单原子材料类酶催化及生物医学应用研究进展
2022-06-29许维庆吴志超顾文玲朱成周
沙 蒙,许维庆,吴志超,顾文玲,朱成周
(华中师范大学化学学院,农药与化学生物学教育部重点实验室,智能生物传感技术与健康国际联合研究中心,武汉 430079)
新型高效生物催化剂的开发在化学研究中占有重要地位[1].在自然界中,酶是一种高效的生物催化剂,由于其具有高催化活性与强的特异性,已广泛应用于工业生产、生物与医疗等领域[2,3].近年来,随着生物技术和纳米材料科学的重大进展,纳米材料可以模拟天然酶的性质、结构和生物学功能,在特定的生物催化中表现出与酶相似的作用机理[4,5].2007年,Yan等[6]首次报道铁磁纳米粒子具有模拟过氧化物酶的活性并引起了广泛的关注.2013年,Wei和Wang[7]定义了“具有酶学性质的纳米材料”为纳米酶.纳米酶结合了化学催化剂和生物催化剂的特性,兼具纳米材料固有的物理化学特性(磁、光、热及电学等)和类酶活性.纳米酶的发现揭示了纳米材料内在的生物效应及新特性,丰富了模拟酶的内涵,为纳米技术与生物学、医学等多学科的连接架起桥梁[8,9].得益于经济、稳定、性质可调、可大规模生产和回收利用等优势,纳米酶克服了天然酶在实际应用中的不足[10,11].近年来,人们发现并设计了多种纳米酶,包括金属、金属氧化物或硫化物和碳基纳米酶等[12~16].此外,大量研究表明,通过调节尺寸[17]、形貌[18]、晶面[19]、价态[20]及配位结构[21]等新策略,能够特异性地调控纳米酶的催化性能[22],这为寻求更高效的纳米酶并探索其生物催化机制提供了可能.
目前,纳米酶领域仍然面临着多方面的挑战.首先,传统纳米酶的活性位点的密度和质量活性较低,在催化过程中仅有表面1~2原子层参与反应,导致其类酶催化效率远低于天然酶[23,24].其次,纳米材料复杂的结构特征使得催化剂活性位点的识别较为困难.如晶面、尺寸、元素组成以及杂原子掺杂等因素都会导致复杂的催化机制和不同的催化路径,难以精准识别类酶活性位点及揭示构效关系[25,26].此外,纳米材料虽然可以模拟出天然酶的性质和功能,但由于本身缺乏精细结构,难以精确模拟天然酶催化中心复杂的配位结构和催化微环境,致使其催化的选择性和活性不理想[3,27].
随着催化科学迈入微观尺度研究,当金属组分以原子级分散的形式存在时,可以最大限度发挥金属原子的利用效率,单原子材料应运而生,成为当前材料科学与催化科学研究的热点[28,29].单原子材料的出现可以很好地应对当前纳米酶急需解决的困境.首先,单原子材料是材料调控的极限,降低了成本并实现了金属原子利用效率的最大化[30];其次,金属原子与载体之间的强相互作用,既能稳定单原子,也能有效调控其电子结构,对提高催化活性和选择性有重要意义[31,32].单原子催化剂明确的活性结构位点,为纳米酶类酶催化机制探索提供了很好的模型体系,有利于揭示催化剂的构效关系[33,34].在众多报道的单原子催化剂中,碳载单原子催化剂,尤其是M-N-C催化剂,其金属原子与周围氮原子之间形成的稳定配位键(M-Nx)[35],在能源催化与生物催化领域已展现出潜在的应用价值[36~39].特别是其金属活性中心与金属酶活性中心M-Nx在几何结构和化学组成上有很大的相似性,如辣根过氧化物酶[40,41]及细胞色素P450[42]等催化中心具有血红素Fe-N4的结构,由此可以实现类酶催化活性的有效模拟[43],这些发现进一步促进了新型类酶材料的理性设计和调控优化,推动了类酶催化本质的探索.
单原子催化剂具有明确的均相催化和多相催化的活性中心,但由于在合成和后续处理过程中有原子聚集的趋势,因此在制备合成方面仍然面临挑战.为了防止原子聚集并加强金属-载体相互作用,研究人员已经开发了一系列合成单原子催化剂的策略,包括自下而上和自上而下两类[44,45].单原子催化剂的构建通常采用自下而上策略,包括质量分离软着陆法[46,47]、原子层沉积法[48,49]、湿化学法[50]和球磨法[51]等,主要利用载体的缺陷和空位位点锚定金属前驱体,并通过化学还原得到单原子催化剂.自上而下策略是以较大尺度的单质材料或者纳米颗粒为前驱体,在特定条件下解离,实现向单原子位点的转变,主要包括高温热解、高温原子迁移[52,53]等.目前对单原子材料的表征技术主要包括球差校正高分辨透射电子显微镜、高分辨的透射电子显微镜和X射线精细结构谱等.X射线精细结构谱能够得到中心原子价态、配位数、配位原子的种类和中心原子与配位原子之间的键长等配位信息.先进的表征手段有利于识别活性位点结构并深入理解单原子类酶材料结构与性能的关系[54~57].本文总结了近几年来单原子材料在模拟酶催化方面的研究进展.首先,主要介绍了单原子类酶材料在类过氧化物酶(POD)、类氧化酶(OXD)、类过氧化氢酶(CAT)和类超氧化物歧化酶(SOD)等方面的研究,同时重点讨论了调控单原子材料类酶活性的策略和类酶催化机理.随后,系统地总结了单原子类酶材料在癌症治疗、抗氧化治疗、抗菌和生物传感等方面的具体应用(Scheme 1).最后,概述了单原子类酶材料发展的机遇和挑战.
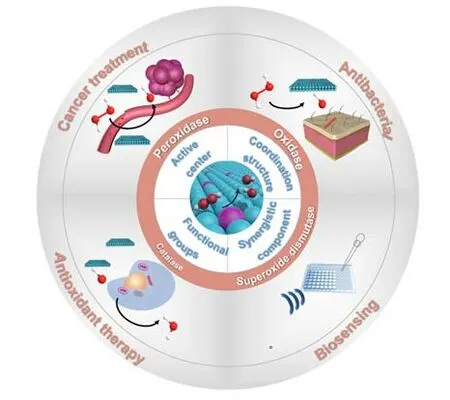
Scheme 1 Single⁃atom materials for enzyme⁃like catalysis and biomedical applications
1 单原子材料的类酶活性
1.1 类过氧化物酶
POD 在自然界中广泛分布,由一大类酶组成,如较为常见的辣根过氧化物酶(HRP)和细胞色素c过氧化物酶等.过氧化物酶可以在H2O2或其它有机过氧化物存在下催化底物的氧化.在催化过程中,过氧化物被还原,底物通常作为电子给体[6].近年来,诸多单原子催化剂被报道具有优异的类过氧化物活性,并被广泛地应用于生物传感和医疗等多个领域[58,59].为了进一步提高单原子催化剂的类过氧化物酶活性,研究者们通过调控金属活性中心类型、提高单原子负载量、调控中心原子配位环境及引入官能团或协同催化组分等策略实现了活性调控.
