过量铜在两个柑橘品种幼苗中的分布特征及根细胞壁响应机制
2022-07-18李欣钰林妹兰胡萍萍陈立松黄增荣
李欣钰,林妹兰,卢 飞,胡萍萍,陈立松,黄增荣
(福建农林大学资源与环境学院,福建福州 350002)
铜是高等植物生长发育必需的微量营养元素之一,参与了光合电子传递,抗氧化保护以及细胞壁修饰等生化反应[1]。在柑橘生产中,含铜菌剂常用于叶片喷施防治柑橘疮痂病和黑斑病等[2]。连年喷施造成铜在柑橘园土壤中逐年累积[3–4]。土壤中的铜主要以正二价(Cu2+)形态存在。相比其他二价金属元素,土壤有机质对Cu2+的结合能力最强[5]。土壤有机质对铜的强烈吸附减少了铜向柑橘树体的迁移。但据本课题组对福建平和300多个琯溪蜜柚果园土壤和叶片的调查,仍有70.3%的叶片铜含量超过适宜浓度[6]。铜超标蜜柚叶片占比高,一方面是由于在柑橘生产中过多施用含铜杀菌剂(如波尔多液、蓝矾等)以及含铜微量元素肥料,导致喷落的铜逐年富集在果园土壤表层[7–8];另一方面是由于果农在蜜柚园养分管理中偏施化肥,少施有机肥,造成果园土壤逐年酸化(300多个琯溪蜜柚果园土壤pH平均值为4.34,且90.0%的土壤样品pH小于5.0[6]),促进土壤中吸附性铜的解吸,提高土壤溶液中铜的活性。活化的铜通过根向柑橘体内迁移,增大柑橘铜中毒风险[9]。因此,研究酸性条件下柑橘苗对过量铜的响应机制对于优化果园铜养分管理、防控柑橘园铜毒害、提高柑橘产量和品质具有重要意义。
铜的迁移与再分布是植物应答铜毒害最重要的机制之一[10]。在组织水平上,大量铜富集在多数植物的根中。铜在根中的固定直接减少了其向地上部的迁移。在细胞水平上,植物可通过“拒铜”或“区隔化”抵抗铜毒害。例如,植物可通过根系分泌的有机酸[11–12]或通过质子泵的外排作用[13]将铜拒之根表。对于进入细胞质的铜,可通过铜转运蛋白将其转运到液泡中,以降低对同化器官的破坏[14]。在亚细胞水平上,有关铜的主要分布部位目前仍存在争议。如Mwamba 等[15]研究表明,过量铜主要结合在油菜根的可溶性成分。Wan等[16]研究表明,过量铜主要富集在苹果根细胞壁上。由于植物细胞可溶性成分复杂,近年来有关植物对铜区隔与固定的研究主要关注植物根细胞壁的多糖组分。植物根细胞壁主要由纤维素、半纤维素和果胶等多糖组分构成[17–18]。各多糖组分富含带有负电荷的羟基和羧基等,可通过配位吸附固定铜。目前,有关过量重金属在细胞壁上的主要结合位点有不同的报道。如Ren等[19]研究表明,蓖麻根细胞壁的半纤维素组分是铜的主要结合位点。Yang等[20]研究表明,甘蔗根细胞壁果胶组分是锰的主要累积部位。
根细胞壁对铜的固定与植物耐铜性相关。研究表明,过量铜主要固定于苹果须根上,在须根亚细胞水平铜分布特征表现为细胞壁>质体>可溶性部分>线粒体>细胞核,且耐铜品种须根细胞壁铜累积量显著高于铜敏感品种[21]。进一步研究表明,根细胞壁对铜的固定与其微结构变化有关[22–23]。例如,经200 μmol/L铜处理30天的毛竹根铜浓度可达810 mg/kg,而转移至地上部的铜浓度仅为91.0 mg/kg,且过量铜处理的细胞壁可见明显扭曲变形与增厚[24]。根细胞壁增厚现象在受铜毒害影响的牛至[25]和蓖麻幼苗中皆有报道[26–27]。FTIR分析显示,大部分铜结合到细胞壁纤维素、半纤维素、果胶和蛋白质的羧基、羟基和氨基甲基等官能团上[28–29]。Krzesłowska[23]的研究结果显示,含羧基的多糖数量决定了细胞壁结合金属离子的能力。张晓斌等[30]的研究表明,耐重金属植物根细胞壁相应基团表现出不同程度的位移或吸光度的差异。目前,关于过量铜在柑橘树体中的迁移与微分布以及过量铜诱导的根细胞壁微结构变化的报道较少。
