Study on Exogenous Ethylene Induced Rice Resistance to Rhizoctonia solani
2022-06-25YangShuoZhongQingyanXuXiaofengYangMingxiuLiuLianfuShuYuweiWangMengMukadasiRexitiYangSongrunandZhangJunhua
Yang Shuo, Zhong Qing-yan, Xu Xiao-feng, Yang Ming-xiu, Liu Lian-fu, Shu Yu-wei, Wang Meng, Mukadasi•Rexiti, Yang Song-run, and Zhang Jun-hua
College of Agriculture, Northeast Agricultural University, Harbin 150030, China
Abstract: Rice sheath blight is one of the main diseases in rice production in China, which can make rice unable to absorb and utilize nutrients, and has a serious impact on rice yield and quality. In this study, exogenous ethylene was used to induce rice resistance against rice sheath blight, aiming at exploring a new environment-friendly control method of rice sheath blight. The results showed that within a range of certain concentrations, ethylene had no significant effects on mycelium growth, but it could induce resistance to sheath blight in rice. The optimum concentration was 0.2 mmol • L-1 and the relative control was 86.17%. It was found that ethylene could effectively increase the activities of peroxidase (POD), phenylalanine ammonia-lyase (PAL), β-1, 3-glucanase and reduce the contents of malondialdehyde (MDA), which could enhance the resistance of rice against Rhizoctonia solani. In addition, qRT-PCR detected the expressions of rice defense genes, which indicated that the expressions of the POX, PAL and OsPR1b genes were up-regulated.
Key words: rice sheath blight, exogenous ethylene, induced disease resistance
Introduction
Rice sheath blight, caused byRhizoctonia solani(Wiwattanapatapeeet al., 2007), is one of the major diseases in the world (Bhaskaret al., 2020). At present,there are many methods to control rice sheath blight,including physical control (Qinet al., 2013), chemical control (Zhanget al., 2018) and biological control(Someyaet al., 2005).
Induced resistance of plant is the phenomenon that plants develop resistant against harmful pathogens after being exposed to external factors. There are two types of resistance: locally acquired resistance and systemic acquired resistance (Dong, 2001). Several studies showed that these types of disease resistance can activate the plant defense system, such as the synthesis of immune substances, pathogenesis-related proteins, lignin and phenols, and thus can induce this characteristic (Colson-Hanks and Deverall, 2000).Induced resistance can change the activities of defenserelated enzymes in plants, mainly including polyphenol oxidase (PPO), peroxidase (POD) and phenylalanine ammonia-lyase (PAL). For example, when plants are infected by pathogens again, the activities of enzymes,such as POD, superoxide dismutase (SOD) and PAL in the plants, are usually increased.
Plant hormones are commonly present in the plants,and they play a regulatory role when plants are under stress (Yanget al., 2013). When the plant is infected with the pathogen, it mobilizes the hormonal molecules to make a series of reactions to regulate the defense related genes, and also plays a significant role in signal transduction and regulation (Robertet al., 2011).Through continuous researches, scientists found that salicylic acid (SA), jasmonic acid (JA) and ethylene(ET) can efficiently complete the task of plant signaling system which leads to the disease resistance(Robertet al., 2007; Lópezet al., 2008; Grant and Jones, 2009). These hormones can activate related kinases, pathogenesis related (PR) genes and even transcription factors (Delteilet al., 2010). Among the three hormones mentioned above, the SA effectively resists pathogens and it is proven in many studies,while the JA and ET also effectively resist pathogens,but very few is known about them so there is need of extensive studies to find the role of ET in plant defense system (Bari and Jones, 2009).
Ethylene is basically a growth hormone, which plays a vital role in the growth, development and maturation of plants (Liuet al., 2018). Under stress conditions and pathogen infections, the ET content increases,which stimulates the defense related response of plants and improves the resistance of plants (Luoet al.,2014). 1-methylcyclopropene (1-MCP) can bind to ethylene receptor protein in plants, thus inhibiting the binding of receptors to ethylene and reducing the sensitivity of plants to ethylene. Many studies have shown that 1-MCP can be used as an ethylene inhibitor in agricultural production (Yueet al., 2019; Salazaret al., 2021; Zhanget al., 2021).
Due to the lack of resistant varieties, chemical methods are widely used to control rice sheath blight,but this method is costly, and if used for a long time and improperly, the pathogen will be resistant to chemicals, thus causing harm to humans, livestock and the environment. Therefore, this study used exogenous ethylene to induce rice defense related response and analyze the changes of resistance to rice sheath blight. In this study, the defense-related enzyme activity and gene expression were examined,in order to provide a new method and theoretical basis for the prevention and control of rice sheath blight in the future.
