松果菊F3'H基因的克隆及表达分析
2022-08-06吴红松秋田祐介
吴红松,秋田祐介
(1.日本埼玉工业大学 生命环境化学系,日本深谷 369-0293;2.菏泽学院 生物科学系,山东菏泽 274000)
松果菊是世界上最著名的药用植物之一,近年来因其观赏特性而备受关注。目前,针对松果菊的分子研究集中于遗传多样性,与花青素生物合成相关的基因研究鲜有报道。
类黄酮3'-羟化酶(Flavonoid 3'-Hydroxylase,F3'H),是花青素合成的关键基因,最初由BRUGLIERA等[1]从矮牵牛(Petunia hybrida)中分离并鉴定,目前已报道多种植物含有此基因。研究表明,F3'H负责B环3’位置的羟基化位置和程度,从而形成不同的花青素。因此,鉴定功能基因F3'H是了解松果菊中花青素羟基化和花色形成的重要步骤。研究发现松果菊F3'H(EpF3'H)的表达特点与菊花F3'H(CgF3'H)在菊花所有器官中的表达和百合F3'H(LvF3'H)在百合有色花瓣和无色花瓣中的表达结果相似[3-4]。EpF3'H的表达水平与花发育过程中花青素的形成和积累趋势基本一致,可能具有协同促进松果菊花瓣中花青素合成和积累的作用。但黄文坤等[5]发现F3'H在紫茎泽兰类花瓣中的表达低于在叶中的表达,可见F3'H基因在不同品种、组织和生长阶段具有不同的时空表达模式,它们的组织特异性表达可能受调控基因的影响
本文利用3'-RACE和5'-RACE法克隆了EpF3'H的cDNA,并分析了该基因在不同组织中的表达,以便开展针对松果菊花色的分子育种计划。
1 材料与方法
1.1 试剂与仪器
松果菊购自Sakata Seed Co.(日本),室外栽培。采摘时将松果菊开花阶段分为四个时期,分别命名为S1、S2、S3和S4:S1——花瓣闭合,绿色(长度小于15 mm);S2——花瓣开始着色(长约20 mm);S3——花瓣绯红,刚刚绽开(长约20 mm);S4——花瓣平展,花色紫红(长约30 mm)[2]。从不同开花阶段中收集花瓣和叶,实验至少重复三次。收集后立刻液氮预冷,在RNA提取前储存于-80 ℃。