1.1.1 Fe基单原子类过氧化物酶 天然酶催化活性中心具有卟啉铁辅基结构,即Fe与卟啉环平面上的4个N配位.因此研究者们致力于开发Fe基单原子材料来模拟自然酶活性中心[60],通过调控Fe单原子材料独特的配位方式和电子结构,从而实现对天然酶催化性能的高效模拟.Jiao等[50]采用葡萄糖和双氰胺分别作为碳源和氮源,利用葡萄糖中的含氧基团有效螯合Fe2+以避免团聚,一步退火合成了Fe-N-C单原子类酶材料.所合成的Fe-N-C单原子材料具有Fe-N4活性结构并表现出优异的类过氧化物酶活性.
金属活性中心的类型和数量是单原子材料类酶催化的核心和关键.为了进一步提高单原子材料的类过氧化物酶活性,调控金属活性中心类型并提高单原子负载量是非常有效的策略.Jiao等[61]采用盐模板法,合成了高负载量和高暴露量的Fe-N-C,Co-N-C 和Zn-N-C 二维超薄单原子类酶材料.其中,Fe-N-C 材料中Fe 的负载量高达13.5%(质量分数),远超当时已报道的Fe 基单原子催化剂.扩展X 射线吸收精细结构拟合得到Fe,Co,Zn 与N 配位数接近于4.实验结果表明,Fe-N-C 的比活性(25.33 U/mg)显著优于Co-N-C(6.33 U/mg)和Zn-N-C(2.46 U/mg)[图1(A)].理论计算表明,活性位点FeN4吸附H2O2分子后发生均裂生成吸附态OH*;随后一个OH*游离出来形成·OH,为反应的决速步骤.相对于CoN4和ZnN4活性位点,FeN4与吸附态OH*的结合能力最弱且反应能垒更小,与其优异的催化性能相一致[图1(B)].
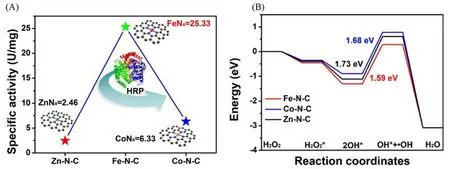
Fig.1 Experimental and theoretical studies on the POD⁃like activity of Fe⁃N⁃C[61]
调控单原子材料催化活性的另一个重要策略是调控中心原子的配位结构.中心原子的配位结构在很大程度上影响中心原子的电子结构[29].不同的配位结构会导致中心金属原子与邻近原子之间的电子相互作用不同,这对提高催化性能具有重要意义.影响单原子位点配位结构的主要因素有配位原子、配位数和配位空间结构等[32].将S,B,P等富电子且易掺杂的原子引入到单原子的配位环境中,一方面引起结构的扭曲和电荷密度的改变,通过杂原子与邻近原子的电负性差异调控活性中心的电子结构[62];另一方面可以在基底上引入几何缺陷,进一步提高催化活性和稳定性.Jiao 等[63]将杂原子B引入到Fe-N-C单原子材料中(FeBNC),实现了对中心原子电子结构的有效调控[图2(A)],可以有效增强类过氧化物酶活性[图2(B)].理论计算表明,B诱导的电荷转移效应可以调节活性中心Fe原子的正电荷,从而降低羟基自由基(·OH)形成的能垒[图2(C)],最终导致FeBNC的类过氧化物酶的活性增强.在此研究的基础上,Jiao等[64]将原子尺寸更大的S原子引入到Fe-N-C单原子材料中(FeSNC),S原子会突出NC平面,导致NC骨架的结构畸变.S原子的引入不仅实现了对活性中心的几何结构的调控,同时也实现了对中心原子电子结构的优化.实验结果表明,以Fe-N3S1为活性位点的FeSNC单原子类酶材料相较于以Fe-N4为活性位点的Fe-N-C,类过氧化物酶活性显著增强.理论计算结果表明,S诱导的几何和电子效应不仅增加了吸附态H2O2的O—O键的键合距离,还加速了Fe与O之间的电子转移,从而降低了反应中间体的形成能垒,实现了类过氧化物酶活性的提高.P和S在原子半径和电负性方面具有相似性,并且P是许多天然酶的重要组成元素,在底物与天然酶活性中心的电子传递中发挥重要作用.Ji 等[27]以沸石咪唑盐骨架8(ZIF-8)为载体,聚(环三磷腈-4,4′-二氨基二苯醚)(PZM)为P 的前驱体,锚定金属活性中心Fe,聚合的PZM包覆在ZIF-8表面,得到Fe/ZIF-8@PZM核壳复合材料.进一步退火得到以FeN3P为活性中心的单原子类酶材料.该单原子材料表现出优异的类过氧化物酶活性,具有与HRP相当的催化效率和选择性.理论计算表明,在吸附氧形成的过程中(2OH→O+H2O),由于异原子P 的引入,在FeN3P 位点上生成的O 吸附在P 上,H2O 吸附在Fe 上,P 的吸附作用导致O 和H2O之间形成氢键的键长最短,反应能垒最小.在随后的决速步骤中,吸附在P上的O迁移到Fe位点,反应能垒低,易于发生.此外,配位数的不同影响着单原子活性位点的几何和电子结构,很大程度上影响了类酶催化性能.Xu 等[65]受自然酶配位环境启发,通过将具有Fe-N4四配位的血红素锚定在超薄N掺杂石墨烯上,合成了活性位点为Fe-N5结构的单原子类酶材料(NG-Heme).作为对比,通过直接将血红素固定到石墨烯上,构筑了Fe-N4四配位的单原子类酶材料(G-Heme).与四配位结构的G-Heme相比,五配位结构的NG-Heme 的类过氧化物酶活性提高了42.1倍.理论计算表明,得益于NG-Heme 中轴向配体与Fe 的相互作用,使得NG-Heme 的d带中心(−0.86 eV)比G-Heme(−1.06 eV)更高,从而促进中间体的生成.
除了直接调控与金属中心配位的原子类型和原子数,Xu等[66]通过对金属有机框架(MIL-101)中有机配体进行官能化,实现对空间邻近的金属活性位点电子结构的直接调整.结果表明,—NO2的引入能显著提高MIL-101 类酶催化活性和对底物的亲和力.而—NH2的引入则恰恰相反[图2(D)和(E)].为了进一步研究不同官能团导致活性差异的背后原因,他们通过理论计算首先研究了—NO2的引入对吸附态的几何效应.由于—NO2的强吸电子作用,使得的O—H 键取向偏向—NO2上的氧,H…O—N—O氢键距离缩短,这有利于O—O键的断裂,降低OH*形成能.而NH2-MIL-101吸附的O—H键没有明显变化.其次,他们研究了—NO2的引入对MIL-101的电子效应.—NO2的引入改变了活性位点Fe的3d轨道分裂,降低了悬空键()上的电子密度,从而增强了中间体OH*的吸附,降低了决速步骤的反应能垒[图2(F)].
