。2012年前后,川渝地区羊肚菌大田生态栽培模式,以“外源营养袋”技术和“易出菇品种”为主要特征,逐渐成型。该模式在播种、发菌之后,将富含培养料的外源营养袋摆放在菌床上,使培养料中的养分通过菌丝输入菌床内的菌丝网络,完成后期有性生殖(出菇)所需能量的同化和转移(图1-C)。目前,各地依据不同的气候条件,相继发展出平棚、平棚套小拱棚、蔬菜大棚、林下、温棚(暖棚)等种植模式,羊肚菌栽培单产和稳定性逐步提升,“亩均产超千斤”的基地不断涌现[13-15,17,18]。
羊肚菌需要在营养“富足”与“贫瘠”的能量势差之间完成营养的转化[1,8,19-21],这与传统食用菌在同一个空间内实现营养同化吸收进而出菇的过程明显不同。虽然上述三种栽培模式的营养供给方式存在差异,但内在机理是一致的:美国室内栽培中,菌核在层级菌种的“贫瘠”土壤侧形成,营养“富足”的有机质层为能量供给体(图1-A),高能量的菌核结构发育成熟后进行刺激,诱导出菇;段木仿生栽培中,段木呈金字塔型堆积,菌种和土壤填充在段木之间,段木的树皮作为高营养势能环境,段木间的菌种和土壤构成了“贫瘠”区域,同样营造了营养势差,实现营养的同化吸收(图1-B);大田栽培是最为清晰的营养势能转化模式,遍布菌丝网络的土壤是营养“贫瘠”区域,外源营养袋作为“富足”营养势的存在,营养可以通过菌丝快速地从外源营养袋转移至土壤的菌丝网络和菌核中转化、储存,实现出菇(图1-C)。美国室内栽培是将菌核培育过程(能量转化过程)和出菇过程分离,而段木仿生栽培和大田栽培是两个过程的整合。
1.2 羊肚菌栽培快速发展科技主因
虽然美国上世纪80年代就实现了羊肚菌室内栽培的技术创新和应用生产,但因多种原因最终未能规模化推广。近年来,我国羊肚菌大田栽培得到快速发展并领先世界,这主要归因于“外源营养袋”补料技术的应用推广和易栽培出菇的种类选育。
“外源营养袋”补料技术创新与应用。Ower在其专利中简单描述有类似我国当前“外源营养袋”补料技术的栽培方式,但重点关注的是培育“菌核”和直接诱导出菇的技术方案[8-10];上世纪90年代Amir等发现是“营养势差”促进了菌核的形成[19,20]。我国科研人员在长期探索中发现了外源性补料技术的效果,并通过营养代谢等研究不断改进与完善这项技术,进而发展成为现今羊肚菌大田栽培中的主流模式[1]。“外源营养袋”补料技术在我国羊肚菌人工栽培快速发展中发挥了关键性作用。
易栽培出菇种类选育。目前全球已记录羊肚菌名称有300余个,其中多基因系统发育分析确定的系统发育种有78个,我国分布有至少30个系统发育种,但目前可栽培的仅限于黑色羊肚菌分支的7个种,即-6至-10分支和-13、-21分支[14,17,22,23],大面积人工栽培仅限于梯棱羊肚菌(,-10)、六妹羊肚菌(,-6)和七妹羊肚菌(,-7)[16]。早期羊肚菌人工驯化栽培中,大量的失败案例可能要归因于不适宜种类的选择。
1.3 羊肚菌栽培规模与市场
2012—2021年,我国羊肚菌人工栽培面积自3 000亩(1亩≈667平方米,下同)增至24.7万亩(图2),10年间增长了80余倍,栽培区域遍布全国20余个省份。2015年、2017—2018年羊肚菌栽培面积大幅增长,之后于2016年、2019—2020年均出现了一段增长停滞或缓慢期,这主要与羊肚菌栽培稳定性差、市场价格波动大导致种植户离场有关。

图2 2012年后我国羊肚菌的栽培面积和平均收购价格
由于适宜的气候条件和领先的栽培技术优势,羊肚菌栽培早期主要集中于川、滇地区(图3),一度占据全国栽培总面积的75%以上。但2018年后,作为主产区的四川盆地,因菌种质量参差不齐、栽培设施不完善、管理技术不到位、气候异常和连作障碍等原因,栽培大面积失败,从而导致栽培面积锐减,如2021年度较2020年度减少超2万亩,同比减少约30%。此情况在云南部分区域也有发生(图4)。