十六烷基三甲基溴化铵(CTAB),日本Sigma;M-MLV逆转录酶,日本Takara;DNase I,日本Takara;3'-RACE和5'-RACE试剂盒,德国Roche。
T100型PCR仪,美国BIO;ABI 3730xl Analyzer型自动测序仪,美国Applied Biosystems;QuantStudio™1型RT-PCR仪,美国Thermo Fisher Scientific。
1.2 EpF3'H全长cDNA的克隆
从不同开花阶段(S1~S4)的花、叶中用CTAB法提取总RNA。2 μg RNA为模板,Oligo(dT) 21为引物,用M-MLV逆转录酶转录,DNase I处理,总体积20 μL,合成得EpF3'H的cDNA。使用3'-RACE和5'-RACE法扩增F3'H基因的全长cDNA片段。
设计一对引物EpF3'H-FP:5'-ATGACTATT CTAACCCTACTATCATACACC-3',EpF3'H-RP:5'-TTAACCACTTTCATATACTTGAGG-3'。PCR 参 数为:95 ℃预变性5 min,95 ℃、30 s,60 ℃、30 s,72 ℃、90 s,30个循环,最后72 ℃延伸5 min,分离、连接、转化、测序,获得全长cDNA片段。
1.3 系统发育分析
使用Genetyx-ver 12软件对不同植物F3’H蛋白的氨基酸序列进行多序列比对,Clustalw多重比对的相邻连接法构建系统进化树,bootstrap检验的重复次数为1 000次。
1.4 生物信息学分析
测序得到的EpF3'H基因的序列信息提交到NCBI,通过BLAST与核酸和蛋白质数据库进行比对搜索,利用NCBI查找开放阅读框(ORF),预测EpF3'H基因编码蛋白的理化性质。
1.5 表达分析
采用SYBR green荧光染料进行RT-PCR,每个PCR反应总体积20 μL,引物EpF3'H-FP1:5'-ACAGTGGAATGGGCAATAGC-3',EpF3'H-RP:5'-TTAACCACTTTCATATACTTGAGG-3'。采用 2-∆∆Cq计算各基因的相对表达量,以EpACT1作为内参基因,实验重复4次。
2 结果与分析
