黄土高原丘陵区典型植物群落土壤粒径分布特征
2022-08-08沙国良魏天兴陈宇轩傅彦超
沙国良, 魏天兴, 陈宇轩, 傅彦超, 任 康
(1.北京林业大学水土保持学院,北京 100083;2.北京林业大学水土保持与荒漠化防治国家林业局重点实验室,北京 100083;3.山西吉县森林生态系统国家野外科学观测研究站,北京 100083)
土壤粒径分布特征是土壤最基本的物理属性,与土壤的水分运动、肥力发展和孔隙状况等有着密切关联[1-3],在地表侵蚀过程中起着重要作用。其中,土壤黏粒因其具备胶体性质,可有效促进土壤团聚体的形成,进而提升土壤结构的稳定性,增强其抗侵蚀能力[4-5]。相关研究表明,采用传统结构描述的方法并不能充分反映土壤粒径的分布特性,而分形理论可以实现土壤结构组成的定量描述,更加准确地表达土壤的结构形态和功能、整体与局部的相似性[6],逐步成为研究土壤粒径分布的有效途径。
黄土高原作为典型的黄土堆积区,在其独特气候条件和长达数千年的人为活动影响下,植被破坏严重,土壤侵蚀强烈,是世界上最严重的水土流失区之一。黄土高原大部分地区的土壤侵蚀模数超过1000 t·km-2·a-1,其中,丘陵沟壑区的土壤侵蚀尤为剧烈,侵蚀模数超过5000 t·km-2·a-1[7]。为治理水土流失,保护生态环境,该地区先后实施了一系列生态修复工程,并取得了巨大成就。截止目前,黄土高原的植被覆盖程度提升显著,入黄泥沙含量和土壤流失状况得以减缓[8-12]。油松(Pinus tabulaeformis)、山杏(Armeniaca sibirica)作为黄土高原的主要造林树种,具有适应性强、抗瘠薄、耐寒旱等特性[13]。然而,大面积的油松、山杏纯林种植会导致林分结构单一,林内生态系统稳定性弱,抗干扰能力差,土壤质量受限,影响林下植被生长等问题[14-15]。此类问题在黄土高原的水蚀风蚀交错区尤为突出,对此,国内外专家学者已围绕水土流失动态监测、土壤侵蚀和土壤养分计量特征等领域开展了大量研究工作[16-20],而针对不同林分配置模式下土壤粒径垂直分异的研究较为少见。
本研究选取油松纯林、山杏纯林、油松山杏混交林为研究对象,草地作为对照,利用激光衍射技术测定了不同植物群落0~200 cm土壤的粒度组成,计算了土壤的粒度参数和分形维数,并绘制了土壤粒配曲线,以期深入了解不同林分配置对土壤粒径垂直分布特征的影响,为黄土高原的生态重建工作提供理论依据。
1 材料与方法
1.1 研究区概况
