不同基因型橡胶树花药体胚发生能力的差异分析
2022-08-03顾晓川彭素娜戴雪梅周权男孙小龙桂明春黄华孙华玉伟黄天带张源源
顾晓川,彭素娜,戴雪梅,周权男,孙小龙,桂明春,黄华孙,华玉伟,黄天带*,张源源
(1.中国热带农业科学院橡胶研究所,农业农村部橡胶树生物学与遗传资源利用重点实验室,海南 海口 571101;2.云南热带作物科学研究所,云南 景洪 572818)
天然橡胶是一种重要的工业原料和战略物资,其主要来源于橡胶树(Hevea brasiliensis (Willd.ex A.Juss.) Muell.Arg.)。目前,橡胶树生产上所应用的种苗主要为芽接苗,即实生苗作为砧木嫁接优良品种的接穗所培育的种苗。因砧木来源于自然授粉的种子,遗传基础不一致,导致芽接苗植株间产量、生长势等存在显著差异;而橡胶树体胚苗是由体细胞胚直接发育而来的种苗,具有自主发育的主根、无需嫁接在砧木上的幼态化无性系(又称橡胶树自根幼态无性系),较目前生产上应用的种苗-芽接树,产量高10%~30%,生长快10%~20%,是天然橡胶产业新一代种植材料[1-4]。
自1978年王泽云等成功移植成活橡胶树花药体胚苗开始[5],法国、马来西亚、印度尼西亚、斯里兰卡、印度等国投入大量的人力、物力、财力对橡胶树的体胚苗生产进行了广泛的研究,取得了一定的进展,但体胚诱导率和植株再生频率较低,生产成本较高,无法满足生产需求,成为制约体胚苗工厂化生产的瓶颈[6-8]。橡胶树雄蕊具有数量大、便于采集、容易表面灭菌处理、易于诱导愈伤组织等优点,是研发橡胶树体胚发生体系应用最多的外植体。研究发现,橡胶树雄蕊诱导的愈伤组织和体胚主要来源于花药壁体细胞[9-13],其愈伤组织和体胚再生植株的倍性同母本一致。以花药诱导的体胚作为外植体,诱导次生体胚循环增殖,可实现一次花药体胚发生进一步诱导多次体胚发生,且次生体胚植株再生频率高达86%以上,成为体胚大量增殖的新途径[14-16]。但目前仍有许多品种的花药难以诱导体胚形成,即使能够诱导体胚的品种,也仅有少数品种的花药外植体体胚诱导率能达到50%[17-23],制约了这一技术的广泛应用。
植物体胚发生能力显著性的受遗传变异影响,存在细胞核因子遗传及核质互作遗传影响现象,且存在遗传背景与培养基互作影响现象,通过分析体胚发生时的转录组表明,体胚发生能力差异悬殊的基因型间存在显著的差异基因表达。在珍珠粟再生能力遗传研究中发现,胚性愈伤组织数量更大比例表现为核质互作遗传影响,而再生频率主要表现为细胞核遗传影响[24]。