本研究以具有耐铜差异的雪柑和沙田柚实生苗[31]为试验材料,设置连续18周含铜浓度分别为0.5 μmol/L (对照)和 300 μmol/L (铜过量)的铜处理,研究过量铜在雪柑和沙田柚实生苗中的分布特征以及柑橘根细胞壁微观结构变化,旨在进一步解析柑橘过量铜应答机制,为优化柑橘铜养分管理,实现高产优质栽培提供理论和实践依据。
1 材料与方法
1.1 试验材料与处理
挑选饱满的沙田柚和雪柑种子,播于含有干净河沙的黑色穴盘中。出芽5周后,将长势一致的两个柑橘品种实生苗移栽到含有干净河沙的陶瓷花盆中,沙田柚和雪柑各24盆,每盆2株,在福建农林大学金山校区的温室大棚内自然光下生长,每两天浇一次营养液,每盆500 mL。营养液配方:KNO32.5 mmol/L、Ca(NO3)22.5 mmol/L、KH2PO40.5 mmol/L、MgSO41 mmol/L、H3BO310 μmol/L、MnCl22 μmol/L、ZnSO42 μmol/L、CuCl20.5 μmol/L、(NH4)6Mo7O240.065 μmol/L 和 Fe-EDTA 20 μmol/L。
移栽6周后,对柑橘实生苗进行不同浓度铜处理,铜浓度分别为 0.5 μmol/L (对照,Cu0.5)和 300 μmol/L (铜过量,Cu300),营养液 pH 用 1 mol/L HCl或NaOH调至4.30~4.50。处理18周后,柑橘苗按主根、侧根、茎和叶分类采集。其中,柑橘根样品用自来水洗净沙粒后用预冷的0.5 mol/L EDTANa2浸泡10 min再用蒸馏水清洗3次,经吸水纸吸干后采集。取部分柑橘苗侧根样品立刻放入固定液中用于透射电镜观察;另取部分柑橘苗侧根鲜样用液氮固定,保存于–80℃冰箱,用于亚细胞组分分离和铜含量测定。剩余侧根和主根鲜样放入70℃烘箱烘干后,用于粗细胞壁的提取及铜含量的测定。柑橘地上部样品用蒸馏水清洗3次,表面干燥后放入70℃烘箱烘干至恒重用于铜含量的测定。
1.2 试验方法
1.2.1 柑橘幼苗主根、侧根、茎和叶中铜含量的测定
烘干至恒重的柑橘幼苗主根、侧根、茎和叶粉末,经浓硝酸–浓高氯酸(V∶V=5∶1)过夜消化后,用原子吸收光谱法 (PerkinElmer Singapore Pte Let.,Singapore)测定雪柑和沙田柚主根、侧根、茎和叶中的铜含量[32],每个样品4个重复。
1.2.2 亚细胞组分的分离与铜含量的测定 参考周丽珍等[33]的方法分离亚细胞组分。称取适量柑橘根尖新鲜样品于预冷的匀浆液(50 mmol/L顺丁烯二酸盐缓冲液,pH 7.8;250 mmol/L 蔗糖;1 mmol/L 二硫赤藓糖醇)中,研磨成匀浆,转入50 mL离心管;于 4℃ 高速冷冻离心机 (Thermo Scientific Sorvall Legend Micro 17R,USA)中,以 3000 r/min 离心1 min,底部沉淀为细胞壁组分;上清液转入新的离心管后,以 15000 r/min 离心 45 min,底层为细胞器组分;上清液转入新的离心管,为可溶性部分(含细胞质及液泡),所得所有组分均于4℃保存。