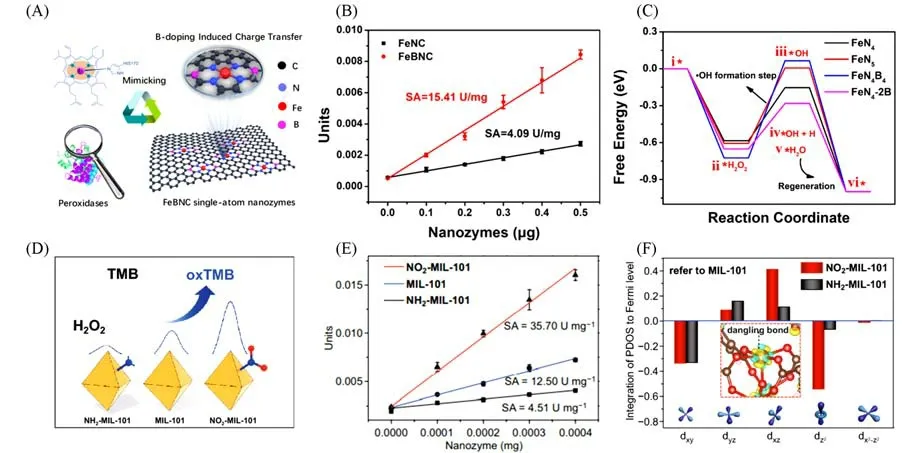
Fig.2 Regulation of coordination structure and functional groups
此外,通过引入一些协同催化的组分也能够进一步调控单原子类酶催化性能.Chen等[67]通过将Pt簇限域在层状多孔的Fe-N-C载体上,构建了单原子与簇偶联的高性能类酶材料(FeSA-PtC).实验结果表明,FeSA-PtC类过氧化物酶活性比Fe单原子提高了4.5倍,比Pt簇提高了7倍.进一步探究表明,Fe单原子与Pt簇之间存在协同作用,单原子Fe上的电子转移到了Pt簇上,这可能是其活性提升的原因.
1.1.2 其它过渡金属基单原子类过氧化物酶 除了Fe基单原子类酶材料,其它过渡金属基单原子类酶材料也有相关报道.Chen等[68]采用一种非高温退火的策略,通过配位作用直接将金属Cu离子锚定在MOFs 的有机配体2,2′-联吡啶-5,5′-二羧酸上,构建了Cu-MOF 单原子类酶材料[图3(A)].其表现出类过氧化物酶活性,在H2O2存在的条件下,可以催化荧光红染料(Amplex-Red)氧化生成有荧光信号的试卤灵[图3(B)].同时在H2O2存在下催化多巴胺生成氨基色素[图3(C)].
除此之外,Zn,Mo,Co基单原子类酶材料也有相关报道.Xu等[69]仿照天然酶卟啉环的配位特点,将金属有机框架ZIF-8作为前驱体,利用介孔硅保护策略,合成了具有Zn-N4活性中心的Zn基单原子类酶材料.该Zn基单原子材料具有较好的类过氧化物活性,催化H2O2产生·OH,可以实现99.85%的抑菌率.此外,Wang等[70]报道了一系列以Mo为活性中心的单原子类酶材料,通过调控Mo位点的配位数实现了对类过氧化物酶特异性的精准调控.他们首先通过密度泛函理论(DFT)模拟,在一系列Mo-Nx-C单原子模型中确定了Mo-N3-C 是过氧化物酶特异性的最佳侯选模型.然后,通过客体模板策略,在800,900 和1000°C 的热解条件下,分别合成了不同配位环境的MoSA-N4-C,MoSA-N3-C 和MoSA-N2-C.扩展X射线吸收精细结构光谱表明,伴随着热解温度的升高,Mo-N/O峰强度逐渐降低,其配位数逐渐变低.实验结果证实,Mo-N3-C过氧化物酶特异性最好,且与Mo结合的N原子数在活性与特异性中起关键作用.理论计算表明,3种类酶材料活性位点吸附的H2O2都能自发分解,且分解产物高度依赖于Mo的配位环境.MoSA-N4-C和MoSA-N2-C中的Mo嵌入平面内,吸附的H2O2会通过异裂的方式自发生成O*和H2O,而MoSA-N3-C 中的Mo 突出平面外,吸附的H2O2更有利于均裂成2 个OH*.研究还发现,MoSA-N3-C的最高占据分子轨道更容易水平吸附H2O2的最低未占据分子轨道,而MoSA-N2-C和MoSA-N4-C则更倾向于吸附垂直构型的H2O2.
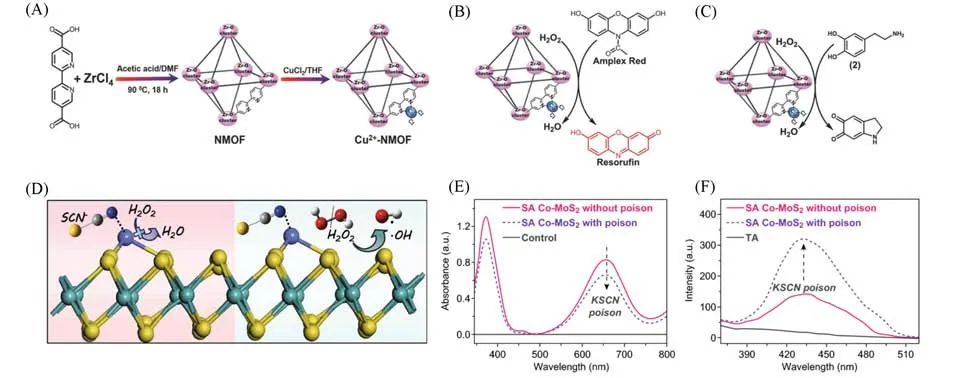
Fig.3 Cu⁃centered and Co⁃centered single⁃atom materials for mimicking peroxidase
目前报道的单原子类酶材料绝大多数是以无机碳材料作为基底,对金属化合物等载体及载体与活性位点的相互关系等研究较少.金属化合物(如硫族化合物)也是广泛用于稳定单原子的载体.金属化合物表面的不饱和位点可以锚定单原子,有效防止金属聚集.载体中的金属位点与单原子位点的相互作用也是影响催化活性的重要因素,而天然酶也往往因为相邻位点的协同作用而表现出高反应活性.Wang 等[71]采用超薄的MoS2纳米片作为载体吸附Co 纳米盘,通过电化学刻蚀得到Co 单原子类酶材料(SA Co-MoS2),该单原子材料具有较高的类过氧化物酶活性.他们分别从实验和理论计算两个方面揭示了Co单原子中心和MoS2载体遵循不同的催化机理[图3(D)].Co活性中心遵循电子转移机制,MoS2则遵循类芬顿反应的机制[图3(E)和(F)],Co单原子和MoS2协同提升类过氧化物酶活性.