Materials and Methods
Effects of ethylene treatment on mycelium growth of Rhizoctonia solani
The PDA media withR. solaniplate (Φ=5 mm) in the middle were opened and turned upside down on the separator of the dryer. Controlled the concentration of ethylene in the dryer to 0.05, 0.1, 0.2 and 0.4 mmol • L-1,respectively. Each treatment contained three replicates,without ethylene as the control. The colony diameters were measured after 24 and 48 h, respectively.
Effects of ethylene treatment on occurrence of rice sheath blight
The rice plants were treated for 12 h in an enclosed space with ethylene concentrations of 0.05, 0.1, 0.2 and 0.4 mmol • L-1, and then inoculated withR. solaniand moisturized. Plants without ethylene were taken as the control. Each treatment was repeated three times.After 7 days, the height of rice plants and lesion length were measured, and the relative lesion length and relative control effects were calculated.
Effects of ethylene inhibitor (1-MCP) treatment on occurrence of rice sheath blight
According to the method of Mülleret al. (2000),a certain amount of 1-MCP powder was weighed in beakers. Beakers were placed in a closed plastic account, so that the concentration of 1-MCP in the account was 0.1, 0.5, 1 and 3 µL• L-1, respectively,and plants were placed for 12 h. Then, inoculated them withR. solaniand moisturized. Only distilled water was used for the control, and each treatment was repeated three times. After 7 days, rice plant height and lesion length were measured and calculated.
Effects of ethylene on activities of defenserelated enzymes and change of MDA content in rice
The leaves were taken at 0, 12, 24, 48 and 72 h afterR. solaniinoculation. Four treatments were set up:ET+R. solani(rice treated with ethylene andR. solani), ET (rice inoculated only with ethylene),R. solani(rice inoculated only withR. solani) and the CK(untreated rice). The activities of the POD, PAL, PPO andβ-1, 3-glucanase were determined by guaiacol method, phenylalanine colorimetric method, catechol method and method of Felix and Meins (1985),respectively. The content of malondialdehyde (MDA)was determined by thiobarbituric acid method.
Effects of ethylene treatment on expression of defense related genes in rice
The relative expression levels ofphenylalanine ammonia-lyasegene (PAL),peroxidasegene (POX)andOsPR1bwere determined. Three treatments were set up: ET+R. solani(rice treated with ethylene andR. solani),R. solani(rice inoculated only withR. solani) and the CK (untreated rice). The total RNA of plants was extracted using the TRIzol (Rioet al., 2010), and cDNA was synthesized using the M-MuLV first strand cDNA synthesis kit. The required primers are shown in Table 1. The reaction system was as the followings: predenaturation at 95℃ for 3 min; denaturation at 95℃ for 5 s; annealing at 58℃ for 30 s; extension at 58℃ for 30 s and 40 cycles; the melting curve was analyzed at 95℃for 15 s, 60℃ for 1 min, 95℃ for 15 s and 60℃for 15 s. The relative expression level of the gene was expressed by the ratio of the target gene to the internal reference gene using the 2−△△Ctmethod for data analysis. All the reactions were performed three independent experiments.

Table 1 Primer sequences for qRT-PCR
Results
Effects of ethylene treatment on mycelium growth of R. solani
The results of mycelium growth are shown in Table 2.Colony diameters were measured after 24 and 48 h,and it was found that the diameters ofR. solanitreated with different concentrations of ethylene had no significant difference compared with the control.The results indicated that within a range of certain concentration, ethylene did not significantly promote or inhibit the growth ofR. solanimycelium.

Table 2 Effects of ethylene on growth of Rhizoctonia solani
Effects of ethylene treatment on occurrence of rice sheath blight
The results of the relative control effects are shown in Table 3. The relative height of rice sheath blight lesion was significantly lower than that of the control after ethylene treatment with different concentrations. The best control was achieved, when the concentration of ethylene was 0.2 mmol • L-1, and the relative control was 86.17%. When the ethylene concentration was 0.05 and 0.4 mmol • L-1, the control effect was significantly reduced. The results showed that ethylene could induce rice resistance to sheath blight in a certain range of concentration.

Table 3 Effects of ethylene on rice sheath blight
Effects of ethylene inhibitor (1-MCP) treatment on occurrence of rice sheath blight
The effects of 1-MCP on rice relative lesion length are shown in Table 4.