在板栗体胚发生研究中发现,遗传背景与培养基对体胚发生效率存在着显著的互作影响,当用美国板栗体胚发生标准流程时,中国板栗不能诱导形成胚性组织,在‘中国板栗与美国板栗杂交后代’中,杂交F1代、F1代与美国板栗回交BC1代也不能诱导形成胚性组织,而BC1代与美国板栗回交BC2代、BC2代与美国板栗回交形成BC3代的自交BC3F3代能够诱导形成胚性组织,推断杂交后代中中国板栗基因的占比可能存在一个体胚发生阈值,超过该阈值的杂交后代不能诱导形成胚性组织;但当用欧洲板栗体胚发生标准流程时,中国板栗能够诱导形成胚性组织,而美国板栗以及中国板栗与美国板栗的杂交后代F1、BC1、BC2、BC3F3均不能诱导形成胚性组织,系统发育研究表明栗属起源于亚洲,首先向西迁移至欧洲,然后迁移至北美,因此,作者推断中国板栗与欧洲板栗的亲缘关系更近,所以,用欧洲板栗体胚发生流程能够诱导中国板栗形成胚性组织[25]。通过对不同体胚发生能力棉花基因型的转录组进行对比分析,表明不同基因型的胚性愈伤组织是高度同质的,而不同基因型的非胚性愈伤组织是高度异质的[26]。通过分析棉花高效胚胎发生基因型和顽拗胚胎发生基因型的早期体胚发生转录组,表明高效胚胎发生基因型的差异表达基因在脂肪酸、色氨酸和丙酮酸代谢中显著富集,而顽拗胚胎发生基因型的差异表达基因则在 DNA 构象变化中显著富集[27]。
橡胶树品种遗传基础狭窄,为通过遗传背景分组优化不同基因型体胚发生体系提供了条件。本研究将重点探讨橡胶树体胚发生能力在不同基因型之间的差异,利用前期试验中在多个品种体胚诱导率都较高的两种培养基,对32个橡胶树品种花药外植体的体胚发生进行研究,以期明确橡胶树不同品种花药体胚发生能力与遗传背景的相关性,为通过基因型分组优化不同品种的体胚发生体系提供依据,为突破橡胶树优良品种体胚苗大量增殖的瓶颈提供基础。
1 材料与方法
1.1 试验材料
以国家橡胶树种质资源圃(海南省儋州市宝岛新村)保存的32个橡胶树品种为材料(表1)。于春季3月份采集黄中带绿、未开放的雄花(花粉发育处于单核靠边期),用75%乙醇表面灭菌60 s后,立即用0.1% HgCl2处理10 min,最后用无菌水冲洗5次,每次3 min[14]。表面灭菌后的雄花用于后续愈伤组织诱导和体胚发生研究。
1.2 培养基成分
改良MS培养基用于愈伤组织诱导和体胚发育,其中,大量元素、微量元素、铁盐所需的药品和蔗糖购自广州化学试剂厂;有机营养、植物激素和生长调节剂采购于上海生物工程技术有限公司和Sigma-Aldrich公司;植物凝胶采购于Sigma-Aldrich公司。
花药愈伤组织诱导采用平底试管为培养容器。体胚分化阶段采用培养皿作为培养容器。
1.3 试验方法