1.2 类氧化物酶
OXD 属于氧化还原酶的一类,可催化O2直接氧化底物.在氧化酶催化过程中,氧气作为电子受体,可以接受4 个电子被还原成H2O,接受2 个电子生成H2O2,接受1 个电子生成.氧化酶种类很多,如细胞色素P450[21]、烟酰胺腺嘌呤二核苷酸(NADH)氧化酶[72]和多酚氧化酶等.目前,已有报道发现单原子材料具有类氧化酶活性[73,74].
1.2.1 模拟细胞色素P450 Huang等[25]报道了一种用金属有机框架包覆铁酞菁的限域策略合成的具有轴向五配位的单原子类酶材料(FeN5SA/CNF).值得注意的是,其原子级分散的活性位点与细胞色素P450活性中心的空间结构十分相似[图4(A)].实验结果表明,FeN5SA/CNF的催化速率远远优于FeN4SA/CNF[图4(B)]和商业Pt/C.理论计算结果表明,与四配位的FeN4SA/CNF 相比,轴向配位的N的引入使FeN5SA/CNF具有较强的电子推动作用,有利于电子填充到O2的反键π*轨道,进一步促进O—O键的裂解,从而使得吸附态氧更容易从底物3,3′,5,5′-四甲基联苯胺(TMB)中得到H+,提高吸附态氧对底物的氧化能力[图4(C)].
目前,关于单原子催化剂模拟氧化酶,研究最多的底物仍然是TMB.通过激活O2生成活性氧,然后氧化底物TMB,完成整个反应过程.Xu等[75]合成的单原子催化剂Fe-N-C不仅模拟了细胞色素P450的Fe-Nx血红素活性中心,而且模拟了细胞色素P450在人体内的功能性.细胞色素P450参与了人体内很多内源性和外源性的药物代谢,他们选取了二氢吡啶降压药(1,4-DHP)作为Fe-N-C 类P450 氧化酶的底物,实现了Fe-N-C对1,4-DHP药物的代谢模拟[图4(D)].不仅如此,此工作还将生物催化与电催化氧还原(ORR)进行比较,揭示了不同的反应机制[图4(E)].电催化ORR与氧化酶催化反应都是活化O2变成H2O的过程,有所不同的是,电催化ORR反应中电子由电极传递到催化剂表面,而氧化酶催化反应中电子由底物提供.他们通过将含铁的离子液体前驱体在850 ℃下热解得到了石墨结构较多的Fe-N-C-850,在400 ℃下热解得到了石墨结构较少的Fe-N-C-400.石墨结构在ORR 应用中起至关重要的作用,因为直接通过石墨框架,电子更容易从电极转移.实验也证明了Fe-N-C-850具有更高的ORR活性,而低温热解的Fe-N-C-400具有更好的类氧化酶活性.由此可见,石墨结构并不是驱动类酶氧化反应的关键因素[图4(F)].
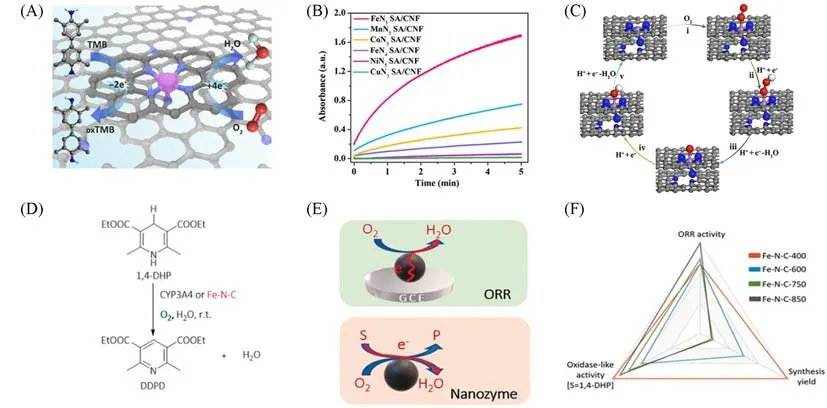
Fig.4 Single⁃atom materials for mimicking the active center of cytocrome P450
1.2.2 模拟多酚氧化酶 含铜的蛋白质广泛参与生命的各种氧化还原过程.如漆酶(氧化苯酚和芳香胺类)、血蓝蛋白(O2转运)、酪氨酸酶(氧化单酚类)和儿茶酚氧化酶(氧化儿茶酚类),在其活性部位都有偶联的多核铜(II)中心[76].通过构建原子级分散的含Cu催化剂,可以实现模拟Cu基氧化酶活性的目的.Liang 等[77]通过用配体鸟嘌呤腺苷单磷酸与二价铜离子配位,合成了具有原子尺度分散的无定形金属有机框架材料(Cu/GMP),是首次报道能够模拟漆酶的材料.漆酶是一种含4个铜离子的氧化酶,可以氧化生物环境系统中比较重要的底物(如多酚、多胺等),也可以直接活化氧气生成水,无H2O2产生.通过实验证明,该类酶材料在4-氨基安替吡啉的存在下可以催化底物2,4-二氯苯酚生成红色产物,并在相同质量浓度下表现出比漆酶更高的催化活性.除了2,4-二氯苯酚,该类酶材料对萘酚、对苯二酚、邻苯二酚和肾上腺素等多种底物均具有催化作用.在酚类氧化酶中除了漆酶,儿茶酚氧化酶也在生物系统中发挥着重要作用,可以催化邻苯二酚类底物氧化成相应的邻醌类;邻醌类可聚合产生黑色素,形成一种不可溶解的屏障以保护受损植物免受病原体的伤害.天然的儿茶酚氧化酶活性中心由耦合的双核铜与6个组氨酸配位组成的,在O2作用下催化氧化邻苯二酚,O2被还原成过H2O2,进一步通过O—O键的裂解形成水.Li等[78]合成具有三核铜中心的MOF-818,不仅模拟了儿茶酚氧化酶的活性结构,且具有高效的儿茶酚氧化酶活性和高特异性,不表现出过氧化物酶活性.与天然酶有所不同的是,MOF-818在O2存在下催化邻苯二酚氧化成邻苯二醌,生成的是H2O2,而不是H2O.
1.3 类过氧化氢酶和类超氧化物歧化酶
过氧化氢酶作为另一类含金属辅基的生物催化剂,能够分解H2O2产生H2O 和O2.Wang 等[79]通过将单原子Ru 锚定在金属有机框架Mn3[Co(CN)6]2中,进一步封装光敏剂Ce6,构建了Ru 单原子催化剂.组装的Ru单原子催化剂具有较好的类过氧化氢酶活性,能够催化细胞内的H2O2原位产生O2,缓解肿瘤微环境的缺氧状态.为了进一步丰富单原子材料模拟酶的类型,研究者们对单原子材料的类超氧化物歧化酶活性进行模拟.超氧化物歧化酶作为生物体内普遍存在的一种抗氧化金属酶,可以催化歧化生成O2和H2O2.Ma等[80]通过框架结构原位封装前驱体的策略合成了活性位点为Fe-N4的单原子类酶材料,表现出优异的超氧化物歧化酶活性,催化转化为H2O2和O2,可以有效清除细胞氧化应激产生的活性氧.
