黄芩对营养性肥胖大鼠血糖血脂的作用及其机理研究
2022-06-17曾治君凌美婷朱鑫辉刘红宁严小军吉燕华江西中医药大学中医基础理论分化发展研究中心江西省中医病因生物学重点实验室南昌330004
★ 曾治君 凌美婷 朱鑫辉 刘红宁 严小军 吉燕华(江西中医药大学中医基础理论分化发展研究中心/江西省中医病因生物学重点实验室 南昌 330004)
肥胖已经成为日常生活中的常谈话题[1],一定程度上对人民的精神造成了影响[2],肥胖也成为糖尿病发病的独立危险因素[3]。为此,如何有效防控肥胖成功为了大家亟待解决的问题[4]。肥胖的发生过程往往伴随糖脂紊乱[5],为此,从药物的开发角度,主要是针对降糖和降脂药物的开发,而西药的传统研究思路是单一靶点小分子药物的设计与验证[6],但是中药从一开始的临床应用,都是多成分多靶点的应用方式,对多种因素可以造成的肥胖来说,无疑有着巨大的调控优势。
临床常用中药黄芩始载于《神农本草经》,具有清热燥湿、泻火解毒和利尿等功效[7],常与黄连作为药对进行用药,临床研究表明,黄芩治疗2型糖尿病效果显著[8-9],具有降血糖和血脂等作用[10]。
上世纪人类基因组计划测序工作完成以来,大规模基因测序日益兴起,产生了基因组学、转录组学和蛋白组学等。后基因组学时代,基于新一代测序技术的转录组学以高通量、高灵敏和易可视化等优势得到迅速应用[10]。转录组学作为系统生物学方法之一,其整体性、系统性和综合性与中药多成分多靶点及整合调节的特点相符。目前关于黄芩防治肥胖的研究多集中在降脂降糖的临床效应上,对其差异基因的报道较少,特别是差异基因的通路上,更是鲜有报道。为了寻找黄芩降糖降脂防治肥胖引起的差异基因和其机制,本文运用高脂饲料建立营养性肥胖大鼠模型,用黄芩汤药灌胃进行干预,使其血糖血脂得到改善。借助转录组学分析方法,探讨黄芩干预营养性肥胖大鼠的降糖降脂差异基因表达,为临床治疗提供参考。
1 材料与方法
1.1 药材的制备
药材加入10倍量水浸泡1 h(浸透为准),用武火开始加热,沸腾后,改用文火(小功率)计时,40 min后,用纱布趁热过滤药液,在剩余的药渣中继续加8倍量水,按照上次煎煮工艺继续煎煮,合并两次水煎液进行旋转蒸发浓缩到终浓度为1 g原生药材/mL溶液,分装,放于-20 ℃冰箱里保存待用。
1.2 造模与给药
4周龄SD雄性大鼠24只,体质量(90±10)g,由湖南斯莱克景达实验动物有限公司提供(SCXK〈湘〉2011-0003),饲养于江西中医药大学实验动物科技中心(SYXK〈赣〉2017-0004),适应性喂养一周后,随机分为空白组8只、造模组16只,空白组给予含10%脂量的普通饲料喂养,模型白组给予含60%脂量的高脂饲料喂养,造模8周后,将模型组随机分为模型组8只和黄芩药物组8只,给药剂量为0.1 g原生药材/100 g大鼠体重,给药5周后,大鼠空腹12 h,称取体重,根据体重计算3%戊巴比妥钠用量(50 mg/kg)进行腹腔注射,在麻醉状态抽取大鼠腹腔静脉血,血液静置2 h后3 000r/min离心10 min,取上层血清,-80 ℃冰箱保存备用。
1.3 表型测定
肥胖相关的体重和体长每周采集一次,进而计算 Lee’s指数,其公式为 Lee’s指数 =[体重(g)]1/3×1 000/体长(cm)。
同时,血液样本每两周采样一次,并按照相应的血糖血脂检测试剂盒上操作流程检测,其检测仪器为全自动生化分析仪(Modular P800,瑞士罗氏)。