研究区位于陕西省延安市吴起县(107°39′~108°33′E,36°33′~37°24′N),属温带大陆性季风气候,年均降水量483.4 mm,降水集中在夏秋两季,多为暴雨,年均气温7.8 ℃,年日照时数2435.7 h,≥10 ℃积温2883 ℃[21],海拔1222~1812 m,无霜期达146 d,土壤为黄绵土,土层深厚。研究区属典型黄土丘陵沟壑区,水土流失严重,自1999 年退耕还林实施后,形成了以沙棘(Hippophae rhamnoides)、油松、柠条绵鸡儿(Caragana korshinskii)和山杏等为主的人工植被群落[22]。样地基本信息见表1。
1.2 土壤样品采集与处理
于2020 年9 月在研究区内选取20 m×20 m 草地、油松林、山杏林和油松山杏混交林样地,为保证相对独立性,样地空间距离大于1 km,每个样地内部采用五点取样法,表层土0~20 cm 每10 cm 取样,20~200 cm 每20 cm 采集土壤样品,共计11 个土层,各土层重复采样3 次作为重复试验,同层土壤样品混合均匀后封装。经风干处理后,过2 mm 筛,去除砾石和植物细根,预处理土样使用H2O2去除有机质,HCl去除碳酸盐后,使用Mastersizer 3000激光粒度分析仪测定土壤粒度组成,每个样品重复测量3次,采用美国制土壤粒级分级标准输出结果[23],并输出粒径体积分布下5%、16%、25%、50%、75%、84%和95%累计值对应的土壤粒径(μm)。
1.3 分形维数的计算
土壤由不同大小土粒组成,具有孔隙结构,本身具有自相似性和非均一性,因此分形理论已广泛应用于土壤科学用以描述土壤的结构和功能[24]。土壤分形维数不仅可以反映土壤粒度分布的整体性特征还与土壤质地和土壤肥力等土壤属性具有相关性[25]。本文采用土壤颗粒体积分形模型对单重分形维数进行计算[26],计算公式如下:
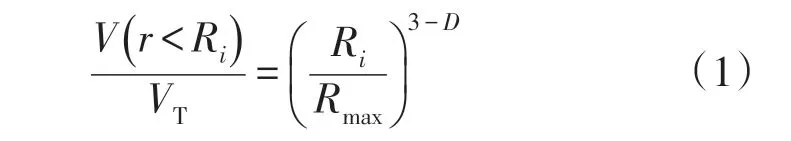
式中:V(r<Ri)为土壤粒径中小于Ri的体积分数;r为土壤粒径(mm);Ri为粒径分区中第i等级的粒径;VT为土壤颗粒总体积;Rmax为土壤粒径中的最大值(mm);D为分形维数。
1.4 土壤粒度特征参数的计算
将实验测定的土壤颗粒体积分布数据,根据Udden-Wenworth粒级标准,采用Krumdein对数转换法将粒径真值d转换为Φ,中值粒径使用Krumbeinn法计算,平均粒径、标准偏差、偏度值和峰态值采用Folk-Word 图解法计算。土壤粒度特征参数用以反映土壤粒度分布趋势、离散程度、分布形态[27]。
计算公式如下:
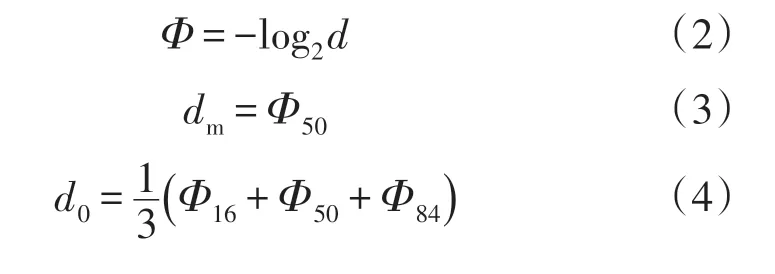
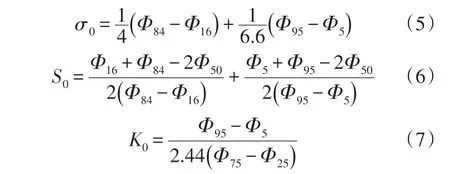
