线粒体蛋白组解析烟草雄性不育分子机制
2022-07-04柯渔洲王平松段丽丽莫泽君喻奇伟刘仁祥
柯渔洲 王平松 段丽丽 莫泽君 何 轶 喻奇伟 熊 晶 刘仁祥,*
(1 贵州大学烟草学院/贵州省烟草品质研究重点实验室,贵州 贵阳 550025; 2 贵州省烟草公司毕节市公司,贵州 毕节 551700)
胞质雄性不育是由核基因组与细胞质基因组共同控制的、表现出雄蕊不能产生有活力的花粉而雌蕊能正常授粉结实的母系遗传现象,普遍存在于多种植物中[1-2]。目前,我国烟草杂交种和不育系种植越发广泛,培育烟草不育系和杂交种品种能促进杂种优势的利用,不仅可以省去人工去雄环节,而且能明显提高杂交种纯度和制种效率[3]。但花粉的形成和发育受多重因素的影响,雄性不育形成的分子机制极其复杂。大多数线粒体蛋白由核基因编码,并在细胞质的核糖体上合成前体蛋白,经线粒体膜上的TOM复合体(translocase of the outer mitochondrial membrane)通道进入线粒体内部[4],且线粒体作为主要供能单位提供细胞代谢活动所需的90%的能量。因此,通过线粒体蛋白组学探究烟草胞质雄性不育的形成机制具有重要意义。
在烟草雄性不育性的成因研究中,前人主要从细胞形态学方面比较小孢子发育的异同[5-7],明确其败育的方式和时期。目前已有研究利用线粒体基因组重测序技术挖掘胞质雄性不育基因[8-10],先后发现了orf768a[11]、atp6[12]、orf25[13]、atp9[14]、ms1[15]等多个不育候选基因,利用基因剪辑技术进行基因功能验证,表明胞质雄性不育与线粒体存在密切关系[16-18],但对烟草胞质雄性不育形成的分子机制鲜见报道。因此,本研究选用烟草雄性不育系云烟87(MSYY87)及其保持系云烟87(YY87)为试验材料,利用石蜡切片结合线粒体蛋白质组学及生物信息学分析方法,探究烟草胞质雄性不育形成的分子机制,明确线粒体蛋白的表达模式,以期为揭示烟草胞质雄性不育的分子机理提供理论基础。
1 材料与方法
1.1 试验材料
以烟草雄性不育系云烟87及其同核异质保持系为试验材料,材料由贵州省烟草品质研究重点实验室提供。于2021年在安顺市西秀区贵州大学烟草科研基地种植,采用3次重复随机随机区组设计。在移栽后25、30、35、40、45 d的花芽分化时期选取长势均匀的烟株6株,其中3株取顶端5 cm处的花芽在甲醛乙酸乙醇固定液(formaldehyde-acetic acid-ethanol,FAA)中固定,用于石蜡切片观察,切片厚度8~10 μm,番红固绿对染,加拿大树脂封片,拍照观察。另外3株立刻用液氮保存,后置于-80℃低温冰箱,后续用于线粒体蛋白分析试验。
1.2 花芽线粒体蛋白的提取和质检
选择分化过程中出现形态差异时期的烟草花芽作为提取线粒体蛋白的材料。采用GMS30034.4v.4线粒体蛋白提取试剂盒(GENMED SCIENTIFICS INC,美国)提取花芽总线粒体蛋白。使用Bradford法绘制标准曲线,计算待测样品的蛋白浓度。采用12%的聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate poly acrylamide gel electrophoresis, SDS-PAGE)对蛋白质进行分离,凝胶使用eStain LG蛋白染色仪(金斯瑞生物科技有限公司,重庆)进行考马斯亮蓝染色后,应用全自动数码凝胶图像分析系统成像。
1.3 TMT标记和液质检测