1.3.1 系谱分析 参考《中国橡胶树育种五十年》并参考相关文献[28-30]信息,整理参试品种亲本并进行系谱分析。
1.3.2 花药愈伤组织和体胚诱导 首先将雄蕊从雄花中剥出,然后接种到愈伤组织诱导培养基上(M培养基和S培养基),每个品种每处理接种数量超过120粒花药,重复3次,26 ℃~28 ℃暗室中培养40 d~60 d。形成愈伤组织后,将花药愈伤组织接种到体胚发生培养基上(E培养基),于24~26 ℃暗室中培养40~60 d。M培养基为改良 MS 添加 4.6 μmol·L−1ZT,4.1 μmol·L−1Picloram,4.5 μmol·L−12,4-D。S 培养基为改良MS 添加 2.3 μmol·L−1KT,2.1 μmol·L−1Picloram,2.3 μmol·L−12,4-D。E 培养基为改良 MS 添加 2.2 μmol·L−16-BA,14.0 μmol·L−1KT,1.4 μmol·L−1GA3,0.1 μmol·L−1NAA,3.8 μmol·L−1ABA[15]。以上培养基均添加 204.5 mmol·L−1蔗糖,2.2 g·L−1Phytagel,灭菌前调整pH至5.8。
1.4 统计分析
愈伤组织形成率=形成愈伤组织的雄蕊数/接种雄蕊总数×100%。
体细胞胚形成率=形成体细胞胚的愈伤组织数/接种愈伤组织总数×100%。
子叶形体细胞胚形成率=形成子叶形体细胞胚的愈伤组织数/接种愈伤组织总数×100%。
试验结果采用SPSS的Ducan法进行差异性分析。
1.5 体胚苗倍性鉴定
体胚苗倍性鉴定用Sysmex Cy-Flow®Cube8流式细胞仪进行。鉴定方法参考仪器附送试剂使用方法(http://www.sysmex-partec.com),即取200 mg新鲜叶片置于塑料培养皿中,添加0.5 mL裂解缓冲液,用刀片快速切碎,30 μm孔径滤网过滤,添加1.6 mL DAPI染色体液,室温静置30 s后上机检测。每株体胚苗随机取1片叶片进行检测。流式数据使用机器自带软件作图。以已知二倍体和三倍体植株叶片混合检测作图为对照。
2 结果与分析
2.1 系谱分析
根据亲本来源,对国家橡胶树种质资源圃保存的32个橡胶树品种进行系谱分析,试验所用32个橡胶树品种分别属于初生代无性系、次生代无性系、三生代无性系和四生代无性系(图1)。PR107、GT1、PB86、Tjirl、海垦1、天任31-45、合口3-11这7个品种为初生代无性系,其余25个品种中的23个与以上7个初生代无性系中的1~3个存在亲缘关系,表明我国橡胶树栽培品种亲本来源狭窄。此外,图1系谱中,含初生代无性系遗传背景的后代中,以含PR107亲本的后代最多,共有14个,以PR107为父本的品种有GT1、93-114、RRIM513、PB86、RRIM600、海垦1共6个母本组。