式中:d为土壤粒径(mm);dm为中值粒径;d0为平均粒径;σ0为标准偏差;S0为偏态值;K0为峰态值。Φ5、Φ16、Φ25、Φ50、Φ75、Φ84、Φ95为累积频率曲线中对应百分位下粒径的Φ值。
1.5 统计分析方法
采用IBM SPPSS Statistics 25 进行K-S检验验证数据正态性,单因素方差分析(One-way ANOVO)用来比较不同林地类型下的土壤粒度组成和粒度参数差异,经方差齐性检验后使用最小显著性差异法(LSD)进行显著性检验。使用R 3.6.3 进行线性回归,分析分形维数与各粒级含量之间的关系,并使用Origin 2019b绘制土壤粒度频率曲线。
2 结果与分析
2.1 土壤粒度组成
如图1所示,研究区土壤(0~200 cm)属粉壤土,不同植物群落下土壤的粒度组成按体积占比排序均为粉粒>砂粒>黏粒。其中粉粒含量为56.58%~71.67%,砂粒含量为21.37%~38.71%,黏粒含量最少为3.55%~6.96%。
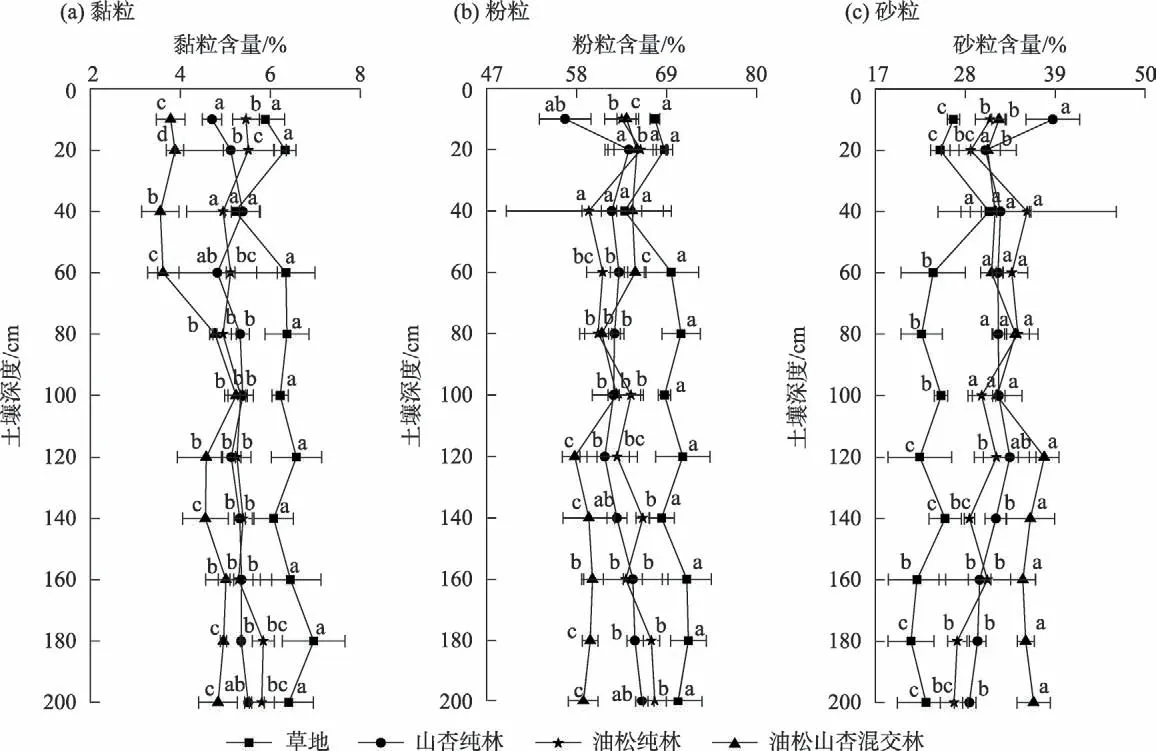
图1 不同植物群落0~200 cm土壤粒度组成Fig.1 Particle size composition of 0-200 cm soil under different plant communities
除个别土层外,0~200 cm 草地黏粒、粉粒含量显著高于油松纯林、山杏纯林和油松山杏混交林(P<0.05),砂粒含量均显著小于油松纯林、山杏纯林、油松山杏混交林(P<0.05)。山杏纯林和油松纯林除0~10 cm、10~20 cm土层外,黏粒、粉粒、砂粒含量均不存在显著差异(P>0.05)。油松山杏混交林黏粒含量显著小于其他林地(P<0.05),砂粒显著大于草地、油松纯林和山杏纯林(P<0.05)。深度达到100 cm后混交林粉粒含量显著小于纯林和草地(P<0.05)。
2.2 粒度参数与分形维数
如图2 所示,研究区土壤平均粒径为Φ4.97~5.50,中值粒径范围在Φ4.66~5.22,土壤颗粒集中分布在较粗范围内。显著性分析表明,除10~20 cm、20~40 cm土层外,草地平均粒径均显著大于油松山杏混交林、油松纯林和山杏纯林(P<0.05),10~100 cm 混交林与纯林平均粒径与中值粒径差异不显著(P>0.05),100~200 cm(除140~160 cm 土层外)混交林平均粒径与中值粒径显著小于纯林(P<0.05),10~200 cm山杏纯林与油松纯林平均粒径与中值粒径差异不显著(P>0.05)。
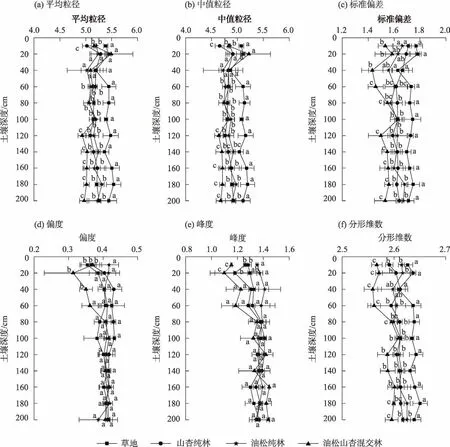