Table 4 Effects of 1-MCP on rice sheath blight
The relative lesion length of rice sheath blight induced by 1-MCP with the concentration of 1 µL • L-1was significantly longer than that of the control. The results indicated that the resistance of rice to sheath blight was weakened after the treatment with ethylene inhibitor in a certain range of concentration.
Effects of ethylene on activities of defense related enzymes and MDA contents in rice
In this experiment, the POD, PAL, PPO andβ-1, 3-glucanase activities and MDA contents were analyzed.The results are shown in Fig. 1A-E. The activity of the POD in plants treated with ET+R. solaniincreased rapidly, peaked at 12 h, and then began to decrease.But after 72 h of inoculation, it increased again. The POD activity of the plants treated with ET only did not increase at the beginning, but it began to increase at 12 h and reached its peak at 24 h, while the plants treated withR. solaniincreased first and then decreased. In these results, plants treated with ET+R. solanihad the overall highest POD activity. And the POD activity of rice treated with ET+R. solaniwas significantly higher than that of other treatments (Fig. 1A). Plants treated with ET+R. solanihad the highest PAL activities from the beginning. The PAL activity started to increase,and reached its peak at 48 h. Then began to decline.Plants treated with ET had the second highest PAL activity, and it surpassed PAL activity of ET+R. solaniat 72 h of treatment application. Other treatments showed lower PAL activities (Fig. 1B). Under the four treatments, the general trend of the PPO activity in rice was the same, but the difference was not significant.All of them showed a trend of rising first and then falling, and all of them reached the peak of activity at 24 h. It could be seen that the PPO activity of rice treated with ethylene was higher than that of rice without ethylene treatments (Fig. 1C). The activity ofβ-1, 3-glucanase for ET+R. solaniwas significantly higher. Its activity started to increase at the beginning and reached its peak at 12 h. After that, its activity started to decrease. All the other treatments had a similar trend. The second highestβ-1, 3-glucanase activity was observed in plants treated with ET alone,and the lowestβ-1, 3-glucanase activity was observed in the CK group (Fig. 1D). Rice plants inoculated only withR. solanihad the highest MDA content,while those treated only with ET had the lowest MDA content. The MDA content forR. solaniincreased from the beginning and reached its peak after 72 h of inoculation. These results were followed by plants treated with ET+R. solaniand the lowest MDA content was observed in plants treated with ET only.These results indicated that exogenous ethylene could alleviate the damage to the rice membrane system caused byR. solaniand reduced the contents of the MDA in rice (Fig. 1E).
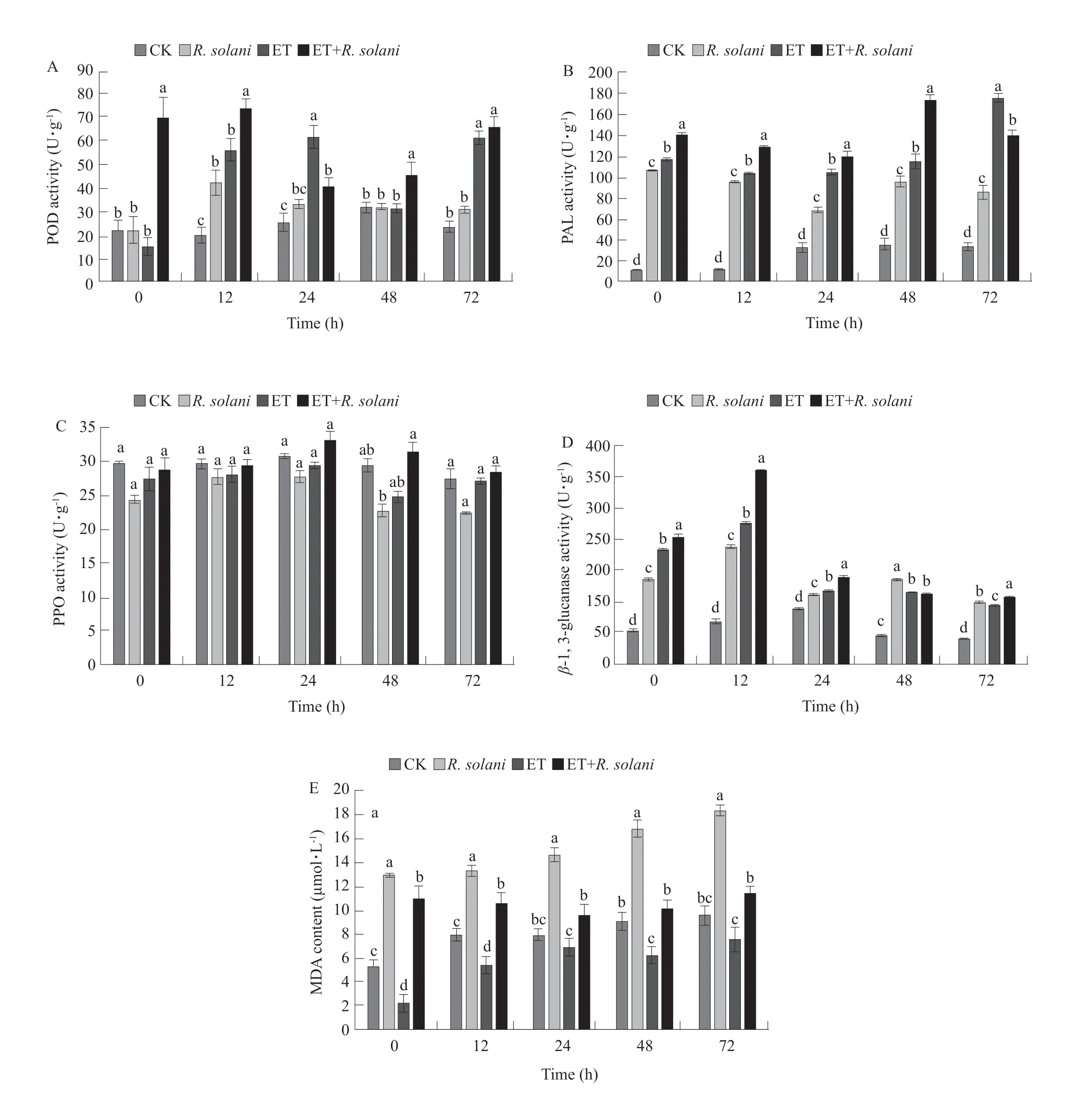
Fig. 1 Defense-related enzyme activities and MDA contents in rice after ethylene treatment
Effects of ethylene on expression of defense related genes in rice

Fig. 2 Effects of ethylene on expression of rice related defense genes