6个蛋白样品经胰蛋白酶Trypsin-TPCK酶切后,分别加入100 mmol·L-1的四乙基溴化铵(tetraethyl ammonium bromide, TEAB)缓冲液和含有不同同位素的串联质谱标签(tandem mass tages,TMT)标记的乙腈,涡流混匀,室温反应1 h,加入5%羟胺终止反应15 min,然后冻干于-80℃保存。使用1100 HPLC高压液相色谱系统(Agilent,美国)进行组分分离和液相色谱检测,依次每隔1 min收集洗脱液到1~15号离心管中,然后收样60 min,收集好后真空冷冻干燥抽干。对样品进行液相二级质谱(liquid chromatorgraphy-mass spectrometry/mass spectrometry, LC-MS/MS)分析,质谱上机条件:一级图谱(MS)分辨率设为 70 000, 自动增益控制值设为1e6,最大注射时间为50 ms;质谱扫描设定为全扫描荷质比(m/z)范围300~1 600, 并对其中10个最高峰进行二级图谱(MS-MS)扫描;所有图谱采集使用数据依赖型的正离子模式下的高能碰撞裂解完成,碰撞能量设为32;MS/MS-MS的分辨率设为17 500,自动增益控制设为2e5,离子最大累积时间为80 ms;动态排除时间设为30 s,最终生成质谱检测原始数据。
1.4 线粒体蛋白鉴定
利用Proteome Discoverer 2.4软件对检索结果进一步过滤,可信度在99%以上的谱肽为有效图谱数(peptide spectrum matches,PSM),至少包含一个肽段的蛋白为可信蛋白,保留可信的有效图谱肽和蛋白,通过错误比例(false discovery rate,FDR)值验证,去除FDR大于0.01的肽段和蛋白。使用烟草基因组结合uniprot-proteome_UP000084051.fasta作为搜索数据库对鉴定到的蛋白进行功能注释和代谢通路注释。
1.5 蛋白定量分析和差异表达蛋白功能分析
根据原始下机的图谱峰面积可以得到各个样品中每个PSM的相对定量值,再根据鉴定出的unique肽段中所包含的PSM定量信息校正得到unique肽段的相对定量值,最后根据每个蛋白质包含的所有unique肽段的定量信息校正得到每个蛋白的相对定量值。将每个蛋白在比较样品中表达量与对照组表达量的比值作为差异倍数(fold change,FC)。将每个蛋白在样品中的相对定量值进行T检验,P值为显著性指标。当FC≥1.2,且P≤0.05时,蛋白表达量上调;当FC≤0.83,且P≤0.05时,蛋白表达量下调。针对筛选出来的差异蛋白进行基因注释(gene ontology,GO)和代谢通路富集(kyoto encylopedia of genes and genomes,KEGG)分析,与所有鉴定到的蛋白背景相比,应用超几何检验方法计算P值,以P≤0.05为阈值,满足此条件的条目即为在差异蛋白质中显著富集的条目。
2 结果与分析
2.1 不育系与保持系的花器官形态特征
由花器官形态特征观察可以看出,MSYY87雄蕊花丝缺失或畸变、无花药,柱头变粗并出现部分弯曲,表现为完全雄性不育(图1-A)。而YY87花丝发育正常,花药饱满,花药略高于柱头(图1-B),能正常进行自花授粉。
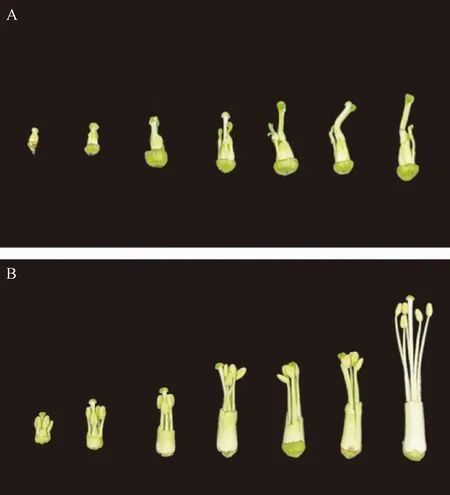
注:A:不育系;B:保持系。Note: A: Sterile line. B: Fertile line.图1 MSYY87和YY87烟草花器官形态变化Fig.1 Morphological changes of floral organs of tobacco MSYY87 and YY87
2.2 不育系与保持系的小孢子发育形态观察
随着叶片数的增加,烟株由营养生长转向生殖生长,这段生理生化变化被称为花芽分化过程。观察烟草MSYY87及YY87的花芽石蜡切片发现,MSYY87在花芽分化起始阶段(图2-A)能正常分化出花序原基;但在雌雄蕊原基分化时期(图2-B),只有3个雄蕊原基且相互粘连,形态异常,不能正常产生孢原细胞;花粉母细胞时期(图2-C),绒毡层细胞提前发生程序性死亡,使孢原细胞不能正常分裂为花粉母细胞,最终花丝畸变或雄蕊缺失(图2-D),无花药。