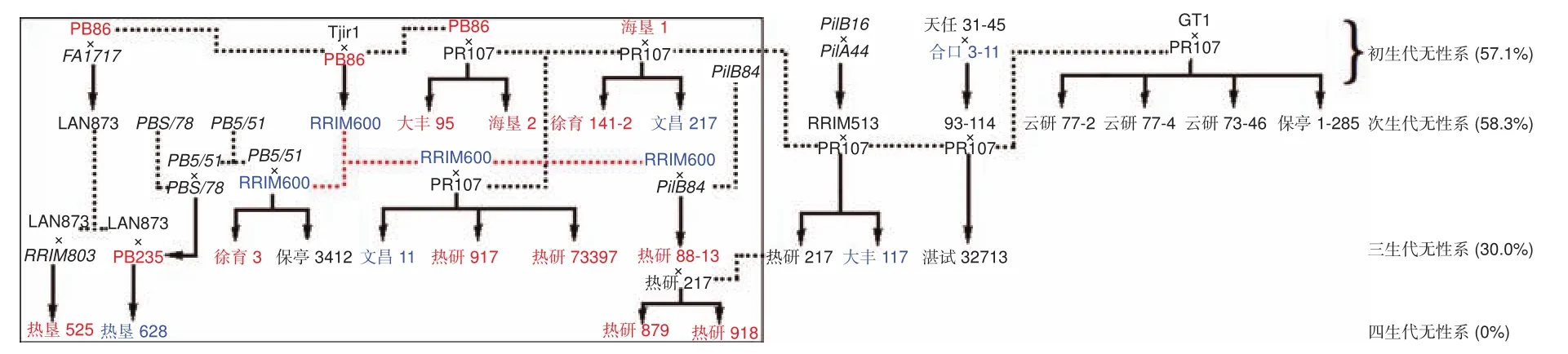
图1 橡胶树系谱和花药体胚发生能力差异分析Fig.1 Pedigree analysis of rubber tree and analysis of differences in anther embryogenic ability
2.2 花药愈伤组织和体细胞胚诱导
橡胶树花药愈伤组织、体细胞胚和体胚苗的诱导过程见图2。参试32个品种均能诱导出愈伤组织,多数品种在M培养基和S培养基上的愈伤组织诱导率接近,其中,31个品种的愈伤组织形成率均达50%以上(表1),表明所试愈伤组织诱导培养基在橡胶树花药愈伤组织诱导中具有广谱性。
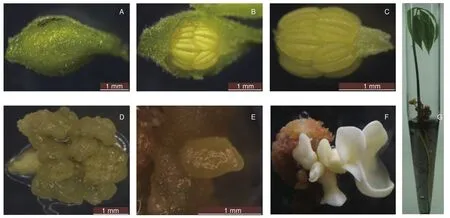
图2 橡胶树花药体胚苗的诱导过程图Fig.2 The Induction process of anther somatic embryo plantlet of rubber tree
愈伤组织转移到诱导体胚发生的E培养基后,来自M培养基和S培养基诱导的愈伤组织在大多数品种上体细胞胚诱导率较接近,只有个别品种存在显著性差异。23个品种能够诱导形成体细胞胚,且有14个品种能够诱导形成子叶形体细胞胚和体胚苗,其中,热研73397、热研88-13、热垦525和徐育3这4个品种的体细胞胚诱导率在50%以上,说明所试体胚发生培养基在橡胶树体胚发生中具有通用性,但仍有14个品种的体细胞胚诱导率在10%以下,且有9个品种未能诱导出体细胞胚(表1)。
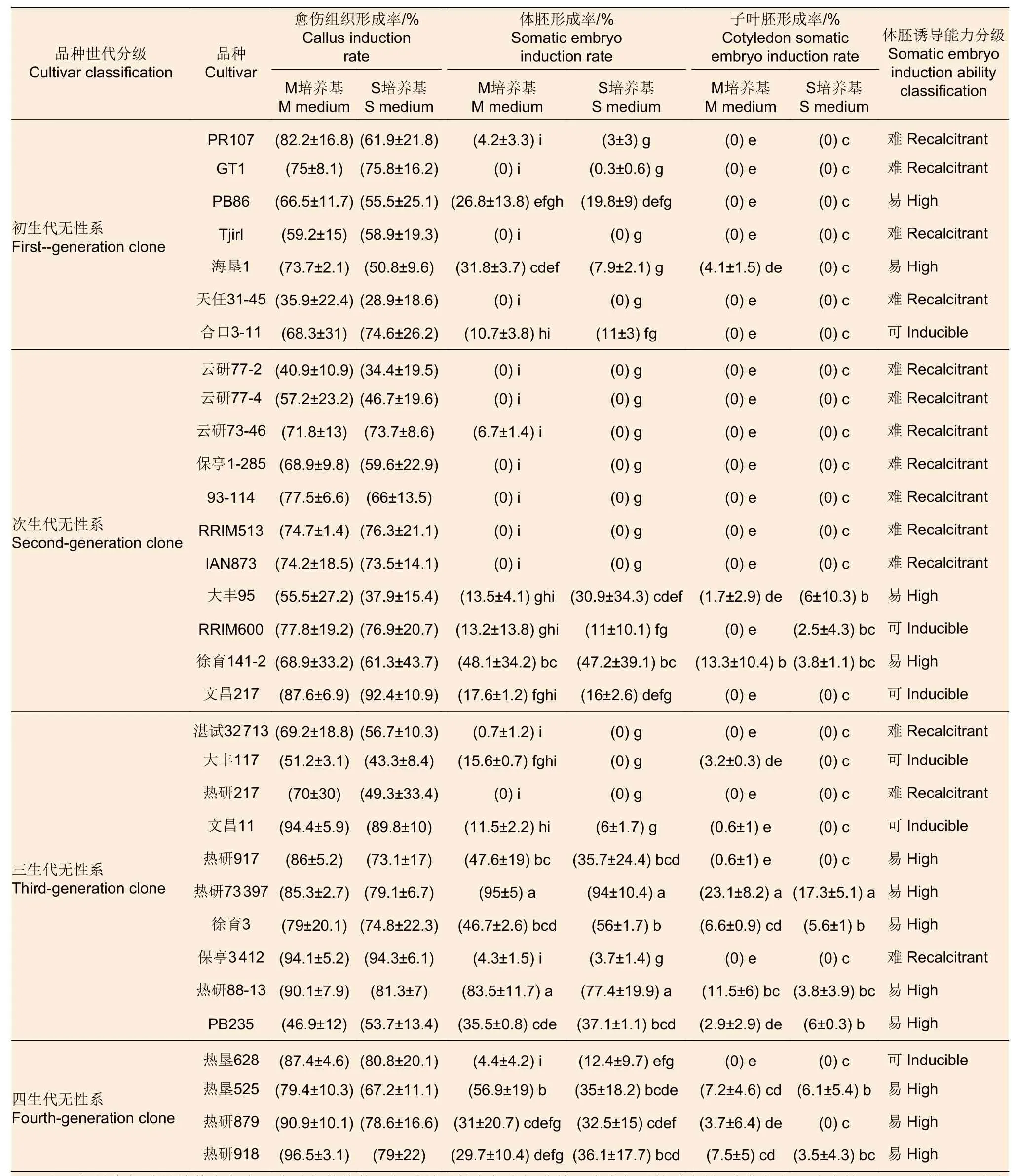
表1 不同橡胶树品种花药愈伤组织、体胚、子叶形体胚诱导率Table 1 Induction rate of anther callus,somatic embryo,cotyledon somatic embryo of different rubber tree cultivars
2.3 体胚诱导率与系谱关系
根据品种体胚诱导能力,将体胚诱导率<10%的品种定义为难诱导品种,体胚诱导率在≥10%且≤20%的品种定义为可诱导品种,体胚诱导率>20%的品种定义为易诱导品种(图1)。7个初生代无性系中,GT1、PR107、天任31-45、Tjirl属于难诱导品种,合口3-11属于可诱导品种,PB86和海垦1属易诱导品种。