1.4 多酶活性
单原子材料是一种无机催化剂,缺乏如酶分子一样精巧的三维空间结构,并且单原子的载体缺少对特定底物的识别位点.然而,酶的催化选择性高度依赖于活性位点附近的空间结构,尤其是活性中心周围存在的重要氨基酸对底物的识别、活化起重要作用.因此,单原子类酶材料很难达到天然酶的选择性.虽然对类酶材料进行合理设计以提升选择性一直是纳米酶研究领域不懈追求的目标,但在实际应用中具有多酶活性的纳米材料也表现出很大优势.近几年陆续有研究报道部分单原子材料具有多种酶的活性.具有多酶活性的单原子材料目前已广泛应用于抗菌、癌症治疗以及细胞保护等领域,能更好地满足实际应用的需求.Zhao等[81]利用高温气体迁移策略将Fe单原子锚定在N掺杂的纳米材料上,合成了以Fe-N4为活性位点的单原子类酶材料.该Fe基单原子材料具有良好的类过氧化物酶、氧化酶和过氧化氢酶活性.Lu 等[82]采用主客体封装策略,通过一锅机械化学方法制备Fe-N-C 单原子类酶材料,无需溶液步骤和酸碱处理.合成得到的Fe-N-C具有类过氧化物酶、氧化酶、过氧化氢酶和谷胱甘肽过氧化物酶活性.
在单原子类酶材料活性中心中,引入另外一个金属原子与其相邻,也是一个调控催化活性的重要策略.双金属原子作为活性中心,可以继承单原子活性中心暴露、原子利用效率大的优势,同时也能整合双金属的功能,利用其潜在的协同作用[29,31].在自然界中,很多天然酶也具有双金属核心,如细胞色素c氧化酶、多酚氧化酶等具有双铜核心,在催化过程中金属二聚体的结构有利于O2或过氧化物的吸附,促使O—O键断[83~85].Jiao等[86]合成以Fe-Fe双位点为核心的双金属催化剂Fe2NC,Fe2NC具有典型的氧化酶活性(细胞色素c氧化酶、NADH氧化酶和抗坏血酸氧化酶)和过氧化物酶活性(NADH过氧化物酶和抗坏血酸过氧化物酶),其类酶活性显著好于以单原子Fe 为位点的Fe1NC.同时理论计算表明,由于双铁位点分别吸附O2或过氧化物中的2个O,可以延长O—O键的距离,加速O—O裂解,从而提高类酶催化活性.
特定的生物催化过程往往需要多酶的协同作用才能完成.生物系统中的多酶级联反应可以高效率、有条不紊地进行,主要得益于以下几点:首先可以减少反应中间体在酶之间的扩散,提高中间体的利用率[87];其次,在有限的空间内提高催化底物的局部浓度、加速传质[88].受到天然酶级联催化的启发,构建集成多酶活性于一体的单原子材料,串联不同的反应路径并按照次序进行,可以极大提高反应效率和反应产率.Zhu等[89]报道了Mn基单原子类酶材料,该材料表现出类过氧化氢酶、氧化酶和过氧化物酶活性,可以利用癌细胞环境中过量的H2O2发生级联催化反应.首先,其过氧化氢酶功能催化H2O2的分解,在细胞中产生大量的O2.随后,其类氧化酶功能可以介导电子转移到O2,从而产生丰富的.同时,其类过氧化物酶活性可催化H2O2分解生成·OH.两种活性氧的产生诱导肿瘤细胞发生凋亡.Yan等[90]在应对颅脑创伤引发的神经炎症时,开发了以CeO2团簇为载体负载单原子Pt的类酶材料(Pt/CeO2),Pt单原子的引入诱导CeO2(111)发生晶格膨胀.Pt/CeO2具有类超氧化物歧化酶、谷胱甘肽过氧化物酶、过氧化氢酶和过氧化物酶多酶活性,且相较于CeO2,四酶活性显著提高.基于颅脑创伤诱发产生的过量活性氧,Pt/CeO2多酶级联反应有显著的清除效果.如,Pt/CeO2的类超氧化物歧化酶活性可以歧化生成H2O2和O2,Pt/CeO2进一步利用类谷胱甘肽过氧化物酶、过氧化氢酶、过氧化物酶活性代谢产生的H2O2,以达到减轻炎症的治疗效果.
2 单原子类酶材料在生物医学方面的应用
2.1 癌症治疗
癌症发病率高且治疗难度大,严重危害着人类生命健康.利用具有类酶活性的纳米材料来进行纳米催化治疗是一种新兴的治疗癌症的技术,具有制备简单、成本低及稳定性高等优势.目前,基于纳米酶的治疗方法主要包括化学动力学治疗(CDT)、光热治疗(PTT)和光动力治疗(PDT)等[91~93].上述治疗过程中,纳米酶通过催化O2或H2O2产生高毒性的活性氧(如,1O2或·OH),或通过光照射光敏剂来产生活性氧,引发肿瘤细胞凋亡或坏死.但纳米酶在肿瘤微环境中催化活性仍然不足,单原子材料活性位点暴露量大、催化活性高,且金属离子泄漏少、副作用小,在癌症治疗方面引起了广泛关注.
CDT 是根据肿瘤微环境弱酸性、H2O2过量的特点,通过芬顿反应或类芬顿反应,催化H2O2产生·OH来杀死肿瘤细胞的一种治疗方法.Huo等[93]利用ZIF-8锚定单原子Fe,高温热解后修饰聚乙二醇(PEG),合成了活性中心是Fe-N4的单原子类酶材料(PSAF NCs).给小鼠静脉注射PSAF NCs 后,由于肿瘤的高通透性和滞留效应(EPR 效应),PSAF NCs 会聚集在肿瘤病灶内.基于弱酸性和H2O2过量的微环境,PSAF NCs会原位催化H2O2生成毒性较强的·OH,抑制肿瘤生长.同时,产生·OH可以诱导脂质过氧化物积累,进一步诱导肿瘤细胞铁死亡,协同增强了治疗效果[图5(A)].然而,在CDT治疗中大量产生的高毒性活性氧,在杀死肿瘤细胞的同时也会损伤正常细胞.如何有选择性地杀死肿瘤细胞是一个关键问题.Wang等[94]采用金属离子与有机配体自组装的策略,合成了具有原子级分散Cu位点的六亚铁酸铜类酶材料(SSNEs).SSNEs具有类谷胱甘肽氧化酶和类过氧化物酶双重酶活性.SSNEs作为类谷胱甘肽氧化酶,通过消耗谷胱甘肽将CuⅡ转化成CuⅠ,CuⅠ的存在增强了SSNEs的类过氧化物酶活性,催化H2O2产生大量的·OH 并诱导肿瘤细胞凋亡.由于肿瘤细胞内谷胱甘肽水平远远超过正常细胞,因此,SSNEs可以选择性地在肿瘤环境中实现治疗.