分离得到的亚细胞组分细胞壁、细胞器及可溶性部分,经浓硝酸–浓高氯酸(V∶V=5∶1)消化后,利用NexION 300XICP-MS (PerkinElmer,Shelton,CT,USA)测定3个组分的铜含量,每个样品含3个重复。采用柑橘叶国家标准物质GBW10020 (GSB–11)进行分析质量控制。其中,柑橘叶标准品铜含量为6.6±0.5(μg/g, DW),质控结果为 6.83±0.48 (μg/g, DW)。
1.2.3 细胞壁的提取、多糖组分的分离与铜含量测定
参考Zhong等[34]的方法,提取细胞壁并分离细胞壁多糖组分。称取适量烘干并磨碎成粉末的柑橘侧根样品于离心管中,采用75%乙醇、丙酮、甲醇–氯仿(V∶V=1∶1)和甲醇4种提取剂(固液比为0.1 g∶10 mL)依次分别提取。加入提取剂充分混匀后,静置 20 min,在 12000 r/min 的离心机中离心 10 min,弃去上清液,并加入去离子水清洗,混匀并离心,弃去上清液,得到粗细胞壁。含粗细胞壁的离心管覆盖单层保鲜膜并扎孔,于55 ℃烘箱烘干至恒重,称量粗细胞壁的质量,计算细胞壁提取率。称取0.01 g干燥的细胞壁样品于离心管中,加入2 mL 0.5%草酸铵 (含 0.1% NaBH4,pH=4.00),沸水浴 1 h 后12000 r/min离心 10 min,取上清液于新的离心管中,重复3次合并上清液得到果胶组分。沉淀加入4% KOH (含 0.1% NaBH4),并于室温下在 200 r/min的摇床振荡 8 h 后 12000 r/min 离心 10 min,取上清液于新的离心管中,重复3次提取(共24 h,每次需8 h),合并上清液,上清液即为半纤维素Ⅰ类。沉淀加 24% KOH (含 0.1% NaBH4)后重复上步提取操作得到半纤维素Ⅱ类组分。最终沉淀为纤维素,烘干至恒重并称重。得到的不同细胞壁多糖组分保存于4℃冰箱备用。分离得到的3种细胞壁多糖组分和纤维素,经浓硝酸–浓高氯酸(V∶V = 5∶1)消化后,用ICP-MS测定各组分中的铜含量,每个样品5个重复。
1.2.4 侧根透射电镜的观察 参考Huang等[35]的方法,处理柑橘侧根根尖鲜样。柑橘侧根在装有3%戊二醛–1.5%多聚甲醛固定液的注射器中抽真空,并于3%戊二醛–1.5%多聚甲醛的固定液中4℃预固定3 h,用 0.1 mol/L 磷酸缓冲液 (pH 7.2)清洗 3 次,每次间隔15 min,在1%锇酸–1.5%亚铁氰化钾的固定液中固定2 h,用蒸馏水清洗3次,每次15 min,最后用30%、50%、70%、80%、90%、95%、100%的乙醇进行梯度脱水,每级15 min,其中100%酒精洗脱3次,加丙酮清洗2次,每次间隔10 min,加树脂–丙酮(V∶V=1∶1)的混合液置换1次,在摇床上摇2 h,加树脂–丙酮(V∶V=3∶1)的混合液渗透1次,在摇床上摇2 h,取出,在滤纸上吸去残余丙酮,加全树脂过夜。取出样品,将单个样品分开放入含全树脂的PCR小管中,用环氧树脂618包埋,并做好标记,凝固后用超薄切片机切片,在日立HT7700透射电镜下观察并拍照,每个样品4个重复。