2.1 EpF3'H基因全长cDNA序列的合成
通过3'-RACE和5'-RACE扩增出cDNA的3'和5'端,从而获得EpF3'H的全长cDNA序列。ORF为1 533 bp,编码510个氨基酸(GenBank登录号OL828207),蛋白分子量为56.49 kD,pI为8.46。通过与NCBI上已登录的菊花、郁金香、葡萄等的F3'H蛋白序列进行比对,发现克隆的基因为F3'H基因。
2.2 EpF3'H的同源性和结构特征
从NCBI Blast的比对结果中选取来源于其他24种植物的F3'H蛋白与松果菊EpF3'H蛋白,用相邻连接法构建系统进化树(见图1)。从图1中可以看出,不同物种来源的F3'H蛋白在进化上归属不同的分支,EpF3'H与非洲菊杂交种、菊花的F3'H亲缘关系较近,处于同一分支上。蛋白质多序列比对分析显示,EpF3'H与非洲菊杂交种(DQ218417)、菊花(AB523844)高度同源,相似度分别为83.52%和82.67%,这表明EpF3'H是F3'H家族成员。F3'H可通过底物竞争羟基化二氢黄酮醇3'位,合成紫红色色素的前体物质,EpF3'H可能与其他F3'H蛋白一样具有相同的催化功能。
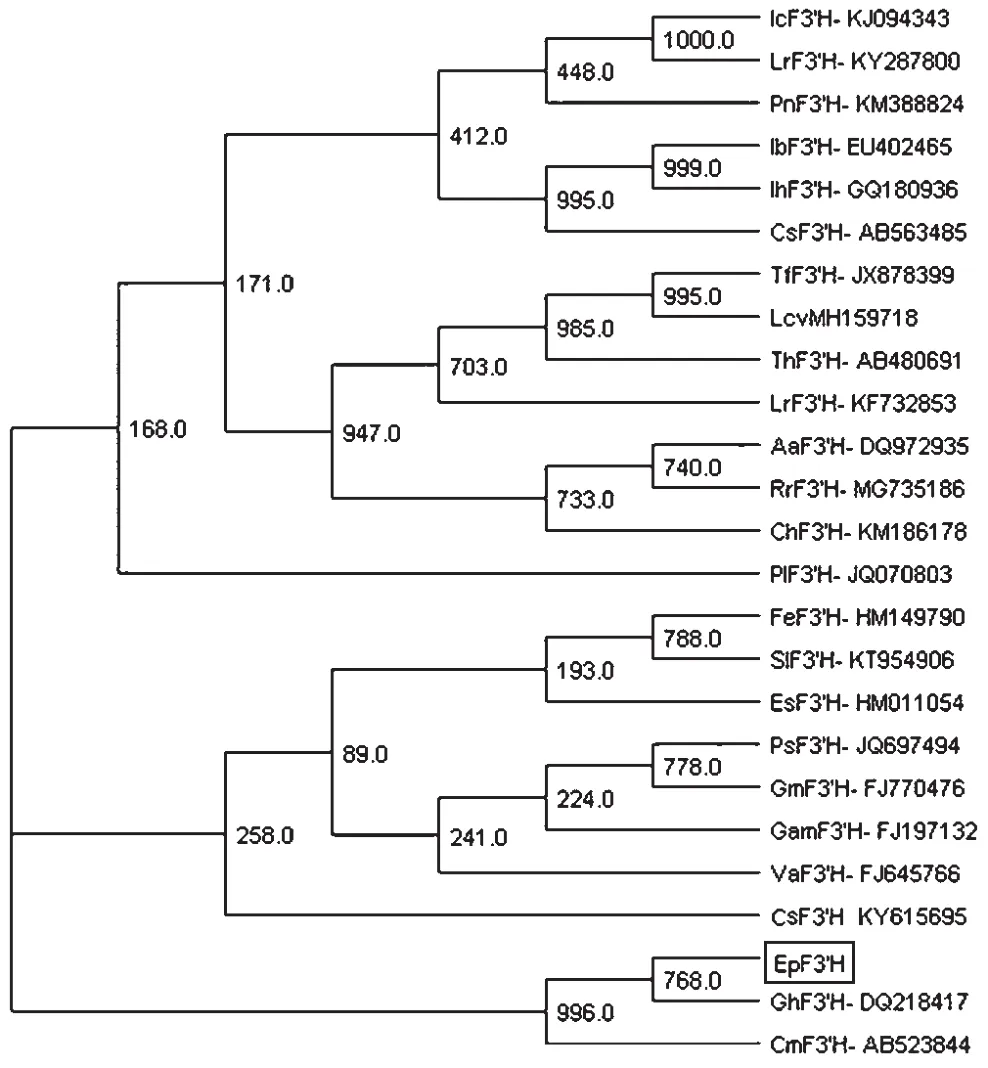
图1 EpF3'H与其他植物F3'H蛋白的系统进化树分析
通过NCBI保守基序的比对发现,EpF3'H蛋白功能结构域(细胞色素P450结构域)有四个特有保守基序和三个F3'H特有保守基序,表明EpF3'H是典型的F3'H蛋白。
2.3 EpF3'H的基因表达
RT-PCR结果(图2)显示,EpF3'H在松果菊的叶和花中均有不同程度的表达。EpF3'H在S4花期中表达量最高,其他时期表达量从高到低依次为S2、S1、S3和叶。可见,EpF3'H的表达模式大致与花青素积累模式相对应。
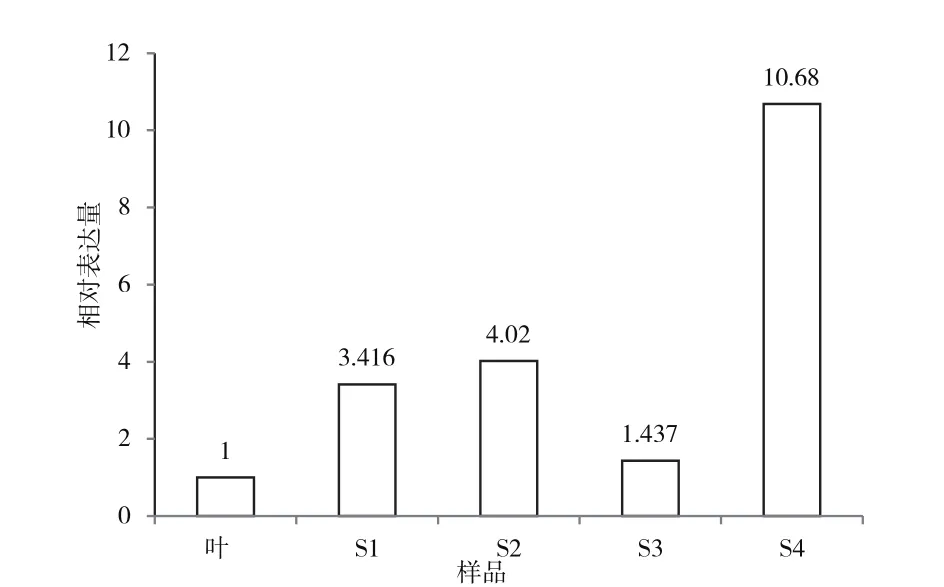
图2 EpF3'H基因相对表达量
3 结论
研究表明,EpF3'H参与了松果菊花色形成,也发挥了类似于F3'H在拟南芥等物种中的功能[6],可决定无色二氢黄酮醇的羟化位置和程度,从而合成不同种类花青素苷。
