MC1R基因影响禽类羽毛颜色变异的研究进展
2022-07-15蒋婧孙晓燕任航行
蒋婧 孙晓燕 任航行*
1.重庆市畜牧科学院,重庆 402460;2.重庆市山羊工程技术研究中心,重庆荣昌 402460
自然界中,禽类羽毛颜色五彩缤纷、变幻多端。羽色既是辨别种类与性别的重要表型性状,也能产生重要的经济效益。禽类羽毛颜色表现为结构颜色和化学颜色的结合,结构颜色是由光反射产生的,化学颜色是由包含在羽毛毛囊中的黑色素造成的[1]。黑色素是一种典型的化学生色色素,存在2 种类型,即真黑色素(黑色至棕色)和褐黑色素(红色至黄色)[2]。这2 种色素在黑素细胞中产生,黑素细胞是主要存在于表皮、眼睛和毛囊中的特化细胞。黑色素在专门的细胞器——黑素体中产生,然后通过树突状突起运输到生长中的毛发或角质细胞中[3]。绚丽多彩的鸟类羽毛的颜色正是这些黑色素类型的结合体。黑色素的合成受复杂的生物学信号通路网络调控,与多种黑素生成基因相关,其中,黑皮质素1 受体基因(melanocortin 1 receptor,MC1R)是重要的调控基因之一,在控制黑素细胞合成的黑色素类型中起着至关重要的作用。
MC1R是黑素细胞上的一种七跨膜螺旋蛋白偶联受体,属于G 蛋白偶联受体(G protein-coupled receptors,GPCRs)超家族,人的MC1R基因结构复杂,含有多个剪接变体且高度多态[4-6],不仅在色素沉积方面发挥重要功能,还参与DNA 修复、促炎细胞因子的调控、棕色脂肪组织代谢等其他关键的生物学通路,与黑斑、痣、皮肤癌、白癜风、帕金森等多种疾病相关[7-11]。但禽类MC1R基因结构相对简单,主要在发育中的羽毛和毛发的黑素细胞中表达,通过与其天然配体α-黑素细胞刺激素(α-melanocytestimulating hormone,α-MSH)或其拮抗剂刺鼠信号蛋白(agouti signaling protein,ASIP)结合,调控真黑素和褐黑素的合成[12]。MC1R的高活性导致黑色/棕色真黑色素的合成,而低活性导致红色褐黑色素的合成或真黑色素合成的缺失,从而影响发育中的羽毛中产生的黑色素的类型和数量[13]。本文现对MC1R基因对家禽(鸡、鸭)和野生鸟类(仓鸮、鹌鹑)等禽类羽毛颜色变异的影响进行综述。
1 MC1R 基因对家禽羽毛颜色变异的影响
1.1 MC1R 基因对鸡羽毛颜色变异的影响
鸡的MC1R基因位于11 号染色体,其编码的E基因座调控毛囊中黑色素的合成,被证明是控制鸡羽色表型的最重要基因座[14]。基于此,鸡羽毛类型主要对应E(Extended)、ER(Birchen)、ewh(Wheaten)、e+(Wild-type)、eb(Brown)、es(Speckled)、ebc(Buttercup)和ey(Recessive Wheaten)等8 个等位基因,它们的显隐性关系为:E>ER>ewh>e+>eb>es>ebc>ey[15]。早在20世纪初,Ling 等[16]就确认了各种等位基因对应的表型和氨基酸位置(表1)。e+基因型为野生型,其公鸡表型为黑色胸、红色背,母鸡背部为棕色斑点、胸部为浅棕色。