12个次生代无性系中,云研77-2、云研77-4、云研73-46、保亭1-285、93-114、RRIM513、IAN873属于难诱导品种,RRIM600、文昌217属于可诱导品种,大丰95、海垦2[5,17]、徐育141-2属于易诱导品种。10个三生代无性系中,湛试32713、热研217、保亭3412属于难诱导品种,大丰117、文昌11属于可诱导品种,PB235、徐育3、热研917(热研7-20-59)、热研73397、热研88-13属于易诱导品种。4个四生代无性系中,热垦628属于可诱导品种,热垦525、热研879、热研918(热研8-333)属于易诱导品种。分析发现,已知体胚诱导率的初生代、次生代、三生代和四生代无性系中难诱导体胚发生品种占比分别为57.1%、58.3%、30.0%、0.0%。
亲本诱导能力决定其子代品种体胚诱导能力。
2个亲本属于难诱导品种,其杂交后代多数为难诱导品种,如 GT1×PR107、93-114×PR107、RRIM513×PR107这3个杂交组合的后代有7个,6个为难诱导品种,只有1个为可诱导品种;1个亲本为难诱导品种,另1个为易或可诱导品种,其杂交后代多数属于易或可诱导品种,如PB86×PR107、Tjir1×PB86、海垦 1×PR107、天任 31-45×合口 3-11、RRIM600×PR107、IAN873×PB235、热研88-13×热研217这7个杂交组合的后代有12个,其中,7个品种属于易诱导品种,4个品种为可诱导品种,只有1个品种为难诱导品种。
具有相同亲本的橡胶树品种体细胞胚诱导率差异显著,且存在超亲现象。RRIM600×PR107子代热研73397、热研917和文昌11三个品种中,热研73397的体胚诱导率和子叶形体胚诱导率分别为95.0±5%、23.1±8.2%,显著性高于热研917(47.6±19%、0.6±1%)、文昌 11(11.5±2.2%、0.6±1%)和双亲;海垦1×PR107子代徐育141-2和文昌217中,徐育141-2的体胚诱导率和子叶形体胚诱导率(48.1±34.2%、13.3±10.4%)显著性高于文昌217(17.6±1.2%、0)和双亲;RRIM513×PR107子代热研217和大丰117中,热研217的体胚诱导率为零,而大丰117在M培养基上体胚诱导率达到15.6±0.7%,且显著性高于双亲;GT1×PR107子代云研77-2、云研77-4、云研73-46和保亭1-285四个品种中,只有云研73-46在所试培养基上能够诱导得到体胚。
2.4 体胚苗倍性鉴定
流式细胞仪倍性检测结果(图3和表2)表明:混合已知二倍体和三倍体植株叶片DNA相对含量在200和300位置均有波峰作为对照(图3A),二倍体的DNA相对含量为200(图3B),三倍体植株为300(图3C),流式细胞仪倍性检测可以准确鉴定出橡胶树体胚苗的倍性。