图2 不同植物群落土壤粒度参数Fig.2 Soil particle size parameters of different plant communities
草地、山杏纯林、油松纯林和油松山杏混交林,标准偏差范围分别为Φ1.70~1.75、Φ1.60~1.71、Φ1.55~1.67 和Φ1.43~1.58,平均标准偏差草地>山杏纯林>油松纯林>油松山杏混交林,土壤颗粒分选性等级均属较差级。显著性分析表明,40~200 cm 草地标准偏差显著大于其他林地(P<0.05),山杏纯林与油松纯林标准偏差差异不显著(P>0.05)。10~100 cm 混交林标准偏差与纯林无显著差异(P>0.05),100~200 cm 混交林标准偏差显著小于纯林和草地(P<0.05)。
草地、山杏纯林、油松纯林和油松山杏混交林,偏度范围分别为Φ0.35~0.42、Φ0.37~0.43、Φ0.38~0.42、Φ0.31~0.42 内,土壤粒度粗细分配对称性差,呈极正偏态。峰度范围在1.10~1.44,属尖窄水平,土壤颗粒分布集中。显著性分析表明,0~20 cm 混交林偏度和峰度均显著小于草地和纯林(P<0.05),20~200 cm各样地土壤偏度和峰度都不存在显著差异(P>0.05)。
草地、山杏纯林、油松纯林和油松山杏混交林,平均分形维数为2.63、2.60、2.61、2.58。显著性分析表明0~200 cm 草地分形维数显著大于纯林和混交林(P<0.05),10~200 cm 山杏纯林与油松纯林分形维数差异不显著(P>0.05),0~60 cm 混交林分形维数显著小于纯林和草地,80~100 cm 混交林分形维数与纯林和草地无显著差异(P>0.05)。
2.3 土壤粒径与分形维数的相关性
由表2 可知,各林地分形维数与黏粒含量具有极显著正相关关系(P<0.01),拟合优度(R2)草地>油松山杏混交林>山杏纯林>油松纯林。草地和油松纯林的分形维数与粉粒含量具有极显著正相关关系(P<0.01),而油松山杏混交林分形维数与粉粒含量呈显著负相关关系(P<0.05),山杏纯林分形维数与粉粒含量无显著相关性(P>0.05)。草地和纯林的分形维数与砂粒含量具有极显著负相关关系(P<0.05),油松山杏混交林分形维数与砂粒含量无显著相关性(P>0.05)。
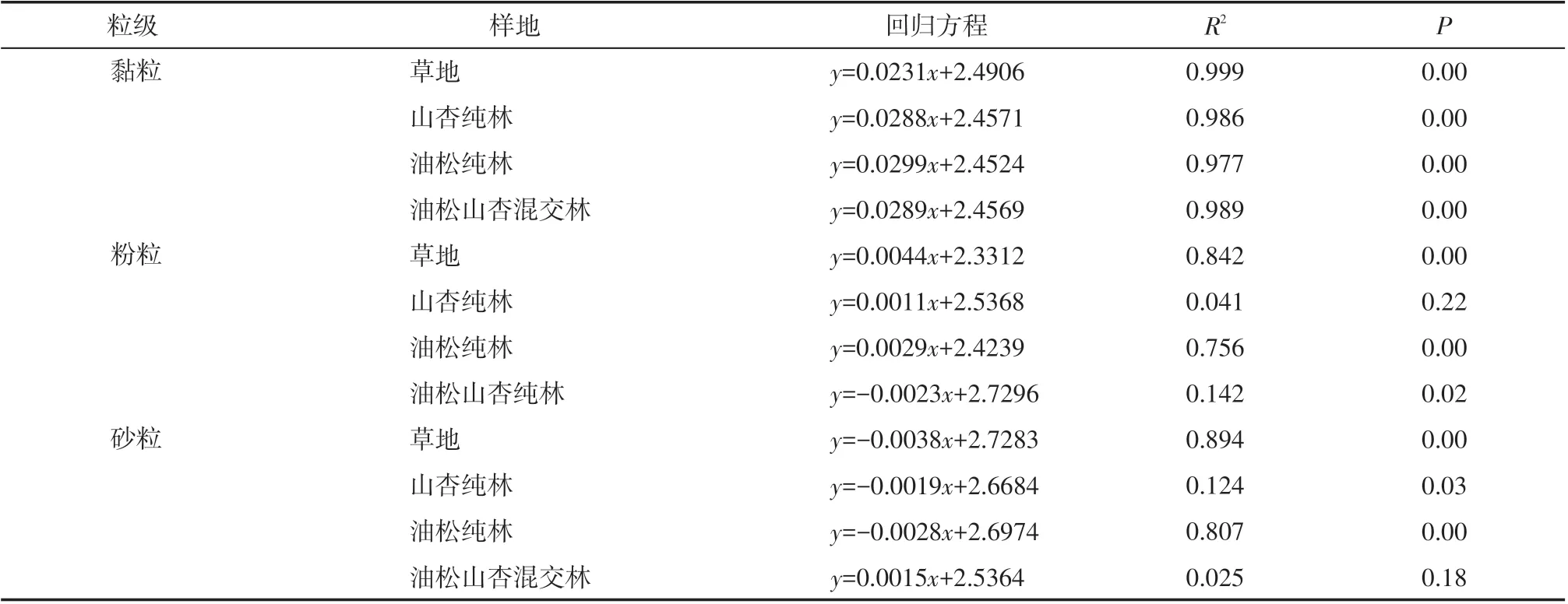
表2 土壤分形维数与粒级含量相关性Tab.2 Correlation between soil fractal dimension and particle size content