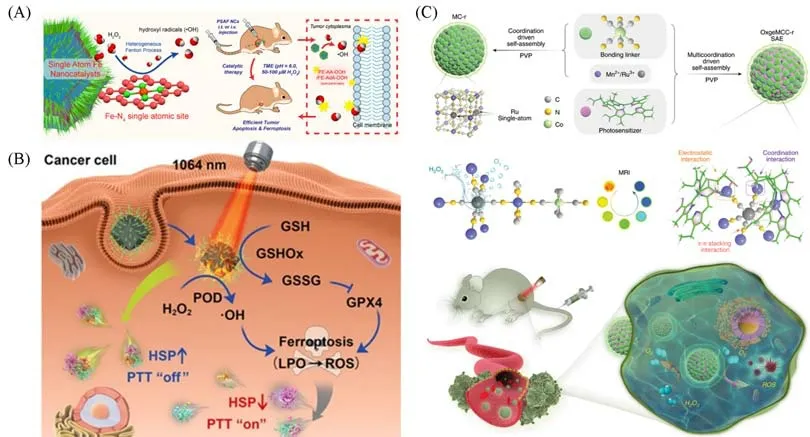
Fig.5 Single⁃atom materials for cancer treatment
PTT是在激光照射下,光热转换材料利用光热转化产生的热量来杀死癌细胞的一种治疗方法.与化疗、放疗等传统治疗手段相比,PTT更高效、精准和可控[35,45].Su等[95]采用氮掺杂的介孔碳纳米球负载铁单原子,合成了Fe 单原子催化剂(SAFe-NMCNs).SAFe-NMCNs 可以模拟过氧化氢酶和过氧化物酶双重酶的活性.一方面,SAFe-NMCNs 作为类过氧化氢酶,能够催化H2O2生成O2,实现在肿瘤部位的超声成像;另一方面,SAFe-NMCNs 作为类过氧化物酶,能够催化H2O2生成·OH 并诱导肿瘤细胞凋亡.此外,SAFe-NMCNs具有优异的光热性能,能够实现光热增强的肿瘤治疗效果.然而,在PTT治疗中往往也存在一些问题.如,过多的热量无法避免地会损伤肿瘤周围的正常组织,但温和的热量对癌细胞造成的损伤也容易被热休克蛋白(HSPs)修复.如何控制温度使得PPT效率最大化,又能减少对正常组织的伤害具有重大意义.Chang等[96]用ZIF-8负载Pd纳米颗粒,高温裂解得到氮配位的碳载Pd单原子类酶材料.该材料具有类过氧化物酶、谷胱甘肽氧化酶活性,且具有较好的光热转化效能.该单原子材料的双重类酶活性可上调脂质过氧化物酶(LPO)和活性氧水平,实现热休克蛋白(HSPs)的失活并诱导铁死亡,从而达到温和的光热治疗效果[图5(B)].
PDT治疗是通过特定波长的光照射肿瘤组织中的光敏剂,从而产生活性氧来杀死肿瘤细胞的一种治疗方法.Wang 等[79]通过自组装策略,将金属离子、有机配体和Ce6 光敏剂封装到聚乙烯吡咯烷酮(PVP)聚合物中,合成了Ru单原子催化剂(OxgeMCC-r).在肿瘤细胞H2O2过表达的情况下,OxgeMCC-r催化H2O2原位生成O2,缓解肿瘤微环境的缺氧状态,同时产生高细胞毒性的1O2,诱发肿瘤细胞凋亡[图5(C)].在实际的治疗过程中,由于恶性肿瘤具有多样性、复杂性和异质性,单模态的治疗方法效率低、治疗效果有限.Du 等[97]采用卟啉基聚合物锚定Pd 单原子,合成了以Pd-N2-Cl2为活性中心的单原子催化剂(Pd-Pta/Por),可用于化学动力、声动力和光动力三模态治疗.首先,Pd-Pta/Por具有优异的过氧化物酶活性,可催化H2O2生成·OH以用于化学动力治疗.其次,卟啉基的声敏剂和光敏剂在超声或激光照射下产生1O2的效率显著提高.因此,Pd-Pta/Por可以有效实现三模态联合治疗.
2.2 抗氧化治疗
败血症是一种世界范围内的高发病率、高死亡率的多器官功能障碍,局部感染可导致活性氧和氮物质(RONS)的过度产生,诱发炎症.Cao等[99]针对败血症的治疗设计了一种抗氧化纳米材料,即以卟啉Co为核心的Co基单原子类酶材料(Co/PMCS).该材料表现出类超氧化物歧化酶、过氧化氢酶和谷胱甘肽过氧化物酶的功能,可以有效消除和H2O2,同时通过氧化还原循环来去除·OH.·NO还可以通过与卟啉Co形成亚硝基金属络合物得到清除[图6(A)].此研究结果证实了该Co基单原子材料可以显著缓解小鼠全身炎症反应.
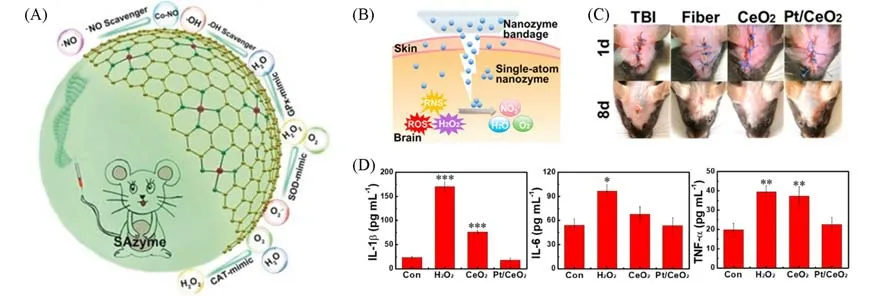
Fig.6 Single⁃atom materials for antioxidant therapy
外伤性脑损伤是一种高发病率和高死亡率的疾病.颅脑损伤后会产生许多RONS(如,·NO,·OH),从而导致一系列的生物和化学反应,造成严重的神经元损伤.Yan等[90]合成的Pt/CeO2单原子绷带,其多酶催化活性比CeO2高3~10倍,清除RONS 的能力比CeO2高2~10倍[图6(B)].体内体外研究表明,Pt/CeO2单原子基绷带可明显修复神经创伤[图6(C)],且炎症细胞因子表达同时减少[图6(D)],星形胶质细胞和小胶质细胞的聚集减轻.根据研究结果,Pt/CeO2绷带可减轻炎症反应,降低氧化应激指标,改善受损神经.