In this experiment, the expressions of some defense related genes were analyzed, which werePOX,PALandOsPR1b. The results are shown in Fig. 2A-C. In untreated rice, thePOXgene was in a stable state at each stage. ThePOXgenereached the highest level in the rice treated with ET andR. solaniat 12 h, which was 88.08% and 31.79% higher than that of rice untreated and inoculated withR. solani, respectively.The relative expression level of ricePOXgene induced by ethylene was always maintained at a high level,which was significantly different from that without ethylene (Fig. 2A). The expression of thePALgene in rice inoculated withR. solaniincreased at 12 and 24 h,which was significantly different from that of rice without inoculation. The expression of thePALgene in rice treated with ET andR. solaniwas significantly higher than that in non-ethylene-induced rice, and the highest expression was observed at 12 h (Fig. 2B). The expression level of theOsPR1bgene in the rice treated with ET andR. solanistarted to increase at the early stage of stress, which was markedly different from that in non-ethylene-induced rice. In addition, the expression level of theOsPR1bgene reached the highest level in 12 h, which was 87.3% and 49.23% higher than that of untreated rice and rice inoculated withR. solaniby 87.3% and 49.23%, respectively (Fig. 2C). These results indicated that ethylene could improve the resistance of rice to sheath blight by inducing the expressions of thePOX,PALandOsPR1bgenes.
Discussion
Ethylene, a natural gaseous hormone existing in most plants, is an important signaling molecule that controls plant growth and maturation, regulating the senescence and abscission of plant organs and accelerating fruit maturation. When plants are under stress or infected by pathogens, the contents of ethylene will increase,inducing plants to produce resistance and activate plant self-defense response. In this experiment, different concentrations of ethylene was used to treatR. solani, and it was found that ethylene had no obvious inhibitory effects on mycelium growth ofR.solani. The results of plants treated with ethylene at different concentrations showed that the relative lesion height of rice sheath blight was reduced in different degrees. The relative lesion height was the lowest, when the concentration of ethylene was 0.2 mmoL • L-1, and the relative control efficiency reached at 86.17%.The treatment of rice plants with ethylene inhibitor(1-MCP) was used to verify whether the rice resistance to sheath blight was related to ethylene or not. The results showed that the relative lesion height of rice plants treated with 1-MCP was significantly higher than that of the control. When the concentration of 1-MCP was 1 µL • L-1, the relative lesion height was the highest and the disease was most serious. Seoet al. (2011) also showed that ethylene played an important role in rice sheath blight resistance, which was consistent with the results of the present study,indicating that a certain concentration of ethylene could improve the resistance of rice to sheath blight.