2.4 土壤粒度频率分布曲线
如图3 所示,除山杏纯林0~10 cm 属三峰型,研究区不同植物群落0~200 cm 土壤粒度频率曲线均属于双峰型,形态相似,曲线第一波峰均约为Φ10.50处取得,第一波峰平均峰值大小排序为草地>油松纯林>山杏纯林>油松山杏混交林,第二波峰均约在Φ4.50 附近出现,第二波峰平均峰值大小排序为油松山杏混交林>油松纯林>山杏纯林>草地。随深度增加,土壤粒度频率曲线变化集中在Φ7.90~8.00和Φ4.00~5.00。
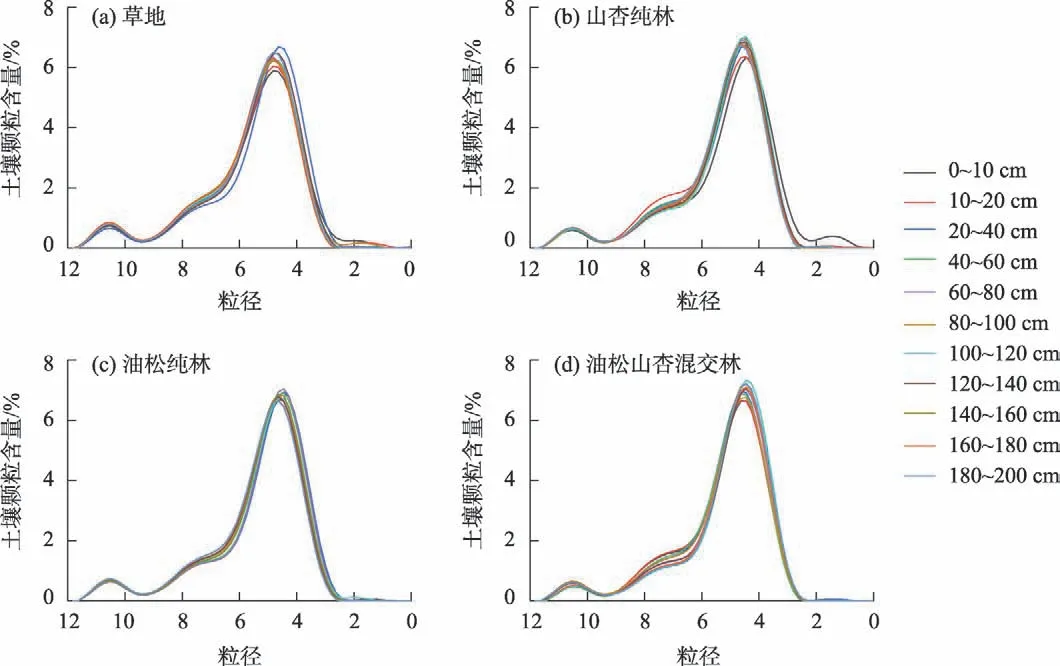
图3 不同植物群落土壤粒度频率分布曲线Fig.3 Frequency distribution curve of soil particle size in different plant communities
3 讨论
3.1 不同植物群落对土壤粒度组成的影响
土壤粒度组成是由外部环境要素(降水量、气温、地形、坡度等)与植被-土壤系统的内部要素(植被根系分泌物、枯落物、土壤微生物等)共同作用的结果,不同植物群落的土壤粒度组成反映了其生境差异[28]。研究区土壤(0~200 cm)粉粒含量最高,砂粒含量居中,黏粒含量最少,这主要与黄土的风成沉积过程有关,在此基础上不同植物群落的土壤粒度组成差异可归因于强烈的土壤侵蚀作用[29]。黄土高原水蚀风蚀作用强烈,强集中性降雨使土壤表层粗颗粒迅速风化崩解,细粒部分随入渗作用进入深层,或随地表径流流失[30]。在各土层深度上,草地土壤各粒级含量均显著区别于人工林,黏粒、粉粒含量显著大于人工林,砂粒含量小于人工林,与Xia 等[31]的研究结果一致,表明草地对土壤的细化作用强于人工林。土壤黏粒含量与团聚体稳定性呈正相关[32],本研究中草地土壤较高的黏粒含量将会促进稳定性团聚体的形成,增强土壤抗蚀能力[33],有利于水土保持。此外,退耕还林在一定年限内没有显著改善土壤粒度组成,造林树种的选择在黄土高原土壤质地改良上的作用可能存在过分高估的现象[34],而退耕还林还草,采用林草复合生态系统,可以增强植被-土壤生态系统稳定性,并提高土壤养分和持水性同时优化土壤结构[35],是控制黄土高原水土流失的有效措施。
土壤细颗粒与土壤水分和有机碳稳定性均具有显著相关性,黄土高原人工林生长受限于降水条件,为补充水分,植物根系向深层不断发展,在根系垂直增殖过程中,土壤砂粒含量多可调节水分渗透和向上毛细提升过程,使植物更好地从地下补充水分[36]。同时,在退耕还林过程中,土壤有机碳稳定性下降[37],使得土壤细粒含量下降,人工林地细粒组分少于草地。