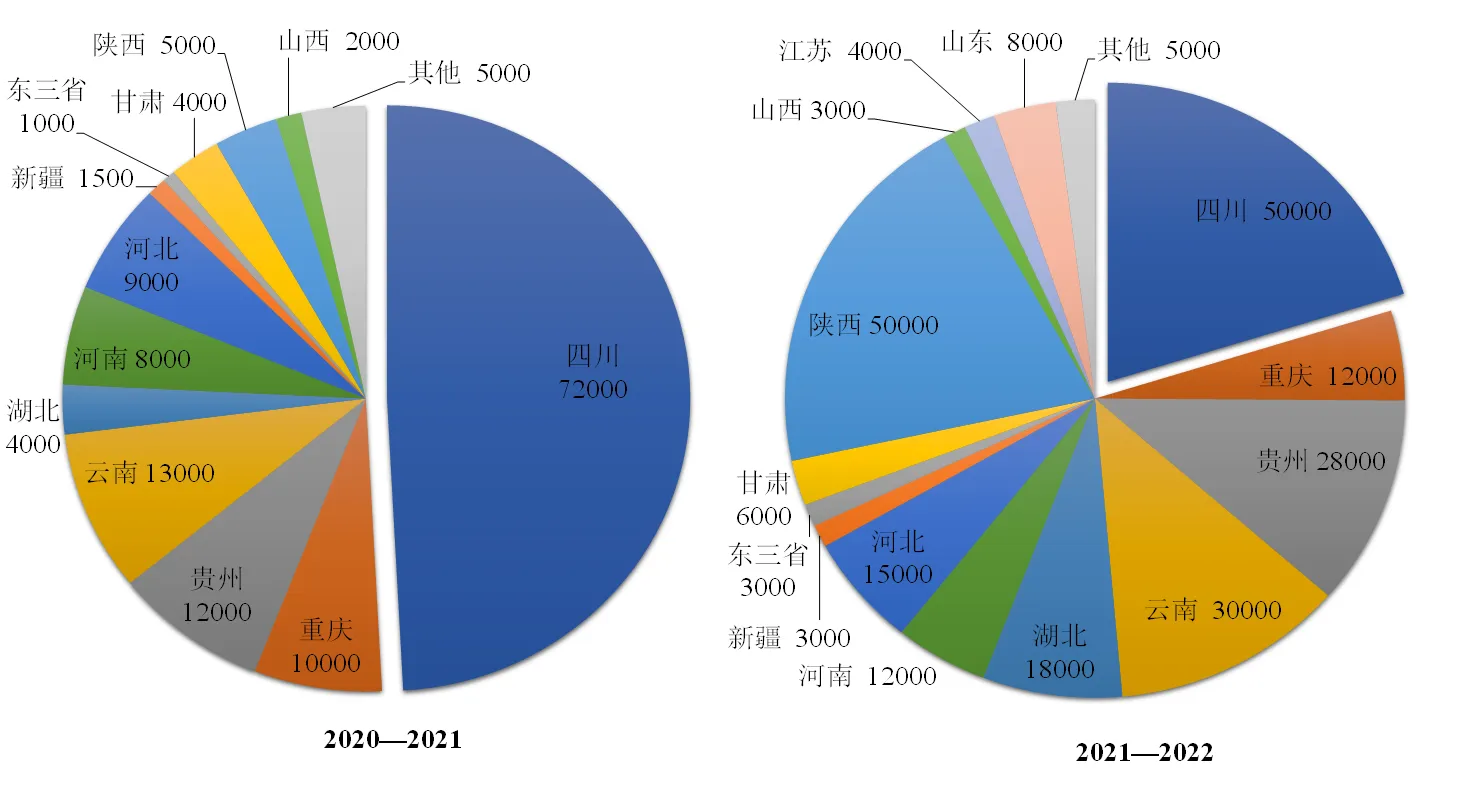
图3 2020—2021、2021—2022年度羊肚菌各栽培区域面积比较(单位:亩)
陕西是2021年度羊肚菌栽培面积增长最快的省份,这与其主要栽培区汉中地区的气候与四川相似,适宜羊肚菌栽培有关。四川盆地的连续种植失败致使部分基地北迁,湖北、河南、河北、甘肃和山西等地呈现稳定增长的趋势,但面积都未超过2万亩(图4)。其中,乡村振兴等政策的助推是一些省份栽培面积激增的主要原因。

图4 2015—2021年我国羊肚菌主产区的栽培面积变化
羊肚菌消费市场广阔,早期仅靠野生羊肚菌流通,贸易总量有限,国内外市场的干品价格一度徘徊在2 000元/千克左右;随着栽培面积和产量的不断增加,市场价格持续下跌(图2),2018年度最低降至600元/千克,这也在一定程度上导致此后2年种植面积增长停滞。之后,随着国内消费市场的逐渐打开,其价格又逐步回归,2020年在栽培面积和产量稳步提升且疫情严重影响消费的情况下,羊肚菌干品的收购价同比仍上涨约7%。
2 羊肚菌栽培的相关基础研究进展
2.1 羊肚菌的交配型基因结构和空间异质性
交配型基因是控制性发育的一个基因簇位点,子囊菌中通常含有两种类型的交配型位点Mat1-1和Mat1-2,分别编码α-box和HMG-box,如果两个交配型位点位于同一个细胞核内,即可在分子层面认定为同宗配合真菌(可自育),反之,则被认定为异宗配合真菌[24-26]。我国科研人员相继对多种羊肚菌开展了交配型结构分析和异宗配合生活史研究。结果表明,羊肚菌属大多数物种都是异宗配合生活史类型的真菌,单一的子囊孢子只含有两种交配型中的一种[26-32]。但也有研究指出,一些羊肚菌子囊果中存在着一定比例的单孢异核体[33],羊肚菌的生活史可能比我们想象的还要多样。
许多子囊真菌细胞中存在着交配型基因比例不均衡现象。He等[34]曾对梯棱羊肚菌全生活史周期中交配型基因比例进行研究,结果显示在各种类型分离得到的分离株和子实体组织中均呈现出显著的不均衡现象,认为这是细胞核水平的竞争所致。Du等[29]检测后指出,羊肚菌存在着一些只含有一种交配型的子囊果,这可能是两种交配型的严重不均衡所致。交配型基因比例严重不均衡的另一个表现是组织分离菌株的交配型丢失,刘伟等[35]对采自不同产区的21份羊肚菌子实体样品、348份分离株进行的交配型基因检测显示,两种交配型的检出率近似相等,但其中151份分离物只检测到一种交配型基因,推测这种现象除与细胞核水平的竞争有关外,也可能与不同交配型所代表的菌丝细胞并未充分融合有关[34]。
He等[36]通过观察梯棱羊肚菌孢子发育过程的核行为,指出梯棱羊肚菌的孢子虽为多核体,但均来自早期减数分裂后有丝分裂形成的八个细胞核之一,因而其孢子为同核体,这与交配型基因分型的结果相一致[26-32]。Ower等于1986年在羊肚菌室内栽培研究中指出单孢萌发的菌丝可以出菇[8]。但异宗配合生活史特点与单孢同核体和单孢出菇现象不符,为解释这种矛盾现象,刘伟等[26]随机选择了一些单孢菌株进行单孢及配对组合的栽培出菇实验,并对出菇的子囊果(F2)及新一代子囊孢子(F3)进行交配型检测,结果显示,虽然大多单孢菌株可以出菇,但单孢菌株只含有一种交配型基因,而出菇的子囊果却同时含有两种互补的交配型基因,推测互补的交配型基因是栽培场所无性孢子的传播引入,所谓单孢出菇是一种“假性出菇”;Liu 等[32]在对六妹羊肚菌单孢出菇的交配型基因检测中也发现了类似现象,出菇的子囊果“获得”了互补的交配型基因。