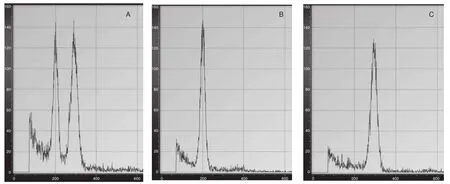
图3 流式细胞倍性检测Fig.3 Flow cytometric analysis
橡胶树不同基因型体胚苗倍性鉴定统计结果见表2,从15个品种共552株体胚苗倍性鉴定结果看,不同基因型体胚苗同其母本倍性一致,证明多基因型花药体胚苗来源于体细胞。
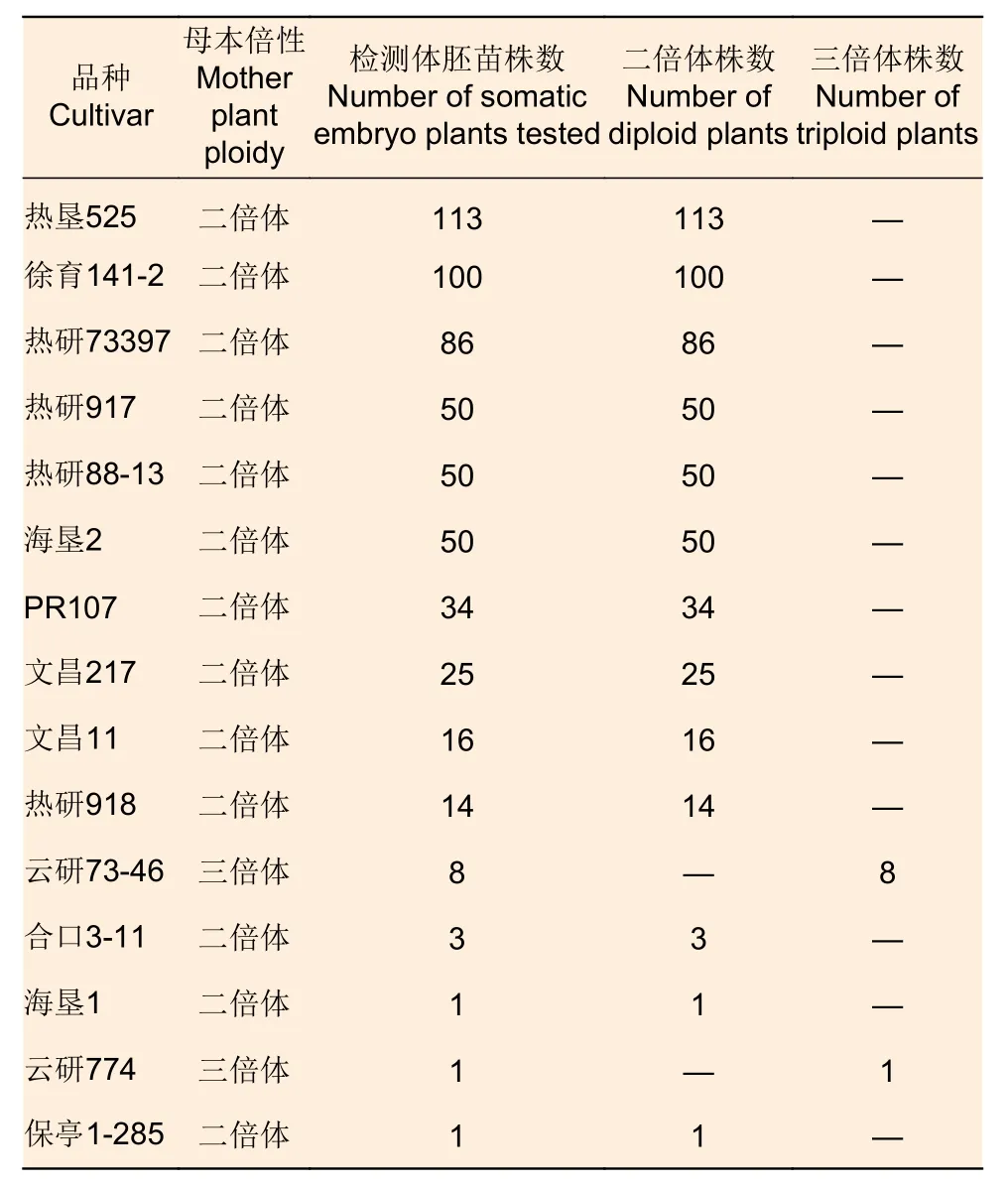
表2 橡胶树不同基因型体胚苗倍性检测结果Table 2 Ploidy test results of somatic embryo plants of different genotypes of rubber tree
3 讨论
3.1 本研究橡胶树品种体胚诱导能力与已报道研究结果一致,部分品种体胚诱导率显著性高于已有研究报道。
目前,未有深入开展橡胶树多基因型体胚发生能力研究的报道,在检索的少量研究报道中,橡胶树品种热研879花药体细胞胚诱导率为10%~14%,RRIM600为11%~49%,热垦525为51%左右,热研73397为21%左右,都属于易或可诱导体细胞胚品种;云研77-2、云研77-4、云研73-46、GT1和PR107花药体细胞胚诱导率都在10%以下,甚至为0,都属于难诱导体细胞胚品种,与本研究结果一致[17-23]。本研究的32个橡胶树品种中,有18个品种的体细胞胚诱导率在10%以上,有14个品种能够诱导得到子叶形体细胞胚,其中,热研73397的体细胞胚诱导率最高达95%,热研879的最高达32%以上,显著高于前人的研究结果。
3.2 基因型是决定植物体细胞胚发生能力的关键因素,并且这种能力可遗传。
Hodges等发现,在相同的培养条件下,玉米A188自交系的体细胞再生能力显著高于其他品系,以其作为亲本与无再生能力的自交系进行杂交,杂交后代具有再生能力;另外以两个再生能力差的自交系进行杂交,杂交后代再生能力也差,表现出不同基因型玉米的体细胞再生能力有较大差异且可遗传[31]。Parrott等发现,基因型对大豆体细胞胚发生能力有很大影响,有良好再生能力的品系在其系谱中都有一个或两个高再生能力的祖先亲本,且高再生能力基因型与低再生能力基因型的杂交F1代表现出中等再生能力,表明大豆体细胞再生能力可遗传[32]。Niskanen等发现,父本和母本的基因型对欧洲赤松体细胞胚发生都有影响,在胚性组织起始诱导阶段,母本的影响大于父本,在胚性组织长期继代增殖阶段,父本的影响相对母本增加[33]。