2.3 抗 菌
由细菌感染引发的一系列疾病已经对全球公共卫生构成了巨大威胁[100].传统的抗生素治疗价格高昂,具有生态毒性,而且抗生素的频繁使用会导致细菌耐药性提高[101].由于部分纳米酶具有优异的类氧化酶或类过氧化物酶活性,能够催化H2O2或活化O2产生活性氧,所以部分纳米酶可以用作新型的抗菌剂.单原子类酶材料具有催化活性高、金属离子泄漏少和生物安全性好等优势[102],可以广泛用于抗菌.Xu 等[69]合成的具有Zn-N4卟啉结构的单原子类酶材料(PMCS),表现出优异的类过氧化物酶活性,可以促进H2O2的均裂产生·OH[图7(A)].制备的PMCS具有良好的抗菌活性,可抑制铜绿假单胞菌的生长,且抑制率高达99.87%.在铜绿假单胞菌感染小鼠的模型中,经PMCS和H2O2处理的小鼠在第6天伤口创面完全愈合,与组织学染色结果一致[图7(B)].Huo等[101]合成了以氮掺杂的碳为基底的Fe 单原子催化剂(SAF NCs).SAF NCs 具有显著的类过氧化物催化活性和光热转化性能,在生理水平的H2O2(50×10−6~100×10−6mol/L)存在下,产生大量的·OH,对革兰氏阳性菌(金黄色葡萄球菌)和革兰氏阴性菌(大肠杆菌)具有优异的清除效果.丰富的·OH结合SAF NCs的本征光热性质,通过808 nm近红外激光照射,可以有效地增强杀菌效果.在感染金黄色葡萄球菌和大肠杆菌的小鼠模型中,经SAF NCs,H2O2处理和808 nm激光照射的小鼠在第5天伤口消毒效果明显优于其它治疗组.
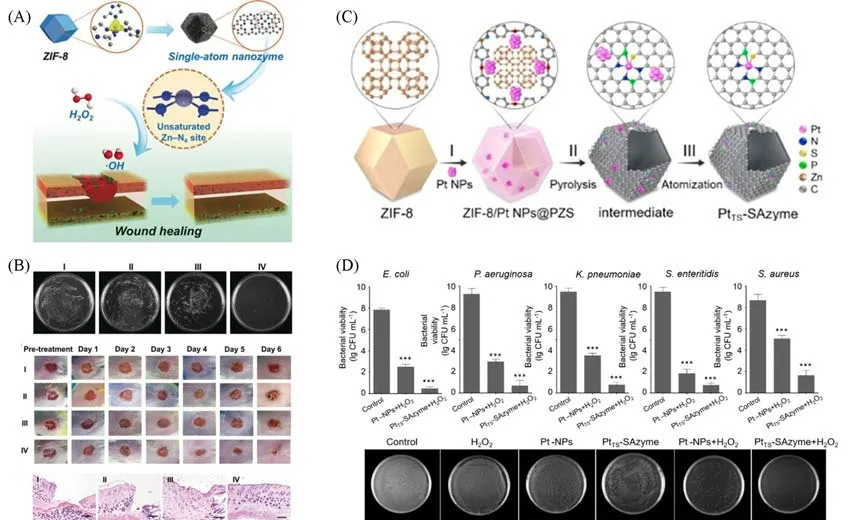
Fig.7 Single⁃atom materials for bacterial disinfection
最近,Chen等[52]首次报道了一种逆转烧结的策略.将Pt纳米颗粒直接原子化,形成热稳定的Pt单原子类酶材料(PtTS-SAzymes)[图7(C)],其具有显著的类过氧化物酶活性,远超过了Pt纳米颗粒.理论计算表明,P和S掺入促进了Pt团簇到Pt单原子的原子化过程,而且P的给电子作用和N,S的吸电子作用使得Pt单原子具有独特的电子空间结构,有利于提高催化活性.利用PtTS-SAzymes优异的类过氧化酶活性,他们还探究了其对大肠杆菌、铜绿假单胞菌、肠炎沙门氏菌、肺炎克雷伯菌和金黄色葡萄球菌这5种代表性细菌的抗菌效果.在H2O2存在的情况下,PtTS-SAzymes对大肠杆菌、铜绿假单胞菌、肠炎沙门氏菌和肺炎克雷伯菌的抑菌率为90%,对金黄色葡萄球菌的抑菌率为81%,抑菌效果远远好于Pt纳米颗粒[图7(D)].
2.4 生物传感
单原子材料原子级分散的活性位点对生物分子的响应具有高灵敏度和选择性.目前,单原子材料作为一种新兴高效的类酶材料,在生物传感领域中显示出极大的潜力[103].基于单原子类酶材料的生物传感器已成功应用于检测H2O2和释放H2O2的小生物分子(如葡萄糖、胆碱等),还原性小生物分子(如抗坏血酸、生物硫醇等),酶活性,金属离子(如Cr5+等)和肿瘤标记物等.Jiao等[50]利用Fe-N-C单原子类酶材料来检测Hela细胞产生的H2O2.由于Fe-N-C具有良好的类过氧化物酶活性,在目标物H2O2存在下,可以诱导TMB发生变色反应,实现了对H2O2的直接检测.H2O2是许多氧化酶催化底物氧化时的两电子产物,如葡萄糖氧化酶、黄嘌呤氧化酶和胆碱氧化酶.因此,通过构建级联反应可以对反应过程中释放H2O2的生物分子进行信号放大检测.Cheng等[104]合成的Fe单原子催化剂(CNT/FeNC)具有显著的过氧化物酶活性.通过构建葡萄糖氧化酶和CNT/FeNC之间的级联反应成功实现了对葡萄糖的比色检测[图8(A)].葡萄糖氧化酶催化葡萄糖产生H2O2,进一步通过CNT/FeNC 的催化作用使TMB变色.Wang 等[70]通过构建黄嘌呤氧化酶和MoSA-N3-C 单原子类酶材料之间的级联反应体系,实现了对于黄嘌呤的检测.Wu等[105]通过构建乙酰胆碱酯酶、胆碱氧化酶和Cu-N-C单原子类酶材料之间的三酶级联反应体系,实现了对乙酰胆碱和有机磷农药的比色检测[图8(B)].对于易于被氧化的还原性生物分子,可以直接通过氧化还原反应进行灵敏检测.Cheng等[104]将合成的具有类过氧化物酶活性的Fe单原子催化剂涂覆在试纸上,随后滴加H2O2和TMB的混合物,TMB由无色变为蓝色,然后利用抗坏血酸的还原作用,将氧化态的蓝色TMB还原为无色,实现对抗坏血酸的直接检测.单原子材料的类过氧化酶活性与催化环境的pH紧密相关,大多数单原子材料在酸性条件下能发挥最优的催化性能.Jiao等[63]通过乙酰胆碱酯酶催化硫代乙酰胆碱生成乙酸来影响催化环境的pH值,进一步调控FeBCN类过氧化物酶活性,从而实现对乙酰胆碱酯酶活性的评价.