2.2 羊肚菌的无性孢子、菌丝和菌核
无性孢子(Mitospore,多被视作分生孢子Conidia)是羊肚菌生活史的一个重要部分,对出菇有着重要的影响。该孢子在大田栽培或野外环境下时常发生,然而在实验室条件下却难以诱导萌发[1,37,38]。研究推测,羊肚菌无性孢子可能扮演着“精子”的作用,因而不能视作分生孢子[38,39];但六妹羊肚菌的无性孢子似乎较容易萌发,具有一定的传播功能[40]。关于羊肚菌无性孢子功能和传播研究还有待深入。
菌丝是羊肚菌生活史中最为重要的阶段,与担子菌菌丝相比,子囊菌的菌丝似乎更容易老化。Hervey等在1978年最早发现普通羊肚菌()的部分子囊孢子萌发物存在生长停滞现象[41]。He等[42]曾对高羊肚菌()的老化特性进行系统研究,发现在继代培养过程中出现明显的发育性老化,指出菌株老化是一个涉及自噬和凋亡的系统性不可逆过程。随后,He等[43]研究了梯棱羊肚菌、六妹羊肚菌系列继代菌株的生物学特性,指出随着老化进程的加快,菌丝内脂质过氧化增加,菌核产生能力下降,色素形成加剧,栽培的产量也持续下降。刘伟等[44]对4种羊肚菌的52个菌株进行了系统老化评估,结果显示,除两个菌株在继代培养5 320 h后仍然存活外,其余菌株均在不同时间段内老化凋亡。钱可晴等[45]对梯棱羊肚菌继代培养的研究也指出,随着继代数的增加,菌丝生长速度、菌落形态和酶活性等都呈现下降趋势。以上研究表明,对羊肚菌来讲,菌丝老化应该是普遍现象。
羊肚菌生活史中的另一个重要结构是菌核,它的产生与培养基中营养的“贫瘠”和“富足”势差有关[19,20],这也是羊肚菌外源营养袋作用的主要机理。比较转录组和细胞学研究均表明,羊肚菌菌核内储存着大量的能源物质,扮演着抵御不良环境和储备能量的双重作用[46,47]。
2.3 羊肚菌种质研发和评价体系构建
在羊肚菌种质资源选育利用方面,2015—2019年四川省农业科院率先审定了包括梯棱、六妹和七妹在内的多个羊肚菌新品种[48]。2020年中华供销总社报道了‘中菌羊肚菌1号’新品种的选育[49]。近日,中国科学院昆明植物研究所驯化的‘昆植1号’和‘昆植2号’ 六妹羊肚菌也在贵州进行了成果鉴定[50]。以上品种多通过野生资源收集和驯化选育而来。He等[51]通过原生质体融合实现了六妹羊肚菌和梯棱羊肚菌的种间融合,为羊肚菌育种提供了一条新思路。然而,截至目前还没有被国家主管部门认定的羊肚菌新品种,这应该与羊肚菌菌种的稳定性差有关,如早期认定的不少优良品种,在3~5年规模化推广后,均因种性逐渐退化而退出历史舞台。
然而,与其他食用菌类群相比,羊肚菌的种质资源研究还十分薄弱,这也是制约新品种研发的一个重要因素。目前,国内外仅有少量关于羊肚菌种群遗传多样性的研究报道[52-57],涉及到可栽培品种的种群研究更少。刘伟等[53]曾用ITS系统发育分析和RAPD随机扩增多态性标记对来自全国12个省份的36个羊肚菌栽培菌株进行研究,系统地评价了其遗传多样性,指出当前我国栽培羊肚菌的遗传多样性较低,不利于产业的可持续发展。也有学者通过ISSR分子标记分别对野生或栽培的羊肚菌菌株进行了鉴定和遗传多样性分析[54-56]。Du等[57]利用梯棱羊肚菌基因组数据设计相关SSR分子标记,并评估了羊肚菌属5个种的遗传多样性,指出SSR在种间具有较好的多态性分辨率,可以用作不同品种的区分鉴定。