1.2.5 FTIR分析 参考Liu等[36]的方法,称取约2 mg干燥细胞壁与200 mg KBr放入玛瑙研钵中研磨混合均匀后倒入压模中,在真空条件下得样品薄片,放入 FTIR 采样器 (Thermo Scientific Nicolet iS50,USA)测定红外光谱,在 4000~400 cm–1光谱范围内,分辨率为4 cm–1,扫描累加次数32次,以各吸收峰的位置和吸收率为指标,构建数据库,即测得各组样品的原始红外光谱数据。在OMNIC 8.0软件上进行自动基线校正和平滑。为了提高谱图的分辨率,以便减少谱图叠加,对不同处理选择含有多糖、酰胺和酯等特征信息的1800~800 cm−1光谱范围进行精细表征,每个样品3个重复。
1.3 统计分析
采用完全随机进行试验设计,4个不同处理组合间采用 2 (柑橘品种)×2 (铜处理水平)双因素方差分析进行差异性检测,利用SPSS 16.0软件(Chicago,USA)进行统计分析,统计结果表示为平均数±标准误差。利用 Image-Pro Plus 6.0 (Media Cybernetics,USA)软件处理透射电镜照片,并使用Origin 2018软件(OriginLab,USA)制图。
2 结果与分析
2.1 过量铜在两个柑橘品种幼苗主根、侧根、茎和叶中的分布
如表1所示,在柑橘幼苗的组织水平上,不论是对照 (Cu 0.5 μmol/L)还是过量铜处理 (Cu 300 μmol/L),侧根中的铜含量均最高。以含量最低的柑橘主根铜含量(雪柑和沙田柚分别为4.46和2.34 μg/g, DW)作为标准(数值为1),对照组雪柑和沙田柚主根、侧根、茎和叶的铜含量相对值分别为 1、6.16、2.02、1.49 和 1、8.06、3.37、3.23。相比对照,300 μmol/L铜处理显著提高了两个柑橘品种幼苗各部位的铜含量。以含量最低的柑橘叶铜含量(雪柑和沙田柚分别为29.21和34.86 μg/g, DW)作为标准(数值为1),300 μmol/L铜处理的雪柑和沙田柚主根、侧根、茎和叶的铜浓度相对值分别为16.3、190、1.18、1 和 14.7、194、1.26、1。对照组的雪柑和沙田柚侧根铜分布率分别为57.32%和51.31%。300 μmol/L 铜处理的雪柑和沙田柚侧根铜分布率分别提高到90.87%和91.99%。但过量铜处理降低了铜在柑橘茎和叶的分布率。品种间比较结果显示,铜处理下沙田柚侧根铜含量显著高于雪柑。
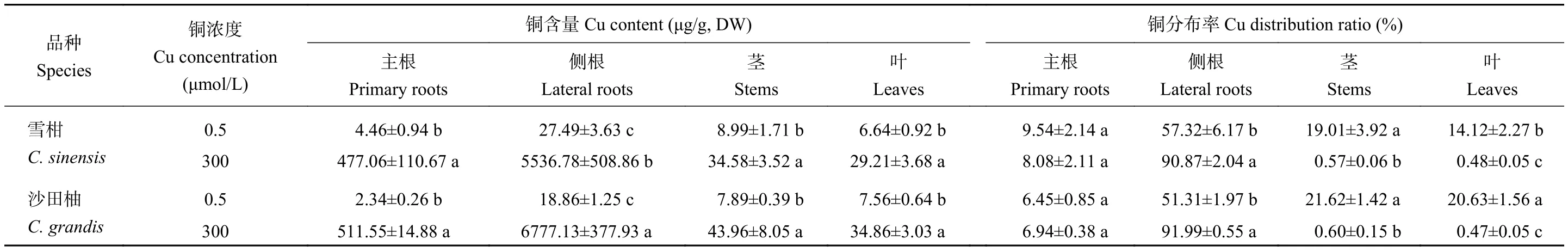
表1 不同铜处理的两个柑橘品种幼苗主根、侧根、茎和叶中的铜含量与分布率Table 1 Cu content and distribution ratio in the primary roots, lateral roots, stems and leaves of two citrus species under different Cu concentrations
2.2 过量铜在两个柑橘品种幼苗侧根细胞壁、细胞器和可溶性部分的分布