在遗传变异对甜根子草胚性愈伤组织诱导和再生能力研究中发现,愈伤组织和胚性愈伤组织的诱导能力受遗传变异显著影响,而胚性愈伤组织的再生频率不受遗传变异影响,愈伤组织诱导能力与胚性愈伤组织诱导能力及再生频率密切相关,表明不同基因型的胚性愈伤组织诱导和再生频率能够基于愈伤组织诱导能力进行预测[34]。在橡胶树体细胞胚发生方面,前人开展了部分品种的花药体细胞胚发生研究,发现不同品种间的体细胞胚发生能力存在显著差异[17-23]。本研究通过对32个橡胶树品种的花药愈伤组织和体细胞胚诱导情况进行分析,发现橡胶树愈伤组织诱导能力不存在明显的基因型影响,而体细胞胚发生能力存在明显的基因型影响;对于橡胶树不同品种来说,子代的体细胞胚形成率与亲本体细胞胚发生能力相关,父本和母本均影响子代体细胞胚发生能力,至少含一个易于诱导或可诱导亲本,子代才易于或可诱导体胚,表明橡胶树体细胞胚发生能力可遗传。另外本研究发现,已知体胚诱导率的初生代、次生代、三生代和四生代无性系中难诱导体胚发生品种占比依次减少,推测随着易诱导亲本与难诱导亲本的杂交,后代中易诱导体胚发生品种有逐渐增多的趋势。
3.3 植物体细胞再生能力遗传复杂,表现为由多基因控制;可通过改进培养条件、连续再生驯化和过表达再生基因等途径提高体细胞再生能力、减少基因型依赖。
Gawel等将能产生体细胞胚的棉花品种与不能产生体细胞胚的品种进行杂交,分析了后代的体细胞胚发生能力,统计结果表明棉花体细胞胚发生是由多基因参与控制的[35]。叶兴国等选用13个小麦亲本基因型及其F1代进行花药培养,发现小麦花药培养力的遗传控制非常复杂,表现为多基因控制的数量性状遗传[36]。何平等用同一种培养基培养籼粳杂交F1代群体110个株系的花药,结果表明水稻花药培养力是由多基因控制的[37]。张晓玲等对来源于美国、墨西哥和中国的144份不同玉米自交系幼胚胚性愈伤组织的再生能力相关性状进行研究,结果表明玉米幼胚培养性状的遗传变异受多基因调控[38]。本研究发现,具有相同亲本的橡胶树品种体细胞胚诱导率差异显著,且存在超亲现象,表明橡胶树体细胞胚发生能力的遗传控制复杂,推测橡胶树体细胞胚发生也是由多基因参与控制的。
Hargreaves等改进了胚性细胞系的启动和早期增殖方法,在20个控制授粉的辐射松合子胚培养中得到成功应用,显著好于以前结果,表明有可能通过改进培养条件,打破遗传背景的影响[39]。李健英等经过连续再生驯化(SRA)的方法提高了棉花再生和转化效率,并在非胚性愈伤组织时期外施DNA甲基化抑制剂促进了体细胞胚发生[40]。王轲等通过在小麦中过表达再生基因TaWOX5,显著提高了小麦组织培养再生植株和转化效率,促进了质量较差愈伤组织的分化,基本克服了基因型依赖,而且未出现过表达WUS2和BBM等再生基因时出现的影响愈伤组织分化和根系生长或导致畸形植株的弊端,同时该基因在大麦、小黑麦、黑麦、玉米中的再生和转化作用也得到了验证[41]。本研究中多个橡胶树品种的体细胞胚诱导率显著高于前人的研究结果,推断得益于培养基改进。