在实际生产中,选用不适宜栽培的羊肚菌种类,或者使用交配型基因缺乏的菌株或退化的菌种,都会导致栽培产量下降甚至绝产[16,25,43,44]。为此,刘伟等[18]总结提出了一套以身份(Identity)识别、交配型(Mating type)基因检测和菌株活力(Vitality)测定为核心的羊肚菌IMV生产菌株栽培适宜性评价系统,该系统可有效提升羊肚菌规模化生产的稳定性,降低风险。
2.4 羊肚菌病害研究
独特的栽培模式使羊肚菌子囊果更易遭受包括虫害、细菌病害和真菌病害在内的病虫害侵袭[15]。目前羊肚菌真菌病害的研究相对偏多,细菌病害和虫害的具体研究则较少。据统计,每年因真菌病害而导致羊肚菌栽培失败的面积约占总面积的25%。Guo等[58]最早报道了由引起的梯棱羊肚菌菌柄腐烂病。随后,He等[59]报道了因引发的羊肚菌白霉病,该病可同时在菌盖和菌柄发生;然而,对这种白霉病的鉴定分析显示,虽然在ITS序列上和具有99%的相似度,但其显微特征尤其是分生孢子的产生和形态,与属差异明显,推定该病原菌应为[60]。近期,Lan等[61]报道了引发的梯棱羊肚菌蛛网病,刘天海等[62]和Lü等[63]分别报道了由和引发的六妹羊肚菌腐烂病。虽然报道不少,但目前关于羊肚菌真菌病害的流行性、发病机理和防控办法等的研究还很有限。
2.5 羊肚菌组学研究
组学(Omic)是从整体的角度研究基因的组成、结构、功能和进化的学科,对应有基因组、转录组和代谢组等。羊肚菌的组学研究起步较晚,截至目前,公开发表的高质量基因组有:梯棱羊肚菌[30]、六妹羊肚菌[64,65]和粗柄羊肚菌[31]。此外,还有学者对梯棱羊肚菌、七妹羊肚菌、-21等进行了以交配型、营养代谢研究为目标的浅层测序[27, 29, 66-68][19, 34, 67-69]。刘伟[69]完成了中国羊肚菌属15个种的基因组测序与分析,美国JGI也完成了来自欧洲和北美约30个种的羊肚菌基因组测序与分析,相关结果已在整理中。未来这些基因组数据的分析结果将会大大加深我们对羊肚菌营养代谢、生活史、进化与发育等的理解。
羊肚菌其他组学研究集中在不同生长发育阶段的转录组、代谢组、蛋白组和微生物组比较分析,其中多以可栽培的梯棱羊肚菌和六妹羊肚菌为主要研究对象[46,70-82]。已有研究者对小海绵羊肚菌()的Cd2+耐受相关基因和代谢途径,以及羊肚菌与病原菌互作方面进行了比较转录组分析[78-80]。基于生长发育不同阶段的环境微生物组学分析,为羊肚菌大田栽培的“重茬”问题提供了一些有意义的参考[76,77,82]。张杰雄[83]对梯棱羊肚菌不同老化程度的菌株进行比较基因组和转录组分析,探究了衰老过程中的DNA和RNA层面的动态变化。其分析结果提示羊肚菌菌丝的衰老可能是由过氧化物酶体通路上相关蛋白(如PEX14、过氧化氢酶CAT等)的低表达引起的;低表达的PEX14会使得过氧化物酶体合成受阻,而低表达的CAT会导致过氧化物体清除活性氧能力降低,二者最终导致活性氧积累,进而诱导细胞进入程序性死亡。这些研究为深入理解羊肚菌的生长发育、代谢调控、环境互作和菌株老化等提供了支撑。
3 羊肚菌栽培研究面临的问题与展望
近十年来,在科研人员和产业人士的共同努力下,我国羊肚菌栽培得到快速发展,取得了令人瞩目的成绩。但作为一种新兴的栽培食用菌,与传统大宗食用菌品种相比,羊肚菌在优良种质选育、标准化生产体系建设和稳产高产等方面仍然存在着不小的差距,亟需加强相关基础与应用技术研究,支撑产业可持续发展。
3.1 优良种质选育与创制
系统地对推广种类进行遗传多样性分析,在组学研究技术日趋成熟的条件下,开发更加稳定的SSR、SNP或Inder标记,用于羊肚菌种质评价和分子标记辅助育种。通过野生资源驯化、诱变育种、杂交育种和基因工程育种等多技术手段开展羊肚菌优良种质选育与创新。
3.2 种质遗传不稳定性与老化