但随着林龄的增长,植被对土壤粒度的影响逐渐明显,土壤黏粒、粉粒含量将不断增加,砂粒含量将不断减少,成熟林阶段土壤粒度组成将达到最佳水平[38]。此外,植物根系也对土壤粒度组成具有重要影响[39],植物根系通过物理缠绕、串联网络和分泌根系分泌物等方式提高了土壤抗蚀能力,阻碍了土壤颗粒转化[40]。相关研究表明提高林下草本盖度可有效增强土壤抗冲性,林下草本根系主要分布在0~20 cm 土层,因此极大程度固定了表层土壤,减少了细粒的流失[41]。而在20~40 cm土层,随着草本根系分布的减少出现了土粒粗化的现象。本文中油松纯林和山杏纯林在不同土层土壤粒径差异不显著,这是因为山杏和油松作为耐旱树种,在根系结构和空间分布上具有相似性[42]。混交林由于根系发达,分布广泛,根系作用放大,使得土壤粒度组成偏粗,有助于植被吸水,缓解生理性干旱。
3.2 不同植物群落下土壤粒度参数的差异
研究区土壤平均粒径为Φ4.97~5.50,中值粒径为Φ4.66~5.22,土壤颗粒组成较细。偏度呈极正偏度,峰态尖窄,这是因为土壤粉粒含量极高,而黏粒含量极低,且砂粒中极细砂占比达70%,因此造成了粒度分布极不对称和波峰集中的现象[43]。标准偏差反映了土壤颗粒分散程度,研究区土壤分选性较差,而混交林的分选性显著优于纯林和草地,表明混交林可使土壤颗粒整体集中分布在某一区间,体现了其在改变土壤粒度组成方面的性能,与前人研究成果吻合[44]。同时不同树种间的混交将对土壤理化性质产生重要影响[45],因此针对混交林对土壤改良作用的研究有待进一步深入。
土壤粒度参数差异主要集中在0~20 cm,这与杜满义等[46]的研究结果一致。表层土壤中大量的植物凋落物、浅根系统及土壤生物体残留,可经微生物分解形成腐殖质,促进土壤养分的转化,增强土壤呼吸[47],进而影响土壤的粒度组成,本研究中土壤粒度参数随深度增加变化不显著,这与前人的研究结果基本一致[48],土壤粒度组成受到成土母质的强烈影响,会继承母质的一般特性[49],研究区的主要成土母质为第四纪黄土,土壤发育于同一母质,因此剖面粒度组成较为均一。此外,表层土壤由于受到植物根系影响在种植、耕作等作用下熟化,初步发育形成黄绵土[50],而深层土壤受植被影响较小,其粒度组成仍由黄土母质沉积过程中物源、搬运条件、沉积环境及沉积后的风化作用决定[51]。
3.3 粒级含量与分形维数的关系