如表2所示,对照组中的柑橘幼苗侧根细胞器铜浓度最高,细胞壁中铜浓度最低。对照组的雪柑和沙田柚侧根细胞器铜分布率分别为58.61%和39.26%;300 μmol/L铜处理显著提高了两个柑橘品种幼苗侧根各亚细胞组分中的铜含量。过量铜处理的柑橘幼苗侧根亚细胞水平上可溶性部分铜浓度最高,细胞器浓度最低,过量铜处理的雪柑和沙田柚侧根细胞器的铜分布率分别为14.34%和11.49%。相比对照,过量铜处理提高了细胞壁和可溶性部分的铜分布率,但显著降低了细胞器的铜分布率。过量铜处理的沙田柚细胞壁中的铜含量与分布率均显著高于雪柑。

表2 不同铜处理的两个柑橘品种幼苗侧根细胞壁、细胞器和可溶性部分的铜含量与铜分布率Table 2 Cu content and distribution ratio in the cell wall, organelles and soluble fractions of lateral roots of two citrus species under different Cu concentrations
2.3 过量铜在两个柑橘品种幼苗侧根细胞壁果胶、半纤维素Ⅰ、半纤维素Ⅱ和纤维素中的分布
由表3可知,对照和过量铜处理,铜在细胞壁多糖组分中的分布均为果胶>纤维素>半纤维素Ⅰ>半纤维素Ⅱ。与对照相比,过量铜处理柑橘侧根细胞壁多糖组分中的铜含量均显著提高,果胶铜含量上升尤为显著。对照组的雪柑和沙田柚侧根细胞壁果胶铜分布率分别为44.69%和37.15%;过量铜处理后,雪柑和沙田柚侧根铜分布率分别提高到98.45%和98.48%。过量铜处理的雪柑和沙田柚侧根细胞壁果胶铜含量较对照分别提高了170和290倍。品种间比较发现,过量铜处理后的沙田柚侧根细胞壁果胶铜含量显著高于雪柑。从铜在柑橘侧根细胞壁多糖组分中的分布率来看,过量铜处理的半纤维素Ⅰ、半纤维素Ⅱ和纤维素中的铜分布率显著降低,而果胶铜分布率显著提高。过量铜处理的两个柑橘品种侧根细胞壁果胶铜分布率均高于98%。
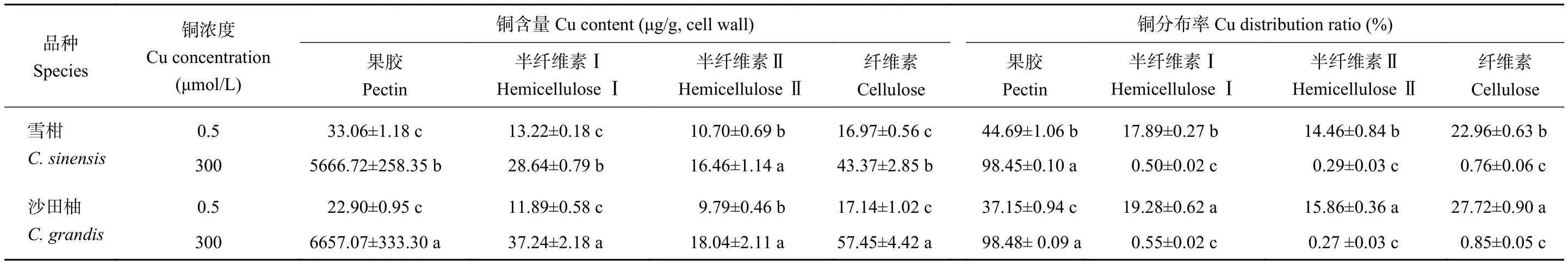
表3 不同铜处理的两个柑橘品种幼苗侧根细胞壁多糖组分中铜含量与铜分布率Table 3 Cu content and distribution ratio in polysaccharide compositions of lateral roots cell wall of two citrus species under different Cu concentrations
2.4 过量铜对两个柑橘品种幼苗侧根细胞壁的影响
相比对照,过量铜处理显著提高了两个柑橘品种幼苗的细胞壁提取率,且过量铜处理的沙田柚细胞壁提取率显著高于雪柑(图1)。透射电镜观察表明(图2),过量铜处理的雪柑侧根线粒体模糊,但沙田柚侧根线粒体膜结构清晰可见。相比对照,过量铜处理的两个柑橘品种幼苗侧根细胞壁增厚。不论是对照还是过量铜处理,沙田柚侧根细胞壁厚度均显著高于雪柑(图3)。

图1 不同铜浓度处理下两个柑橘品种幼苗侧根细胞壁提取率Fig.1 The extraction rate of lateral roots cell wall of two citrus species under different Cu concentrations
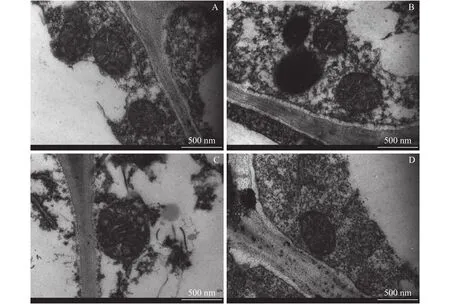