注:a~d: 不育系;e~h: 保持系。a、e:花序原基形成期;b、f:雌雄蕊原基分化时期花芽横切面×200;c、g:花粉母细胞时期×200;d、h:花粉成熟期(d×50,h×200)。TP:花序原基;PP:雌蕊原基;SP:雄蕊原基;E:表皮层;En:内皮层;ML:中间层;T:绒毡层;P:雌蕊;S:雄蕊;PG:花粉粒。Note: a~d: Sterile line. e-h: Maintainer line. a,e: Inflorescence primordium formation stage. b,f: Cross section of flower bud at the primordia differentiation stage×200. c,g: Pollen mother cell stage×200. d,h: Pollen maturity stage(d×50, h×200). TP: Inflorescence primordium. PP: Pistil primordium. SP: Stamen primordium. E: Epidermis. En: Endodermis. ML: Middle layer. T:Tapetum. P: Pistil. S: Stamens. PG: Pollen grains.图2 烟草MSYY87与YY87小孢子发育形态差异Fig.2 Morphological differences of microspore development between MSYY87 and YY87 in tobacco
YY87在花序原基形成期(图2-E)正常分化出花序原基,此时烟株出叶速度快,随着发育进行,逐步分化出1个雌蕊原基和5个雄蕊原基,随着孢原细胞继续平周分裂产生造胞细胞;造胞细胞(图2-G)能分裂形成花粉母细胞,花粉母细胞继续分裂产生小孢子,在药室中有成熟花粉(图2-H)。选择在雌雄蕊原基分化时期的花芽来做线粒体蛋白组分析。
2.3 蛋白的质检与分析鉴定
线粒体蛋白的SDS-PAGE结果显示,各样品的条带清晰,说明提取线粒体蛋白质量较好,未受到污染(图3)。利用可信蛋白的表达量进行各样本间的主成分分析(图4),从不同维度可发现MSYY87与YY87样品间存在明显差异,可进行后续试验。再通过数据库搜索,鉴定到二级图谱(MS-MS)总数 285 815 个,有效图谱数(PSM)5 828个,肽段(peptides)数 3 580 个,蛋白数(protein groups)1 251个。

图3 烟草花芽线粒体蛋白的SDS-PAGE图谱Fig.3 SDS-PAGE map of mitochondrial protein from tobacco flower bud

图4 烟草花芽线粒体蛋白质样品的主成分分析Fig.4 Principal component analysis of mitochondrialprotein samples from tobacco flower buds
2.4 差异表达蛋白
根据限制条件筛选差异表达蛋白,在注释到的 1 251 个蛋白中,对每个蛋白质差异倍数以2为底取对数,将P值以10为底取对数的绝对值,绘制火山图(图5)以分析可信蛋白在不同样品间的上调、下调情况。2组样品无显著差异的蛋白为735个,差异表达蛋白总数为113个,其中CMSYY87较YY87显著上调的蛋白为48个,显著下调的蛋白为65个。说明MSYY87与YY87线粒体蛋白的表达存在差异,上述差异蛋白可能是导致不育系产生的关键原因,进一步推测其可能在不育系的线粒体中发生代谢紊乱。

图5 MSYY87与YY87差异表达蛋白火山图Fig.5 Volcano diagram of differentially expressed proteinsof MSYY87 and YY87
2.5 差异蛋白的GO功能富集分析