The pathogen infection would increase the activities of some enzymes related to phenolic metabolism in the host, mainly including the PPO, POD and PAL. The PPO could oxidize phenols in plants into quinones that were poisonous to pathogens. It not only contributed to the accumulation of phenols in plants, but also helped to increase the content of quinones (Schlumbaumet al.,1986). The POD was able to metabolize hydrogen peroxide and promote the accumulation of lignin,which could limit the proliferation of fungi (Josephet al., 1998). The PAL participated the production of lignin and phytoalexin, and was also a key enzyme and rate-limiting enzyme for phenylpropane metabolism(Legrandet al., 1976; de Sáet al., 1992). The MDA was the final product of membrane lipid peroxidation, and was directly related to the damage degree of cell membrane.Many studies had shown thatβ-1, 3-glucanase was also involved in the defense response in plants (Schlumbaumet al., 1986; Mauchet al., 1988; Flachet al., 1993). In this study, the activities of the POD andβ-1, 3-glucanase increased slowly at the initial stage ofR. solaniinfection,then reached the peak with the extension of infection time, and began to decline. The activity of the PAL decreased and then increased. This was because the activities of the POD andβ-1, 3-glucanase were more easily activated byR. solani. After exogenous ethylene was applied, the MDA contents in rice were significantly decreased. The MDA was an important product of membrane lipid peroxidation, which played an important role in resisting pathogen infection (Suet al.,2016). Its accumulation would accelerate membrane destruction. This indicated that application of exogenous ethylene could reduce the damage ofR. solanito rice membrane system, and reduce the contents of the MDA.After ethylene induction, the contents of the PPO had no obvious changes, which might indicate that ET had no obvious induction effects on plant PPO activity.These results indicated that ethylene could improve the disease resistance of plants by increasing the activities of defense-related enzymes.
When rice was under external stress or infected by pathogens, it would induce its own defense system,thus promoting the expression of the PR genes. The expression of the PR genes could improve the disease resistance of rice, and it was often used as a characteristic molecular marker in researches related to rice disease defense. In this experiment, thePOX,PAL,andOsPR1bgenes were selected. After treatment with 0.2 mmoL• L-1ethylene, the expression levels of theOsPR1b,PALandPOXgenes were significantly higher than those without ethylene treatment. The result was consistent with the report of Massoloet al.(2011) that ethylene significantly increased the expressions of thePALandPOXgenes in fruits. These results indicated that ethylene could enhance the self-defense ability and resistance of plants by increasing the expressions of defense-related genes, such asOsPR1b,PALandPOX.
Conclusions
The mechanism of ethylene induced resistance to sheath blight of rice was systematically studied at physiological, biochemical and genetic levels. The results showed that after treatment with ethylene for 24 and 48 h, there was no significant inhibition on the mycelial growth ofR. solani. The appropriate concentration of ethylene could improve the resistance and alleviate the damage of rice to sheath blight.Among them, the best concentration of ethylene induced treatment was 0.2 mmoL• L-1, and the relative control efficiency was 86.17%. After rice was treated with 0.2 mmoL• L-1ethylene, the activities of the PAL, POD andβ-1, 3-glucanase were significantly increased in rice, but the change of the PPO activity was not significant. At the same time, the contents of the MDA in rice were decreased. These results indicated that proper concentration of ethylene could regulate the activities of defense enzymes related to rice disease resistance, participate in the plant defense reaction and enhance the resistance of rice to sheath blight. The expression levels of thePAL,OsPR1bandPOXgenes in rice after induction were significantly higher than those of the genes without induction treatment. These results indicated that ethylene actively participated in the induction and regulation of defense related genes, during the interaction between rice and sheath blight, which produced a series of disease-resistant mechanisms and enhanced the defense ability of rice against sheath blight.
杂志排行

Journal of Northeast Agricultural University(English Edition)的其它文章
- Research on Automatic Diagnostic Technology of Soybean Leaf Diseases Based on Improved Transfer Learning
- Difference Analysis of Liver Metabolic Response Between Diploid and Triploid Rainbow Trout Oncorhynchus mykiss Under Fishing Stress
- Attribution of Antioxidation of Quercetin in Vitro and Arbor Acre Broilers
- Effects of Encapsulated Enzyme and Yeast Products on in Vitro Rumen Fermentation
- Effects of Planting Density and Cutting Time on Hay Yield and Nutritional Value of Forage Soybean HN389
- Measurement of Grain Production Efficiency in Main Grain-producing Areas and Analysis of Inter-provincial Differences
—— A Study Based on Super-SBM Model and Malmquist Index