图2 不同铜浓度处理下两个柑橘品种幼苗侧根细胞壁微观形态Fig.2 The micromorphology of lateral roots cell wall of two citrus species under different Cu concentrations
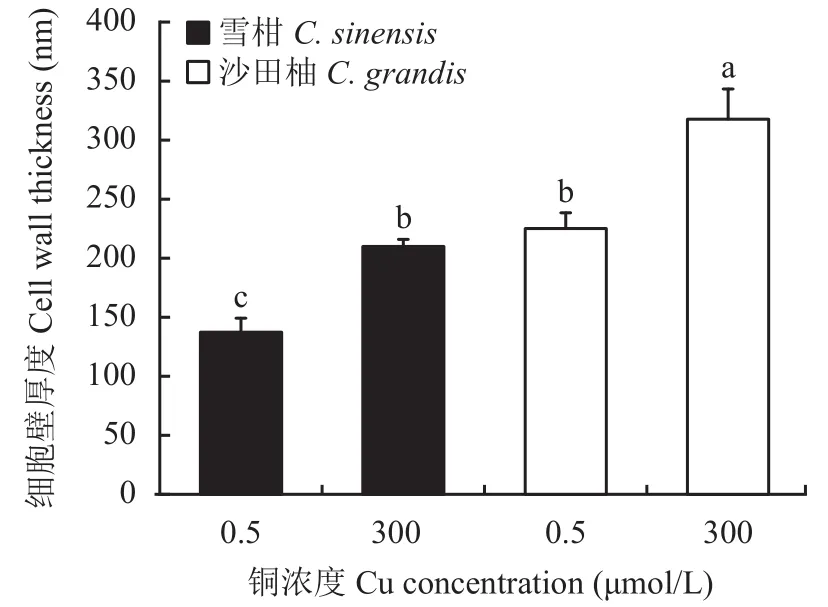
图3 不同铜浓度处理下两个柑橘品种幼苗侧根细胞壁厚度Fig.3 The lateral root cell wall thickness of two citrus species under different Cu concentrations
2.5 铜处理的两个柑橘品种幼苗侧根细胞壁FTIR分析
对照和过量铜处理后的雪柑和沙田柚幼苗侧根细胞壁的 FTIR 谱图如图4 所示。3338 cm–1(No.1)附近的吸收峰为—OH和—NH伸缩振动峰的叠加,主要来自于根细胞壁中脂肪酸、蛋白质和碳水化合物(果胶、半纤维素、纤维素、多糖),2921 cm–1(No.2)附近的吸收峰为甲基C―H键的对称伸缩振动,主要来自于细胞壁中亲水脂分子,1736 cm–1(No.3)附近的吸收峰归属为酯化果胶中酯基C=O的伸缩振动,1652 cm–1(No.4)处的吸收峰为酰胺Ⅰ带C—N伸缩,是细胞壁蛋白质上的特征峰,1508 cm–1(No.5)附近的吸收峰为蛋白质上的酰胺Ⅱ带N―N弯曲和伸缩特征吸收峰,1423 cm–1(No.6)附近的吸收峰归属为果胶质糖醛酸中羧酸根离子COO–的特征吸收峰,1374 cm–1(No.7)附近的吸收峰为纤维素中甲基的伸缩振动吸收峰,1337 cm–1(No.8)附近的吸收峰对应于纤维素中醇羟基的C―O弱吸收峰,1247 cm–1(No.9)处对应硫酸盐C―O―S或羧基C―O或磷酸盐 C―O―P 的特征吸收峰,1150 cm–1(No.10)左右为果胶的多糖环结构的C―C或C―O吸收峰,1035 cm–1(No.11)附近的吸收峰主要为纤维素的多糖链C―C的特征吸收峰。

图4 不同铜浓度处理下的两个柑橘品种幼苗侧根细胞壁红外光谱Fig.4 The FTIR spectra of Cu-treated lateral roots cell wall of two citrus species
参考张虹等[37]的方法,以 2921 cm–1处―CH3中C―H的特征吸收峰的吸光度A2921为标准值,计算其它特征峰的吸光值与A2921的比值,通过该比值的大小变化半定量分析过量铜处理的雪柑和沙田柚侧根细胞壁官能团特征峰变化(表4)。雪柑和沙田柚幼苗经铜处理后,根细胞壁上的官能团在No.1特征峰(—OH/—NH)、No.6特征峰(羧酸盐C=O不对称伸缩振动)和No.10特征峰(果胶多糖环结构C—C或C—O)发生了较明显的偏移,且雪柑主要向高频偏移,沙田柚主要向低频偏移。另外,雪柑在No.4特征峰(酰胺Ⅰ带C—N)处向高频偏移了3 cm–1,沙田柚在No.3特征峰(酯基C=O)、No.5特征峰(酰胺Ⅱ带N—N)及No.9特征峰(硫酸盐C—O—S或羧基C—O或磷酸盐C—O—P)处向低频偏移,表明—OH、果胶、蛋白质、羧酸盐、硫酸盐、磷酸盐及纤维素在一定程度上参与了铜吸附。