eb、ewh和ey基因型的公鸡表型外观和e+基因型一样,只是羽毛的底色即靠近皮肤的羽毛颜色与野生型的灰色底色有所不同,表现为白色或者奶油色;ewh和ey基因型的母鸡羽色均为浅棕色,与e+基因型母鸡的胸部羽色相同;eb基因型的母鸡背部和翅膀的羽色与e+基因型母鸡一样,均为棕色斑点,但其胸部颜色也表现棕色斑点,与e+基因型胸部羽色不同。E 等位基因的公鸡和母鸡全身都为黑色羽毛。ER等位基因的公鸡与野生型相比,其飞羽由半红半黑变为全黑,母鸡有金色或银色的颈羽,且所有的公鸡和母鸡的表皮均有黑色素沉着。
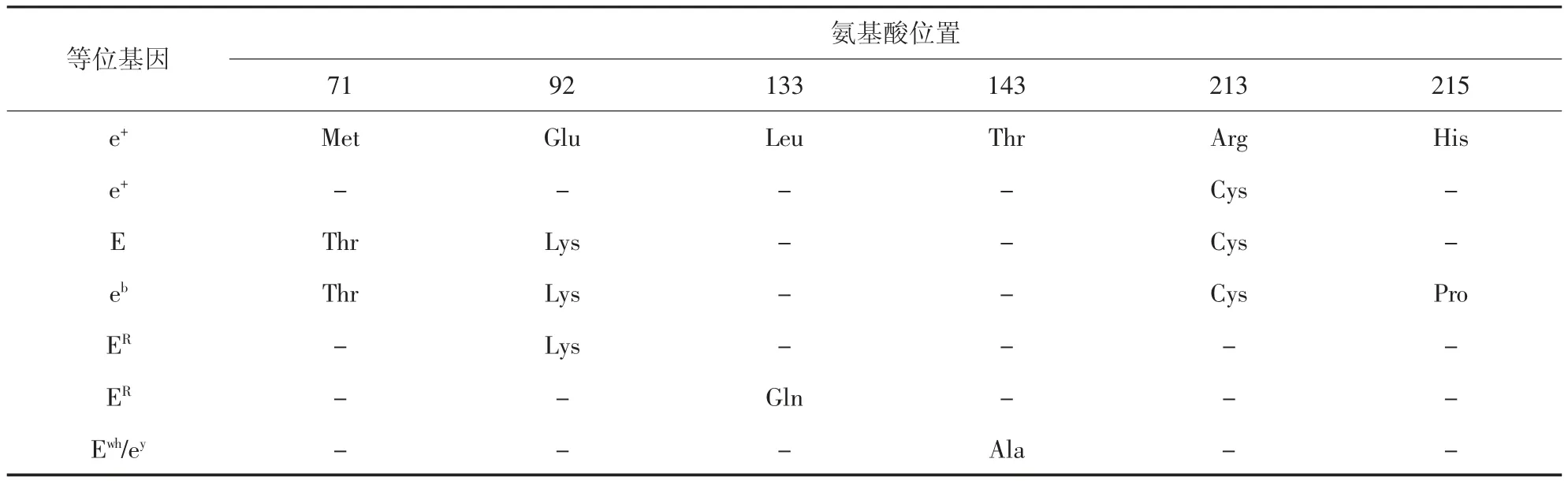
表1 E 基因座上的等位基因对应的氨基酸位置[16]
国内外鸡品种中发现MC1R基因存在大量SNPs,除了表1中所出现的突变位点外,后来的研究又发现了一些新的突变位点。冉金山[17]在广元灰鸡(灰羽)、旧院黑鸡(黑羽)、罗曼蛋鸡(白羽)、芦花鸡(横斑羽)和HS1(麻羽)群体中发现MC1R基因存在3 个错义突变,分别是T398A(p.L133Q)、T637C(p.C213R)和G920C(p.R307P),它们与鸡羽色的形成有较强的相关性。其中T398A 在黑羽和麻羽群体中没有发现,T637C 与T398A 均是前人已经验证过的位点,G920C 在黑羽群体中没有出现。在乌骨鸡群体中[18],共发现23 个SNPs,其中15 个位于启动子区(C-2536T、T-2260C、C-2257T、C-2175T、T-2022C、A-1952G、C-1934T、G-1925A、C-1924G、C-1881T、C-1876G、A-1750G、A-1737G、C-693T、G-405A),8 个位于外显子区(Exon1-C69T、Exon1-T212C、Exon1-G274A、Exon1-T398C、Exon1-G636A、Exon1-T637C、Exon1-A644C、Exon1-C834T)。经过基因多态位点与黑色素关联分析,在MC1R基因上发现4 个与黑色素相关物质关联的位点,即C-2175T、Exon1 -G23A、Exon1 -T398C 和 Exon1 -T212C,可作为乌骨鸡乌度选育的遗传标记或辅助参照。Zhang 等[19]在4 种不同羽色的鸡群体中发现2个同义SNP(C69T 和G636A)和8 个非同义SNP(T212C、A274G、A356G、A427G、A551G、T637C、A646C 和T881C),其中3 个突变(p.E92K、p.N119G和p.I194T) 位于跨膜区、4 个替换(p.M71T、p.T143A、p.C213R 和p.H215P)位于细胞膜外、1 个替换(p.N184S)位于膜外。MC1R突变位点在不同品种、不同羽毛颜色群体中的基因频率和基因型频率的研究结果表明,C69T、T212C 和A274G 与瓦灰色羽毛显著相关。在含有5 种羽色表型的鸡群体中检测到MC1R基因存在3 个SNP,即T398A(p.L133Q)、T637C(p.C213R)和G920C(p.R307P),基于这3 个SNP 获得了6 个单倍型(H1-H6),其中H1(TTG)单倍型在各种羽毛颜色中出现频率较高,推测H1 单倍型是优势单倍型[20]。