羊肚菌大田栽培的不稳定性众所周知,但关于其内在的遗传不稳定性却鲜有研究。在细胞、核酸和/或基因水平上研究羊肚菌种质遗传不稳定性,解析其快速老化的机理机制,通过化学或基因工程技术手段延缓老化或构建抗老化菌株,是未来提升和稳定栽培产量的重要方向。
3.3 营养代谢与互作机制
目前关于羊肚菌营养代谢的研究多基于比较组学,少有营养物质流、基因与代谢调控和其他微生物促生机制等方面的深入研究。同时,化感作用、营养物质和功能微生物群落失衡会导致羊肚菌栽培出现棘手的“重茬”问题,严重影响大田生产。通过多组学技术手段,研究羊肚菌与生物和非生物因子互作机制,可为羊肚菌栽培中产量不稳定和“重茬”问题的解决提供思路。
3.4 菌种质控技术体系与工厂化栽培
羊肚菌具有异宗配合、易退化、基因型丢失、遗传不稳定和对环境敏感等特点,对菌种和营养袋生产的质量控制具有很高的要求,这也是影响我国羊肚菌产量及其稳定性的重要因素,亟需研发制定优良种质评价体系以及制种、储存和运输等标准和技术规范。
羊肚菌的生长周期短、中低温出菇和市场价格高等特点,使其较适合发展工厂化栽培模式,但目前进展仍然缓慢,究其原因还是种质创新、营养代谢、与生物和非生物因子互作机制等基础研究不足所致。
[1] 刘伟, 张亚, 何培新. 羊肚菌生物学与栽培技术[M]. 长春: 吉林科学技术出版社, 2017: 340.
[2] Rotzoll N A, Dunkel T, Hofmann T. Quantitative studies, taste reconstitution, and omission experiments on the key taste compounds in morel mushrooms (Fr.)[J]. Journal of Agricultural and Food Chemistry, 2006, 54(7): 2705-2711.
[3] Pilz D. Ecology and management of morels harvested from the forests of Western North America[M]. Portland: General Technical Report, 2007: 161.
[4] Kuo M. Morels[M]. Ann Arbor: University of Michigan Press, 2005: 205.
[5] 蔡英丽, 马晓龙, 刘伟. 羊肚菌营养价值与保健功能[J]. 食药用菌, 2021, 29(1): 20-27.
[6] 谭方河. 羊肚菌人工栽培技术的历史、现状及前景[J]. 食药用菌, 2016, 24(3): 140-144.
[7] Ower R. Notes on the development of the morel ascocarp:[J]. Mycologia, 1982, 74(1): 142-144.
[8] Ower R D, Mills G L, Malachowski J A. Cultivation of: United States, EP86903077[P]. 1986-04-28.
[9] Ower, R.D., Mills G L, Malachowski J A. Cultivation of: United States,US07/217840[P]. 1988-07-12.
[10] Ower, R.D., Mills G L, Malachowski J A. Cultivation of: United States, US4866878[P]. 1989.