与对照相比,过量铜处理后沙田柚A1734/A2921高于雪柑A1737/A2921,表明过量铜处理的沙田柚根细胞壁中果胶质甲基酯化程度上升明显;过量铜处理后,沙田柚 A1653/A2921和 A1509/A2921上升值分别高于雪柑A1655/A2921和A1509/A2921,表明过量铜处理的沙田柚根细胞壁蛋白质含量升高显著;与对照相比,过量铜处理后,沙田柚A1373/A2921和A1244/A2921比值升高,而过量铜处理后雪柑的A1373/A2921和A1247/A2921比值较对照组降低,可知过量铜使沙田柚根细胞壁中的纤维素、硫酸盐、羧酸盐及磷酸盐含量增加。过量铜处理后,相比沙田柚,雪柑根细胞壁A3293/A2921和A1426/A2921的比值有明显降低,意味着其根细胞壁中―OH和果胶在铜处理下显著减少;过量铜处理后,相比沙田柚,雪柑的A1337/A2921、A1156/A2921和A1035/A2921的比值下降较大,可能与过量铜处理后的雪柑根细胞壁中主要组分纤维素含量减少有关。
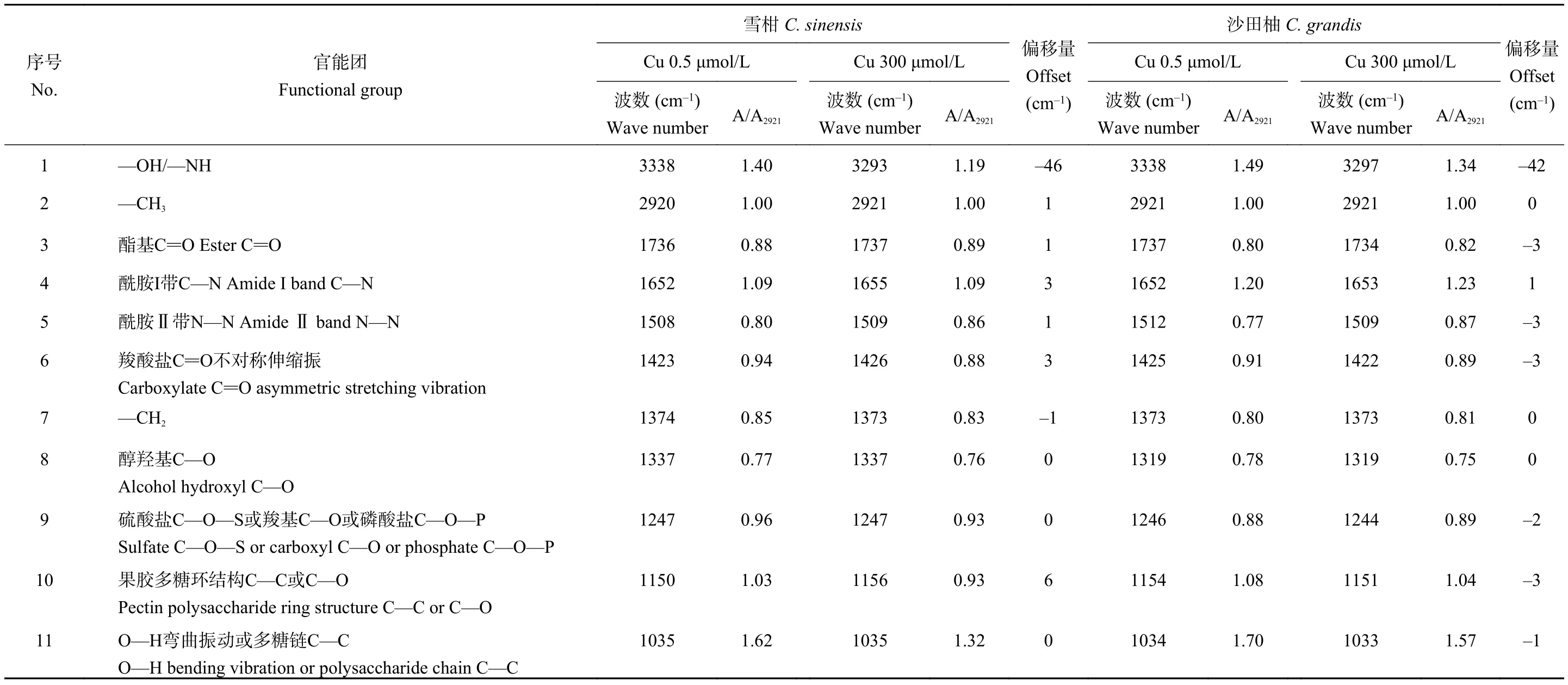
表4 不同铜浓度处理下两个柑橘品种幼苗侧根细胞壁红外光谱特征峰的半定量分析Table 4 The semi-quantitative analysis of FTIR spectra of Cu-treated lateral roots cell wall of two citrus species
3 讨论
根是植物最早感知根际重金属浓度,启动胁迫响应最关键的部位[38]。铜在根中的固定,减少了其向地上部的装载,起到保护地上部同化器官的作用。如,Hippler等[39]研究表明,24 μmol/L 铜处理 90 天后的水培甜橙幼苗根中累积的铜占植株吸收铜总量的80.0%。本研究结果进一步表明,根际铜过量时,柑橘苗侧根是铜主要的累积部位。这与史萍等[40]在铜处理的牡丹根中的研究结果一致。相比铜处理的主根,雪柑和沙田柚幼苗侧根的铜浓度分别提高了10.6和13.2倍。铜处理下,柑橘苗组织水平上的铜分布特征表现为侧根>主根>茎>叶。相比对照,过量铜处理显著提高了柑橘幼苗不同组织的铜浓度。300 μmol/L砂培铜处理的沙田柚侧根铜浓度显著高于雪柑可能与沙田柚侧根对铜的固定能力较强有关。
亚细胞组分中重金属的分布特征与植物耐性机制有关[41]。本研究显示,在亚细胞水平上,铜主要分布在对照组柑橘幼苗侧根的细胞器组分。过量铜处理下,细胞器中的铜相对含量显著降低,但细胞壁和可溶性部分铜含量占比显著提高。这与柑橘铜响应机制有关。当柑橘幼苗根际铜浓度较低时,铜主要存在于细胞器,参与细胞代谢反应。当铜浓度过高时,过量铜主要分布于细胞可溶性部分与细胞壁,而细胞器铜相对浓度显著降低可以减少过量铜对细胞器的结构损伤和功能破坏。通常认为,根细胞壁是植物响应重金属胁迫的“第一道屏障”[42]。例如,刘婷婷[43]揭示了水培条件下,铜浓度为100 μmol/L时,耐铜植物海州香薷根细胞壁铜累积量在亚细胞水平占比高达68.6%。类似的报道显示,过量锌处理的栾树、臭椿和银杏根细胞壁中锌的累积量最高可占亚细胞锌含量的82.6%[44]。胡萝卜侧根细胞壁是过量铅富集的主要部位[45]。玉米根细胞壁也是铜[46]、镉[47]和钒[48]等重金属的主要分布区域。不同的是,过量镉在油菜根中的分布特征表现为细胞液>细胞器>细胞壁[49]。