H5(ATC)和H6(ACC)的单倍型主要集中在白鸡和灰鸡,在黑鸡中没有发现,因此猜测2 种单倍型可能部分抑制黑色素的产生[20]。在对日本品种和非日本品种鸡MC1R基因的研究中[21],发现了3 个同义核苷酸替换和11个非同义核苷酸替换,产生15 个单倍型(H0-H14),其中H9、H13 和H14 是新发现的3 种单倍型,H9 由已知的C69T、T212C、G274A 和G636A 组成;H13 除了已知的C69T、G178A、G274A、G636A和T637C 外,还有新发现的非同义突变C919G;H14包括已知的C69T、G274A 和G409A 和新发现的C919G。此外,A644C 取代(H5 单倍型)纯合子的鸟类的羽毛颜色首次被定义为小麦样,A427G 和G274A 替代分别与褐色和黑色羽毛颜色有关。MC1R基因G274A 的AA 基因型在黑色羽毛的河北鸡优势表达,而G376A 位点的GG 基因型却除黑河北鸡以外的所有群体中都很常见,表明这2 个SNP 真黑色素色素沉着中起重要作用。有趣的是,多项研究发现G409A(Ala137Thr)可能抑制G274A(Glu92Lys)的效果,这种效果通常会导致黑色羽毛的形成[21-22]。
1.2 MC1R 基因对鸭羽毛颜色变异的影响
鸭的羽毛颜色也受MC1R基因的表达和多态性调控,Wang 等[23]在研究腹部白色和背部深色的浅褐色绿头鸭背腹颜色变异的机制时,发现MC1R等8 个色素相关基因在不同部位的表达存在差异,提示背腹羽色差的分子调控机制可能与白鸭和黑鸭相似,是由于黑色素合成途径中多个基因表达不同所致。董飚等[24]在番鸭MC1R基因编码区发现G274A(p.E92K)、G279A、C485A(p.T162N)突变,其中G279A 突变的GG 基因型只在白羽番鸭群体中仅有,推测该突变对番鸭的羽色影响较大。夏波[25]在家鸭品种中检测到MC1R基因2 个突变(Ala137Thr、Arg217Cys)的不同基因型在不同黑色素表型群体中的分布存在明显差异,表明MC1R基因对家鸭羽色具有重要影响。一项对13 个品种鸭MC1R基因多态性的研究发现G52A(p.K18E)、G376A(p.I126V)、G409A(p.R137T)和C649T(p.R217C)位点,其中G52A 和G376A 与黑色羽毛个体显著相关[26]。后来的研究证实了这一结果,并进一步发现G52A 和G376A 对增强MC1R活性有促进作用,从而促进鸭羽毛中真黑素的沉积[27]。虽然A399G突变位点对羽色未造成影响,但它可以从本质上有效鉴定和区分家鸭、白番鸭和半番鸭三类鸭群体,可成为番鸭品种保护的新方法[28]。
2 MC1R 基因对野生鸟类羽毛颜色变异的影响
2.1 MC1R 基因对号鸟属羽毛颜色变异的影响