3.4 结合体胚诱导率,通过基因型分组优化橡胶树品种体胚发生条件和在育种上应用
在目前栽培的主要橡胶树品种中,含PB86、RRIM600、热研88-13和海垦1等亲本的后代基本为容易或可诱导体胚的品种类型;含GT1、天任31-45、93-114和RRIM513等亲本的后代体胚诱导率较低;含PR107亲本的后代体胚诱导难易均有,当其后代亲本中不含易或可诱导亲本遗传基础时,后代体胚诱导率低,如母本中含GT1、93-114和RRIM513等亲本的PR107后代体胚诱导率低,当其后代亲本中含有易或可诱导亲本时,后代体胚诱导率高,如母本中含PB86、RRIM600和海垦1等亲本的PR107后代基本为容易或可诱导体胚的品种类型。本研究结果为今后橡胶树体胚发生培养体系的优化指明了方向,可以按亲本进行分组,进一步优化体胚培养条件,如含PB86、RRIM600、热研88-13和海垦1等遗传基础的后代可在现有培养条件的基础上进一步优化,得到适宜培养条件,如含GT1、天任31-45、93-114和RRIM513等遗传基础的后代需在现有培养条件基础上进行大幅度的调整和优化,如从外植体选择与处理、基本培养基筛选、新药品探索、连续再生驯化、过表达再生基因等更多方向开展试验。同时本研究结果可以为橡胶树的传统杂交育种提供参考,以性状优良且体胚诱导率高的橡胶树品种为亲本配置杂交组合,获得高体胚诱导率的优良子代几率大,也为高产高抗无性系优质种苗的工厂化生产和转基因育种研究提供了基础。
4 结论
本研究通过对32个橡胶树品种的遗传关系和体胚发生能力进行比较分析,发现体胚诱导率存在显著的基因型差异,且这种差异可遗传。依据体胚诱导率和遗传关系,将亲本的诱导率同子代的诱导率结合进行分析并分组,为通过基因型分组优化橡胶树多品种体胚发生提供了依据。结果表明,亲本PB86、RRIM600、热研88-13和海垦1及其后代可按本研究所采用的培养条件进行小幅度优化,得到更适合的培养条件;亲本GT1、天任31-45、93-114和RRIM513及其后代需要在现有培养条件基础上进行大幅度的优化,以明确其培养条件。本研究中多个橡胶树品种的体细胞胚诱导率显著高于前人的研究结果。另外,本研究通过对15个品种的体胚苗进行倍性鉴定,证明不同基因型花药体胚苗均来源于体细胞。