本研究结果显示,尽管300 μmol/L砂培铜处理都显著提高了两个柑橘品种幼苗根细胞壁和可溶性部分的铜含量,但相比细胞壁,可溶性部分结合的铜含量占比更高。这与经铅、镉和锌处理的马缨丹[41]以及经砷处理的狭叶香蒲[50]上的研究结果一致。亚细胞可溶性部分富含小分子有机酸以及螯合态蛋白等,可以有效结合并固定铜,通过液泡对过量铜的区隔化作用降低细胞铜损伤[51]。鉴于细胞可溶性成分种类多,成分杂,有待通过质谱分析与蛋白分离等技术进一步研究。
根细胞壁多糖组分由葡萄糖、阿拉伯糖以及半乳糖醛酸等单糖聚合而成[52]。半乳糖醛酸富含半乳糖残基,可作为细胞壁的阳离子吸附位点[53]。报道显示,细胞壁各多糖组分的铜结合能力存在差异。例如,任超[54]报道了铜主要结合于蓖麻根细胞壁的半纤维素上。李芹等[55]的研究也表明,半纤维素Ⅰ是水稻根细胞壁蓄积镉的主要场所。相反,周冉[56]和张涵[57]等分别报道了镉和铝主要结合在植物根细胞壁的果胶上。本研究通过差速离心法分离柑橘幼苗侧根细胞壁多糖组分并分析各多糖组分的铜累积量。铜在柑橘侧根细胞壁多糖组分的累积量表明,果胶是铜在柑橘侧根细胞壁的主要结合位点,其结合的铜浓度占细胞壁铜浓度的百分比高于98%。
研究表明,不同重金属胁迫对细胞壁厚度的影响并不一致。例如,过量铅[58]和镉[59]使小麦根细胞壁变薄。相反,镉毒害引起苔藓[60]和钩沙菜[61]细胞壁增厚。本研究发现,过量铜处理的柑橘幼苗根细胞壁厚度显著增加。细胞壁多糖组分富含大量可与金属离子结合的官能团,细胞壁的增厚可结合更多的金属离子,从而降低重金属对植物的毒害。本研究还发现,过量铜处理的两个柑橘品种幼苗侧根细胞壁提取率(图1)较对照组都显著提高。可见,细胞壁受过量铜影响做出了应激反应,通过增加细胞壁的量以结合更多的铜,对细胞内部进行保护。细胞壁含量提高且厚度增加可能是Kulikova等[62]提出的过量铜引起植物根尖膨大的主要原因。两个柑橘品种比较发现,过量铜处理的沙田柚侧根细胞壁铜含量(表2)、细胞壁提取率(图1)与厚度(图3)都显著高于雪柑。因此,柑橘幼苗侧根细胞壁特性可能是影响两个柑橘品种幼苗铜分布特征与耐铜差异的关键因素。
细胞壁果胶、纤维素和蛋白质等组分对铜具有较高的吸附和固定能力,主要源于羟基、羧基和氨基官能团[63]。FTIR谱图是植物细胞壁各主要化学成分的综合反映,可以根据相应红外光谱吸收峰找到对应的官能团。特征峰吸光度的比值越高,暗示该官能团的含量越高,当官能团参与铜的吸附时,根细胞壁上官能团的特征吸收峰会发生不同程度的位移和强度的变化,官能团没有参与铜的吸附时,其特征吸收峰的位移不变或变化较小[42]。在本研究中,两个柑橘品种的特征吸收峰向低频或高频的位移程度表明,果胶、纤维素和蛋白质处的吸光值与A2921比值大小变化显著,这与刘思怡等[64]和曹升等[65]在茶树上的研究报道类似。
4 结论
相比对照,过量铜处理显著提高了两个柑橘品种幼苗各部位的铜含量。组织水平上,过量铜主要富集在柑橘苗侧根;亚细胞水平上,过量铜主要结合于侧根可溶性部分和细胞壁果胶组分。品种比较表明,过量铜处理的沙田柚侧根铜含量、侧根细胞壁铜含量以及侧根细胞壁果胶铜含量均显著高于雪柑。提高细胞壁含量、增厚细胞壁以及促进细胞壁官能团对铜的吸附可能是沙田柚耐铜性强于雪柑的主要原因。