相关性分析结果表明,分形维数与土壤黏粒含量呈极显著正相关,而不同植物群落分形维数与土壤粉粒、砂粒含量的相关性不同。这与董莉丽等[52]的研究结果不一致,可能是由于选取树种不同造成的,但也有研究表明分形维数与黏粒含量相关性最强,而与粉粒、砂粒相关性弱[53],由此可见对于分形维数与粒级含量间的关系仍需进一步研究。平均分形维数草地>油松纯林>山杏纯林>油松山杏混交林,本文中混交林分形维数小于纯林,直接原因在于黏粒含量少,混交林的垂直根系分布广泛可以改变土壤密度和土壤孔隙度增强土壤呼吸[54],土壤呼吸强度与气温在一定范围内呈正相关[55],进而对土壤粒度组成产生重要影响。
本研究表明不同植物群落土壤分形维数与粒级含量的拟合存在差异,这与前人的研究结果一致[56-57]。草地各粒级含量拟合的效果均优于人工林,因此,分形维数在不同植物群落中具有不同的适用性,且对草地描述效果更好。对于黏粒含量稀少的土壤使用分形维数,描述土壤粒径分布特征和结构时,分形维数的适用性应进一步研究。
3.4 土壤粒度频率曲线变化
研究区不同植物群落相似的土壤粒度频率曲线,源自于样地临近所导致的风沙搬运沉积的一致性[57]。0~200 cm土层,土壤粒度频率曲线的变化主要集中在Φ4.00~5.00 和Φ7.90~8.00,推测该区间为土壤细粒在垂直方向上的移动适宜区间。同时,不同植物群落土壤粒度频率曲线峰值均处于Φ4.00~5.00 区间范围内,且峰值大小表现为油松山杏混交林>油松纯林>山杏纯林>草地,表明混交林土壤颗粒分布更集中,土壤组成更均匀。此外,相关研究表明0~60 cm 是受植物影响较大的土层[58]。0~10 cm土层,不同植物群落的土壤粒度频率曲线差异最大,曲线在Φ7.96~10.00 区间范围内高度重合,而在其余区间范围存在显著差异。10~20 cm,土壤粒径分布差异主要集中在Φ3.35~5.19 和Φ5.56~8.69。20~60 cm,土壤粒径分布差异主要集中在Φ4.27~4.82、Φ6.11~8.32 和Φ9.79~11.27。综上,植物根系对Φ4.00~5.00、Φ6.00~8.00 和Φ10.00~12.00 区间的土壤颗粒影响更强,调节作用更显著。
4 结论
(1)研究区土壤属粉壤土,粉粒含量约占总体的56.58%~71.67%,砂粒含量21.37%~38.71%,黏粒含量占3.55%~6.96%;各植物群落土壤粒径分布呈极正偏度,峰态尖窄,分选性均较差,平均分形维数介于2.58~2.63,其中,草地土壤粒径分布特征优于人工林。
(2)研究区不同植物群落土壤粒度频率曲线呈相似分布,粒径分布差异主要集中在土壤表层(0~20 cm),随土壤深度增加,土壤粒度组成变化不显著。
(3)土壤分形维数与黏粒含量呈显著正相关,不同植物群落土壤分形维数与粒级含量的关系存在差异,分形维数与草地粒级含量相关性最高,表明分形理论可以更好地揭示草地土壤的粒径分布特征。