差异蛋白GO显著性富集分析主要包括生物学过程、细胞组分以及分子功能。经GO功能富集分析,两类烟草样品中的113个蛋白去除仅包含1个差异蛋白的条目,剩余30个可靠的GO条目。其中,达到显著富集(P<0.05)水平的GO条目为30个,属于生物学过程10个、细胞组分10个、分子功能10个(图6)。生物过程类别分别为糖酵解过程(glycolytic process)、脂质运输过程(lipid transport)、未折叠蛋白细胞响应(cellular response to unfolded protein)、内质网对未折叠蛋白响应(endoplasmic reticulum unfolded protein response)、重折叠蛋白过程(protein refolding)、内质网泛素依赖性降解途径(ubiquitin-dependent ERAD pathway)、蛋白重折叠依赖性伴侣结合蛋白(chaperone cofactor-dependent protein refolding)、脂类代谢过程(lipid catabolic process)及内质网应急响应(response to endoplasmic reticulum stress)。
注释到细胞组分,分别包括细胞溶质(cytosol)、细胞膜(membrance)、细胞核(nucleus)、内质网膜泡(endoplasmic reticulum lumen)、内质网膜互作复合体(endoplasmic reticulum lumen chaperone complex)、液泡(vacuole)、磷酸丙酮酸水合酶复合体(phosphopyruvate hydratase complex)及完整地膜组分(integral component of membrane)。
注释到的分子功能,分别为营养库活性(nutrient reservoir activity)、蛋白质二硫键异构酶活性(protein disulfide isomerase)、酰基甘油脂肪酶活性(acylglycerol lipase activity)、错误折叠结合蛋白(misfolded protein binding)、折叠蛋白伴侣(protein folding chaperone),磷脂酶活性(phospholipase activity)、及热休克蛋白结合物(heat shock protein binding)、磷酸丙酮酸水合酶活性(phosphopyruvate hydratase activity)、二磷酸果糖醛缩酶活性(fructose-bisphosphate aldolase)及葡萄糖醛酸脂脱羧酶活性(UDP-glucuronate decarboxylase activity)。

图6 GO富集柱状图Fig.6 Histogram of GO enrichment
2.6 差异蛋白的KEGG代谢通路分析
通过对差异蛋白进行KEGG代工谢通路分析(图7),主要分为有机物代谢途径、遗传物质代谢及次生代谢物合成和抗氧化代谢三类。有机物代谢主要为细胞呼吸的系列反应,主要包括糖酵解和糖原异生途径(glycolysis/gluconeogenesis)、果糖和甘露糖代谢(fructose and mannose metabolism)、氨基糖和核苷酸糖代谢(amino sugar and nucleotide sugar metabolism)、磷酸戊糖途径(pentose phosphate pathway)、光合固碳有机体(carbon fixation in photosynthetic organisms)、丙酮酸代谢(pyruvate metabolism)、三羧酸循环(tricarboxylic acid, TCA)、甘油酯代谢(glycerolipid metabolism);遗传物质代谢途径主要包括RNA降解(RNA degradation)、内质网加工蛋白过程(protein processing in endoplasmic reticulum)、蛋白外运(protein export)、蛋白酶体(proteasome)、色氨酸代谢(tryptophan metabolism)、组氨酸代谢(hisidine metabolism);次生代谢产物合成和抗氧化代谢主要包括过氧化物酶体(peroxisome)、抗坏血酸和醛达酸代谢(ascorbate and aldarate metabolism)、谷胱甘肽代谢(glutathione metabolism)、类黄酮生物合成(flavonoid biosynthsis)、苯丙素生物合成(phenylpropanoid biosynthsis)。