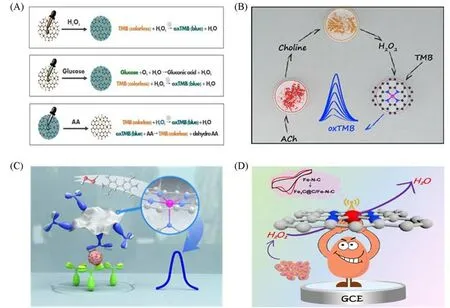
Fig.8 Single⁃atom materials for biosensing
单原子类酶材料的比色传感还被应用于检测金属离子、肿瘤标记物等方面.Mao等[106]将Fe单原子锚定到氮掺杂的石墨烯单层上(SA-Fe-NG),制备的SA-Fe-NG具有类过氧化物酶活性,可以催化H2O2,使得TMB显色.而8-羟基喹啉(8-HQ)是一种TMB的氧化抑制剂,利用检测物Cr(Ⅵ)与8-HQ之间的相互作用,使得原本被抑制的TMB恢复显色作用,从而对实现Cr(VI)的检测.Wu等[107]通过合成具有氰基缺陷的氮化碳纳米颗粒(DCN),构建了能够区分多种金属离子的阵列传感器.通过利用氰基与不同金属配位能力不同,DCN表现出不同程度的类过氧化物酶活性,从而实现了对多种金属离子的区分.酶联免疫吸附试验(ELISA)是一种目前应用十分广泛的免疫检测方法,是临床诊断中检测各种生物标志物的“金标准”.基于单原子材料的酶联免疫比色传感也有相关报道[108].Xu等[65]基于Fe基单原子材料的类过氧化物活性,构建了一种“三明治”式的双抗夹心的酶联免疫检测方法,用于比色检测癌胚抗原[图8(C)].与商业化的酶联免疫传感器(HPR 基ELISA)相比,该Fe 基单原子传感器具有更低的检测限.
上面讨论是基于单原子类酶材料的比色传感器,是利用变色底物吸光度值的变化实现分析物的灵敏检测.此外,通过生物传感技术将被分析物的浓度转化为电信号[109]、化学发光信号[110]进行检测,具有简单、灵敏以及特异性高的特点.基于Co-MoS2单原子类酶材料对H2O2的催化有显著的循环伏安响应,Wang等[71]建立了一种灵敏检测H2O2的电化学生物传感器.与文献报道的H2O2电化学检测方法相比,该电化学生物传感器的线性响应范围为50~7.241 mmol/L,检测限为10 nmol/L.Wei 等[109]通过将Fe单原子位点与碳封装的Fe3C晶体耦合,合成了Fe单原子类酶材料(Fe3C@C/Fe-N-C).Fe3C@C/Fe-N-C具有优异的类过氧化物活性,并对H2O2的催化有灵敏的电化学响应,实现了对活细胞释放的H2O2的灵敏监测[图8(D)].目前,单原子类酶材料也被用于化学发光传感器来实现对目标物的分析检测.Wang等[110]通过将Cu2+固定在氧化石墨烯表面,合成了原子级分散的Cu2+-GO,该材料具有类过氧化物酶和类烟酰胺腺嘌呤二核苷酸(NADH)过氧化物酶活性.Cu2+-GO类酶材料在H2O2的存在下,可以催化鲁米诺产生化学发光信号,还能在H2O2存在下催化多巴胺和NADH的氧化,在此基础上实现了对H2O2的灵敏检测.
3 总结与展望
近年来,单原子材料由于原子利用率高、活性位点清晰明确,且具有与金属酶相似的配位结构等多重优势,推动了新一代纳米酶的发展和新型高效生物催化剂的开发.本文总结了近几年单原子催化剂在模拟酶催化活性方面的研究进展,重点讨论了单原子材料类酶催化活性的调控策略(调控金属活性中心类型、提高单原子负载量、调控中心原子配位环境、引入官能团或协同催化组分和载体-活性中心相互作用等).此外,本文还总结了单原子类酶材料在生物医学方面的具体应用,主要包括癌症治疗、抗氧化治疗、抗菌和生物传感.
虽然单原子类酶材料领域已取得重大进展,但仍面临一些挑战.(1)特异性不足.天然酶的活性中心是由金属中心与邻近原子配位形成,且周围存在重要的氨基酸对底物的识别和活化起重要作用.单原子材料缺少像酶一样的三维空间结构,单原子的载体对底物没有识别位点,导致单原子类酶材料特异性不足.通过杂原子掺杂、载体调控、特定基团修饰以及与天然酶整合等策略可以进一步提高单原子类酶材料的特异性.同时,具有多酶活性的单原子材料在实际应用中同样具有很大优势,可以拓展具有单酶活性材料的应用场景,尤其在生物医学和复杂生命分析领域中具有不可代替的作用.(2)单原子材料的类酶活性仍有待提升.虽然单原子材料原子利用率高,但金属负载量、活性位点密度仍然不够.通过开发新的合成策略(如缺陷工程策略、协同催化策略和空间限制策略)来提高活性位点密度,可以有效地提高催化活性.此外,可以着重利用金属-载体相互作用和几何/电子效应,实现对金属原子配位环境的调控,这为提升单原子材料本征催化活性提供了机会.(3)单原子材料的类酶新活性还有待开发.目前报道的单原子材料模拟酶的类型还主要集中于氧化还原酶和水解酶两种,对于转移酶、裂合酶、连接酶和异构酶等其它类酶活性的研究还鲜有报道,合理设计和开发新型单原子材料并发掘其类酶新特性尤为重要.(4)单原子类酶材料在实现仿生催化的同时,也表现出纳米材料本身的功能性,如部分单原子材料具有优异的光热性能、光电性能及磁学性能,可以突破天然酶在应用中的桎梏,完成天然酶力不能及的工作.充分利用单原子材料自身特性,可以在分析新方法构建等方面为实现高灵敏的生物传感提供解决方案.单原子类酶材料可以结合光电化学、表面增强拉曼光谱等其它传感模式.还可以与免疫传感结合,开发便携式检测设备.(5)目前,单原子类酶材料的催化行为、催化机制具有多样性,DFT理论计算难以涵盖不同类型的催化机理.尤其是当单原子材料应用于体内诊断和治疗时,由于材料本身性质和复杂的生物环境等因素,很难建立起体内催化机制.通过将原位表征技术与计算建模以及其它人工智能技术结合起来,可以为深入理解反应机制指明方向.(6)单原子类酶材料在临床医学应用中处于起步阶段,仍然面临着很多挑战,如单原子材料生物相容性差、具有一定的毒性和不良的药代动力学,无法大规模合成等.开发新的表面工程技术和大规模合成策略,对提高生物相容性和促进临床转化具有重大意义.