[11] Masaphy S. External ultrastructure of fruit body initiation in[J]. Mycological Research, 2005, 109: 508-512.
[12] Masaphy S. Biotechnology of morel mushrooms: successful fruiting body formation and development in a soilless system[J]. Biotechnology Letters, 2010, 32(10): 1523-1527.
[13] 刘伟, 张亚, 蔡英丽. 我国羊肚菌产业发展的现状及趋势[J]. 食药用菌, 2017, 25(2): 77-83.
[14] 刘伟, 蔡英丽, 张亚, 等. 我国羊肚菌人工栽培快速发展的关键技术解析[J]. 食药用菌, 2018, 26(3): 142-147.
[15] 刘伟, 蔡英丽, 何培新, 等. 羊肚菌栽培的病虫害发生规律及防控措施[J]. 食用菌学报, 2019, 26(2): 128-134.
[16] 程远辉, 赵琪, 杨祝良, 等. 利用圆叶杨菌材栽培羊肚菌初报[J]. 中国农学通报, 2009, 25(21): 170-172.
[17] 何培新, 刘伟, 蔡英丽, 等. 我国人工栽培和野生黑色羊肚菌的菌种鉴定及系统发育分析[J]. 郑州轻工业学院学报(自然科学版), 2015, 30(3): 26-29.
[18] 刘伟, 蔡英丽, 马晓龙, 羊肚菌生产菌株栽培适宜性评价系统[J]. 轻工学报, 2022, 37(3): 50-57.
[19] Amir R, Levanon D, Hadar Y, et al. The role of source-sink relationships in translocation during sclerotial formation by[J]. Mycological Research, 1994, 98(12): 1409-1414.
[20] Amir R, Levanon D, Hadar Y, et al. Factors affecting translocation and sclerotial formation in[J]. Experimental Mycology, 1995, 19(1): 61-70.
[21] Tan H, Kohler A, Miao R, et al. Multi-omic analyses of exogenous nutrient bag decomposition by the black morelreveal sustained carbon acquisition and transferring[J]. Environmental microbiology, 2019, 21(10): 3909-3926.
[22] 杜习慧, 赵琪, 杨祝良. 羊肚菌的多样性、演化历史及栽培研究进展[J]. 菌物学报, 2014, 33(2): 183-197.
[23] 杜习慧. 黑色羊肚菌支系的物种资源、生殖模式和遗传多样性研究进展[J]. 菌物研究, 2019, 17(4): 240-251.
[24] Metzenberg R L, Glass N L. Mating type and mating strategies in[J]. Bioessays, 1990, 12(2): 53-59.
[25] Du X H, Yang Z L. Mating systems in true morels ()[J]. Microbiology and Molecular Biology Reviews, 2021, 85(3): e00220-20.
[26] 刘伟, 蔡英丽, 何培新, 等. 梯棱羊肚菌单孢及杂交群体的栽培出菇试验和极性分析[J]. 菌物研究, 2019, 17(1): 7.
[27] Chai H, Chen L J, Chen W, et al. Characterization of mating-type idiomorphs suggests that,-20 andare heterothallic[J]. Mycological Progress, 2017,16(7): 743-752.
[28] Chai H, Chen W, Zhang X, et al. Structural variation and phylogenetic analysis of the mating-type locus in the genus[J]. Mycologia, 2019, 111(4): 551-562.
[29] Du X H,Zhao Q, Xia E H, et al. Mixed-reproductive strategies, competitive mating-type distribution and life cycle of fourteen black morel species[J]. Scientific Reports, 2017, 7: 1493.
[30] Liu W, Chen L F, Cai Y L, et al. Opposite polarity monospore genomesequencing and comparative analysis reveal the possible heterothallic life cycle of[J]. International Journal of Molecular Sciences, 2018, 19(9): 2525.
[31] Liu W, Cai Y L, Zhang Q Q, et al. Subchromosome-scale nuclear and complete mitochondrial genome characteristics of[J]. International Journal of Molecular Sciences, 2020, 21(2): 483.
[32] Liu Q Z, Q S, He G Q, et al. Mating-type genes play an important role in fruiting body development in[J]. Journal of Fungi, 2022, 8(6): 564.
[33] 刘萍, 马渊浩, 赵永昌, 等. 羊肚菌单孢菌株的性亲和与营养体亲和特性[J]. 食用菌学报, 2021, 28(1): 40-47.
[34] He P, Cai Y L, Yu F Q, et al. Spatial and temporal disequilibrium of nuclear distribution in heterothallic[J]. Journal of Agriculture and Food Research, 2021, 6: 100240.
[35] 刘伟, 蔡英丽, 何培新, 等. 羊肚菌组织分离物交配型基因缺失现象分析[J]. 食用菌学报, 2020, 27(3): 7.