图7 KEGG富集气泡图Fig.7 KEGG enrichment bubble diagram
3 讨论
通过烟草花芽的石蜡切片结果可知,MSYY87在发芽分化早期(雌雄蕊原基分化期)开始出现小孢子发育异常现象,这与前人的烟草雄性败育发生在雄蕊原基分化后至孢原细胞形成前的观点基本一致[5-6]。因此选用雌雄蕊分化时期的花芽进行线粒体蛋白组学分析。TMT标记是一种高分辨率高通量的蛋白定量检测技术[19]。本试验通过线粒体蛋白质组学TMT标记结合高效液相色谱质谱联用技术和生物信息学分析,共鉴定出1 251个蛋白,在MSYY87和YY87的花芽中表达差异显著的线粒体蛋白113个,其中48个蛋白在MSYY87的花芽中高表达,65个蛋白在YY87的花芽高表达。两类样品中表达差异的蛋白富集到7个类别细胞组分,分别为细胞溶质、细胞膜、细胞核、内质网膜泡、内质网膜复合体及细胞液泡。这与线粒体蛋白要经核基因在细胞质核糖体上翻译、加工成前体蛋白,再运输到作用部位线粒体的观点[20]相一致,说明本研究采用的线粒体蛋白组研究方法可行,鉴定出的差异蛋白具有较高的可信度。
GO功能富集分析主要注释到生物过程中糖酵解和脂的运输代谢,脂肪转运和代谢过程通常在线粒体基质中进行[21-22],对呼吸代谢有着重要的意义;由于线粒体相关呼吸代谢酶差异表达,致使线粒体代谢紊乱[23-24],注释到的分子功能中营养库活性、磷酸丙酮酸水合酶及二磷酸果糖醛缩酶等呼吸酶,推测线粒体代谢紊乱与烟草雄性不育有关。蛋白质空间结构的修饰影响其功能和作用。已有研究报道二硫键异构酶亚基[25-26]与热休克蛋白[27]参与协同催化氧化蛋白折叠修饰过程,在拟南芥[27]中线粒体热休克同源蛋白(mtHSC70-1)缺失突变体的花形态、花药发育存在明显缺陷,导致育性显著减低,且经蛋白组学分析发现,在线粒体呼吸中电子传递和蛋白折叠的生物学过程发生改变。注释到的生物过程有未折叠蛋白细胞反应过程、未折叠蛋白内质网反应、蛋白质重折叠过程、内质网泛素依赖性降解途径、重折叠蛋白的伴侣蛋白复合体及内质网应急反应过程,及注释到分子功能中的蛋白二硫键异构酶活性、错误折叠结合蛋白及热休克蛋白,说明不育系中的线粒体蛋白的修饰受到影响。当线粒体受到细胞内外的胁迫刺激,会通过HDA-1调控线粒体未折叠蛋白反应的途径来维持线粒体内的稳态和功能,当线粒体受损信号传递至细胞核,可促进核编码的应激基因上调表达,从而保护和修复受损线粒体[28]。由此即推测,当烟草不育系线粒体基因与核基因之间互作不协调的信号不能传递给细胞核,则不能促进核编码的应激基因上调表达,修复线粒体功能紊乱中断,从而影响小孢子的形成和发育。

注:图中字符表示酶或者亚基。红色表示上调蛋白,绿色表示下调蛋白。Note: The characters in the figure indicate enzymes or subunits. Red represents up-regulated proteins and green represents down-regulated proteins.图8 MSYY87花芽线粒体代谢紊乱图Fig.8 Mitochondrial metabolic disorders in sterile lines