在野生动物中,MC1R基因表达水平与色素沉积相关性状有关,灰林鸮的羽色呈浅红色到深红色渐变,雏鸟黑色素的着色程度与MC1R基因的表达水平显著正相关,当父母的红色较深时,其后代MC1R基因的表达水平较高[29]。有趣的是,MC1R 基因不同位点的错义突变会导致突然的颜色变化,从而导致种群内部或种群之间出现不同的颜色变化。Burri 等[30]发现欧洲仓鸮的MC1R基因p.I126V 的杂合子和II 纯合子个体羽色比VV 纯合子个体羽色更红,并进化出了一种新的红色表型。该位点在矛隼、绿头鸭群体中也得到了验证[31],这种缬氨酸-异亮氨酸替代与较深羽毛颜色相关,同时既改变了黑素生成相关基因的表达,还改变了颜色与MC1R上游的黑素皮质蛋白基因表达之间的关系[32]。表明MC1R突变可以通过多效性黑素皮质蛋白基因调节色素沉着。北极贼鸥和仓鸮中同时存在的p.R8H 位点也与羽毛颜色相关[33]。此外,MC1R基因多态性还与鸟类迁徙进化有关,来自英伦三岛的仓鸮羽色呈白色,比欧洲大陆任何种群都要白,在这些群体中,红色的MC1R等位基因几乎是不存在的,但在相同纬度的欧洲大陆上,其频率接近50%,那里的仓鸮羽色更深一些[33]。因此证明,英伦三岛的仓鸮并不是由欧洲大陆迁徙而来,且携带一个红色等位基因的单个副本就足以确保较深的表型[34]。
2.2 MC1R 基因对鹌鹑羽毛颜色变异的影响
日本鹌鹑通过与不同羽毛颜色的鹌鹑杂交,在羽毛颜色上表现出多态性,包括黑色、黄色、白色、野生型(栗色)和各种中间色。黑色鹌鹑的MC1R表达高于野生型鹌鹑和白色鹌鹑,在野生型日本鹌鹑胚胎中瞬时表达过MC1R基因可导致皮肤黑素生成增加,这在表型上类似于黑色鹌鹑[35]。褐羽鹌鹑和野生型鹌鹑群体中发现的p.E92K 突变与羽毛表型密切相关,该突变位于MC1R的跨膜2 区,可能通过激活MC1R基因来增加cAMP 活性和TYR 活性从而导致真黑色素增加[36]。在中国彩羽鹌鹑群体中,除了鉴定出已被证实可导致显性黑色羽毛的p.E92K突变外,还发现另一个错义突变p.P292L,该突变可能与鸟类的受体失活相关,不直接影响真黑素/褐黑素的平衡,而影响羽毛图案和/或颜色分布[37]。此外,在日本鹌鹑群体中发现的MC1R基因p.I57V 突变也与黑羽鹌鹑的黑色表型显著相关[38]。
3 小结和展望
自然界中,无论是家禽还是野生鸟类,其羽毛颜色都是绚丽多彩的,这一特性除了是禽类本身重要的表型性状外,也能产生不菲的经济效益,因此研究禽类羽毛颜色变幻多端的分子机制很有意义。禽类羽毛丰富的颜色是由结构颜色和化学颜色共同构成,其中化学颜色与皮肤和毛囊中黑色素细胞的增殖、分化、迁移和黑色素的合成以及色素沉积强烈相关。调节黑色素细胞发育和黑色素合成的生物信号网络相当复杂,截至目前已被确认参与其中的基因超过了150 个[39],MC1R基因正是其中最重要的基因之一。在对鸡、鸭、鸮和鹌鹑群体MC1R基因的研究发现,它们既有相同的突变位点影响羽色的变异,也存在特有的突变位点,说明MC1R基因在物种间存在差异,但总的来说,无论是MC1R基因表达还是其多态性都与禽类羽毛颜色变异显著相关。禽类群体产生的大量自发突变为解析MC1R基因调节黑色素生成的分子机制提供了便利,研究结果既拓展了黑色素合成机制的调控网络,又为禽类的遗传选育提供参考。MC1R基因对黑色素细胞和黑色素的调控并不是独立的,而是与黑色素调控网络的其他基因共同调控的,因此,在未来,需进一步开展MC1R基因与其他基因协助或交叉调控的研究,配合生物信息学分析、全基因组关联分析等新型研究手段,真正弄清楚禽类羽毛颜色变异背后的分子机制。