[36] He, P X, Wang K, Cai Y L, et al. Live cell confocal laser imaging studies on the nuclear behavior during meiosis and ascosporogenesis inunder artificial cultivation[J]. Micron, 2017, 101: 108-113.
[37] Molliard M. Mycelium et forme conidienne de la morille[J]. CR Hebd Seances Acad Sci, 1904, 138: 516-517.
[38] Carris L M, Peever T L, McCotter S W. Mitospore stages of,andin the inland Pacific Northwest USA[J]. Mycologia, 2015, 107(4): 729-744.
[39] 刘伟, 蔡英丽, 何培新, 等. 梯棱羊肚菌无性孢子形态及显微结构观察[J]. 菌物研究, 2016, 14(3): 157-161.
[40] Yuan B H, Li H, Liu L, et al. Successful induction and recognition of conidiation, conidial germination and chlamydospore formation in pure culture of[J]. Fungal Biology, 2021, 125(4): 285-293.
[41] Hervey A, Bistis G, Leong I. Cultural studies of single ascospore isolates of[J]. Mycologia, 1978, 70(6): 1269-1274.
[42] He P, Cai Y L, Liu S M, et al. Morphological and ultrastructural examination of senescence in[J]. Micron, 2015, 78: 79-84.
[43] He P X,Yu M, Cai Y L, et al. Effect of aging on culture and cultivation of the culinary-medicinal mushrooms,and(Ascomycetes)[J]. International Journal of Medicinal Mushrooms, 2019, 21(12): 1089-1098.
[44] 刘伟, 何培新, 蔡英丽, 等. 羊肚菌衰老特性的测定分析[J]. 食药用菌, 2021, 29(1): 44-49.
[45] 钱可晴, 成盼盼, 范君佩, 等. 羊肚菌菌种继代培养代数与菌种退化关系[J]. 菌物研究, 2021,19(4): 277-284.
[46] Liu W, Cai Y L, He P X, et al. Comparative transcriptomics reveals potential genes involved in the vegetative growth of[J]. 3 Biotech, 2019, 9(3): 81.
[47] He P X, Wang K, Cai Y L, et al. Involvement of autophagy and apoptosis and lipid accumulation in sclerotial morphogenesis of[J]. Micron, 2018, 109: 34-40.
[48] 陈影, 彭卫红, 甘炳成, 等. 羊肚菌新品种‘川羊肚菌1号’[J]. 园艺学报, 2016, 43(11): 2289-2290.
[49] 刘春丽, 刘绍雄, 李建英, 等. 羊肚菌新品种“中菌羊肚菌1号”选育[J]. 中国食用菌, 2020, 39(10): 22-26, 36.
[50] 羊肚菌新品种介绍[J]. 菌物研究, 2021, 19(4): 214, 295.
[51] He P, Yu M, Wang K, et al. Interspecific hybridization between cultivated morelsandby PEG-induced double inactivated protoplast fusion[J]. World Journal of Microbiology and Biotechnology, 2020, 36(4): 58.
[52] Dalgleish H J, Jacobson K M. A first assessment of genetic variation among(Morel) populations[J]. Journal of Heredity, 2005, 96(4): 396-403.
[53] 刘伟, 兰阿峰, 张倩倩, 等. 羊肚菌栽培菌株遗传多样性分析及种特异性RAPD-SCAR标记开发[J]. 菌物学报, 2018, 37(12): 1650-1660.
[54] 杨燕, 田鸿, 张小平, 等. 基于ITS和ISSR的羊肚菌种质资源遗传多样性分析[J]. 西南农业学报, 2018, 31(10): 2004-2009.
[55] 曹君, 王红, 张敏, 等. 辽宁地区野生和栽培羊肚菌ITS序列及ISSR多态性分析[J]. 中国食用菌, 2019, 38(2): 37-41.
[56] 于凤鸣, 赵琪. 羊肚菌不同单孢及不同萌发孔菌丝间的遗传多样性[J]. 菌物学报, 2020, 39(6): 1117-1129.
[57] Du X H, Wang H, Sun J, et al. Hybridization, characterization and transferability of SSRs in the genus[J]. Fungal Biology, 2019, 123(7): 528-538.
[58] Guo M P,Chen K, Wang G Z, et al. First report of stipe rot disease oncaused byspecies complex in China[J]. Plant Disease, 2016, 100(12): 2530.
[59] He X L, Peng W H, Miao R Y, et al. White mold on cultivated morels caused by[J]. FEMS Microbiology Letters, 2017(5): 364.
[60] He P, Li C, Cai Y L, et al. First report of pileus rot disease on cultivatedcaused byin China[J]. Journal of General Plant Pathology, 2018, 84(1): 65-69.
[61] Lan Y F, Cong Q Q, Wang Q W, et al. First report ofcausing cobweb disease on cultivated[J]. Plant Disease, 2020, 104(3): 977.