植物呼吸代谢是在线粒体中将有机物逐步氧化并释放能量供其利用的生物学过程[29]。通过KEGG通路富集分析发现,磷酸戊糖途径的6-磷酸葡萄糖异构酶G6PI(glucose-6-phosphate isomerase)、1,6-二磷酸果糖醛缩酶F16BA(fructose-bisphosphate aldolase)、转醛酶T(transaldolase)、6-磷酸葡萄糖脱氢酶G6PD(6-phosphogluconate dehydrogenase)都下调表达,推测磷酸戊糖途径代谢受阻(图8)。首先糖酵解途径中磷酸己糖异构酶(GPI)、果糖醛缩酶FA(fructose aldolase)、烯醇化酶E(enolase)、乙醇脱氢酶AD(alcohol dehydrogenase)都表现下调表达,说明丙酮酸生成受到抑制,其次在三羧酸循环途径中苹果酸脱氢酶(malate dehydrogenase,MD)和异柠檬酸脱氢酶(isocitrate dehydrogenase,ID)下调表达,然后在氧化磷酸化过程中焦磷酸酶(inorganic pyrophosphatase,IP)表达下调,ATP合酶的F1中α亚基(F-type H+-transporting ATPase subunit alpha)和δ亚基(ATPase subunit delta)蛋白表达上调,说明ATP合酶结构异常,最终结果说明由于蛋白酶下调和亚基异常导致不育系线粒体呼吸代谢过程ATP合成受阻。这与Wang等[30]研究相似,即细胞缺失C9orf72线粒体蛋白后,在合成ATP的氧化磷酸化途径活性下调,最终诱导细胞死亡,其原因是C90r72蛋白通过招募抗增值蛋白(prohibitin,PHB)来阻断氧化磷酸化中复合体I组装因子TIMMDC1的降解,稳定复合体I的有效组装和维持正常产能反应。
植物线粒体中RNA编辑调控线粒体PPR蛋白的形成[31]。RNA的稳态由转录速率和降解速率之间的平衡决定,核糖体RNA的稳定可能受核糖体蛋白装配过程影响[32]。本研究发现核糖体RNA的大亚基L4e(large subunit ribosomal protein L4e)、L7(large subunit ribosomal protein L7)及小亚基中SAe(small subunit ribosomal protein SAe)的表达异常,调控RNA降解的烯醇酶EL(enolase)下调表达(图8),说明核糖体RNA失调会影响线粒体蛋白装配。多数线粒体蛋白由核基因编码[33],经核糖体翻译成的前体蛋白由线粒体外膜TOM复合体通道进入线粒体内,当线粒体蛋白导入受阻,通过改变蛋白酶体活性和伴侣蛋白水平调控线粒体蛋白表达[34-36]。研究发现,Ubx2与线粒体膜上TOM复合体结合并通过蛋白酶体去除未成功导入的前体蛋白,维持TOM通道的流畅[4]。本研究发现不育系中内质网蛋白加工酶(endoplasmic reticulum chaperone BiP)、蛋白二硫键异构酶(protein disulfide-isomerase,PDIs)和磷脂酶(phospholipase D,PLD)均下调表达,使线粒体蛋白的修饰和导入受阻,且蛋白酶复合体的PA700亚基Rpt3(proteasome regulatory subunit T3)和20S小亚基α5(proteasome subunit alpha 5)表达下调(图8),导致蛋白酶体功能失调,不能降解未导入线粒体的前体蛋白,使线粒体蛋白的导入受阻而功能紊乱,不能维持花粉形成时细胞快速分裂分化所需的能量而小孢子形成中断。
4 结论
本研究发现,烟草胞质雄性不育发生在发芽分化的雌雄蕊原基分化时期;不育系在线粒体蛋白修饰和导入过程中,内质网蛋白加工酶(BiP)、蛋白二硫键异构酶(PDIs)和磷脂酶(PLD)等下调表达,说明线粒体蛋白导入受阻,引起线粒体功能紊乱。存在线粒体呼吸代谢中6-磷酸葡萄糖脱氢酶(G6PD)、苹果酸脱氢酶(MD)和焦磷酸酶(IP)等表达下调,说明呼吸代谢受阻。由此推测,烟草胞质雄性不育由于线粒体蛋白的合成、修饰及导入受阻,致使线粒体功能紊乱,即在呼吸代谢途经中蛋白酶下调表达,抑制了ATP的合成,从而不能为小孢子形成过程中的快速分裂分化提供充足的能量和营养物质,抑制了小孢子的形成和发育,表现为雄性不育。