[62] 刘天海, 周洁, 王迪, 等. 一种六妹羊肚菌的新型柄腐病害[J]. 菌物学报, 2021, 40(9): 2229-2243.
[63] Lü B, Sun Y, Chen Y, et al. First Report ofcausing rot ofin China[J]. Plant Disease, 2021, 102(3): 675.
[64] Han M, Baiyintala W. The whole-genome sequence analysis ofsextelata[J]. Scientific Reports, 2019.
[65] 王昊明, 范立刚, 董永镕, 等. 六妹羊肚菌PacBio基因组测序和DNA 6mA甲基化分析[J]. 菌物学报, 2021, 40(9): 2299-2316.
[66] Murat C, Payen T, Noel B, et al. Pezizomycetes genomes reveal the molecular basis of ectomycorrhizal truffle lifestyle[J]. Nature Ecology & Evolution, 2018, 2(12): 1956-1965.
[67] Du X H, Wu D, Kang H, et al. Heterothallism and potential hybridization events inferred for twenty-two yellow morel species[J]. IMA Fungus, 2020,11(1): 4.
[68] Wingfield B D,Bills G F, Dong Y, et al. Draft genome sequence of,,(syn.),, twostrains,,, Pcf. hyalina, and[J]. IMA Fungus, 2018, 9(1): 199.
[69] 刘伟. 梯棱羊肚菌生长发育及羊肚菌属的组学研究[D]. 武汉: 华中农业大学, 2020.
[70] 王金秋, 叶鸿亮, 梁道崴, 等. 梯棱羊肚菌子实体转录组测序及解析[J]. 食品科学, 2018, 39(18): 81-87.
[71] Deng K, Lan X, Chen Y, et al. Integration of transcriptomics and metabolomics for understanding the different vegetative growth in[J]. Frontiers in genetics, 2022, 12: 829379.
[72] Liu Q, He G, Wei J, et al. Comparative transcriptome analysis of cells from different areas reveals ROS responsive mechanism at sclerotial initiation stage in[J]. Scientific reports, 2021, 11(1): 9418.
[73] Hao H, Zhang J, Wang H, et al. Comparative transcriptome analysis reveals potential fruiting body formation mechanisms in[J]. AMB Express, 2019, 9(1): 103.
[74] Lü B B, Wu G G, Sun Y, et al. Comparative transcriptome and endophytic bacterial community analysis ofSH[J]. Frontiers in Microbiology, 2021, 12: 682356.
[75] Wang J, Xiao J, Geng F, et al. Metabolic and proteomic analysis of morel fruiting body () [J]. Journal of Food Composition and Analysis, 2019, 76: 51-57.
[76] Longley R, Benucci G M N, Mills G, et al. Fungal and bacterial community dynamics in substrates during the cultivation of morels () indoors[J]. FEMS Microbiology Letters, 2019, 366(17): fnz215.
[77] Benucci G M N, Longley R, Zhang P, et al. Microbial communities associated with the black morelcultivated in greenhouses[J]. PeerJ, 2019, 7: e7744.
[78] Xu H Y, Xie Z L, Jiang H C, et al. Transcriptome analysis and expression profiling of molecular responses to Cd toxicity in[J]. Mycobiology, 2021, 49(4): 421-433.
[79] Chen C, Fu R, Wang J, et al. Genome sequence and transcriptome profiles of pathogenic fungusreveal its interactions with edible fungus[J]. Computational and Structural Biotechnology Journal, 2021, 19: 2607-2617.
[80] Yu Y, Tan H, Liu T, et al. Dual RNA-Seq analysis of the interaction between edible fungusand its pathogenic fungusuncovers the candidate defense and pathogenic factors[J]. Frontiers in microbiology, 2021, 12: 760444.
[81] Cai Y L, Ma X L, Zhang Q Q, et al. Physiological characteristics and comparative secretome analysis ofgrown on glucose, rice straw, sawdust, wheat grain, and MIX substrates[J]. Frontiers in Microbiology, 2021, 12: 636344.
[82] Yu F M, Jayawardena R S, Thongklang N, et al. Morel production associated with soil nitrogen-fixing and nitrifying microorganisms[J]. Journal of Fungi, 2022, 8(3): 299.
[83] 张杰雄. 羊肚菌发育衰老机制研究[D]. 西安: 西北工业大学, 2021.
S646
A
2095-0934(2022)04-261-10
云南省科技厅基础研究专项面上项目(202101AT070541);2022年云南省博士后定向培养资助项目
刘伟(1984—),博士,主要从事大型真菌遗传育种与栽培学研究。E-mail:zhenpingliuwei@163.com。
于富强(1976—),博士,正高级工程师,主要从事大型真菌种质资源调查评价、收集保藏与创制利用研究。E-mail:fqyu@mail.kib.ac.cn。