取肝脏约50 mg用Trizol法提取总RNA,质检合格后上BGISEQ-500平台测序(深圳华大基因科技服务有限公司)。
1.4 统计分析
2 结果
2.1 体重体长和血糖血脂变化
造模8周后,与空白组比较,造模组SD大鼠腹部圆而肥胖,体重、体长和Lee’s指数均明显增加,血脂中的总胆固醇最先开始产生差异,发生在第2周,而甘油三酯和血糖发生在第4周,其他血脂指标只有高密度脂蛋白胆固醇在第6周产生了差异,而给药5周后,黄芩药物组的体重和Lee’s指数并没有得到逆转,但是血糖水平显著下降,高密度脂蛋白胆固醇显著升高(P<0.05),而总胆固醇和低密度脂蛋白胆固醇水平均有下降趋势(P>0.05)。见表1。
表1 高脂饲料喂养8周体征和血糖血脂变化()
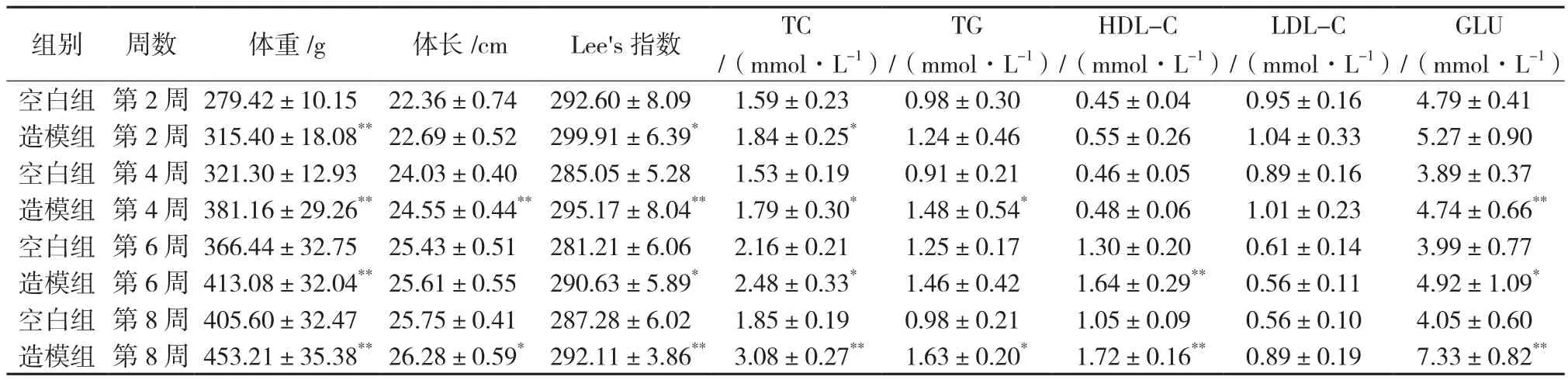
表1 高脂饲料喂养8周体征和血糖血脂变化()
注:空白组8只,造模组16只。与空白组比较,*P<0.05,**P<0.01。
GLU/(mmol·L-1)空白组 第2周 279.42±10.15 22.36±0.74 292.60±8.09 1.59±0.23 0.98±0.30 0.45±0.04 0.95±0.16 4.79±0.41造模组 第2周 315.40±18.08**22.69±0.52 299.91±6.39* 1.84±0.25* 1.24±0.46 0.55±0.26 1.04±0.33 5.27±0.90空白组 第4周 321.30±12.93 24.03±0.40 285.05±5.28 1.53±0.19 0.91±0.21 0.46±0.05 0.89±0.16 3.89±0.37造模组 第4周 381.16±29.26**24.55±0.44**295.17±8.04** 1.79±0.30* 1.48±0.54* 0.48±0.06 1.01±0.23 4.74±0.66**空白组 第6周 366.44±32.75 25.43±0.51 281.21±6.06 2.16±0.21 1.25±0.17 1.30±0.20 0.61±0.14 3.99±0.77造模组 第6周 413.08±32.04**25.61±0.55 290.63±5.89* 2.48±0.33* 1.46±0.42 1.64±0.29** 0.56±0.11 4.92±1.09*空白组 第8周 405.60±32.47 25.75±0.41 287.28±6.02 1.85±0.19 0.98±0.21 1.05±0.09 0.56±0.10 4.05±0.60造模组 第8周 453.21±35.38**26.28±0.59* 292.11±3.86** 3.08±0.27** 1.63±0.20* 1.72±0.16** 0.89±0.19 7.33±0.82**组别 周数 体重/g 体长/cm Lee's指数 TC/(mmol·L-1)TG/(mmol·L-1)HDL-C/(mmol·L-1)LDL-C/(mmol·L-1)
2.2 肝脏组织RNA-seq结果
聚类图显示,空白组和造模组以纵坐标主成分有较好的分离,而黄芩药物组基本处于二者之间。见图1。相对于模型组,黄芩干预后,上调基因有23个,下调基因有15个(P<0.05)。这些差异基因主要富集在生物节律和支链氨基酸合成代谢通路途径中,黄芩药物组Per1、Per3和Bhlhe40等生物节律相关基因上调,Sds等支链氨基酸合成代谢通路相关基因上调。见图2和表2。
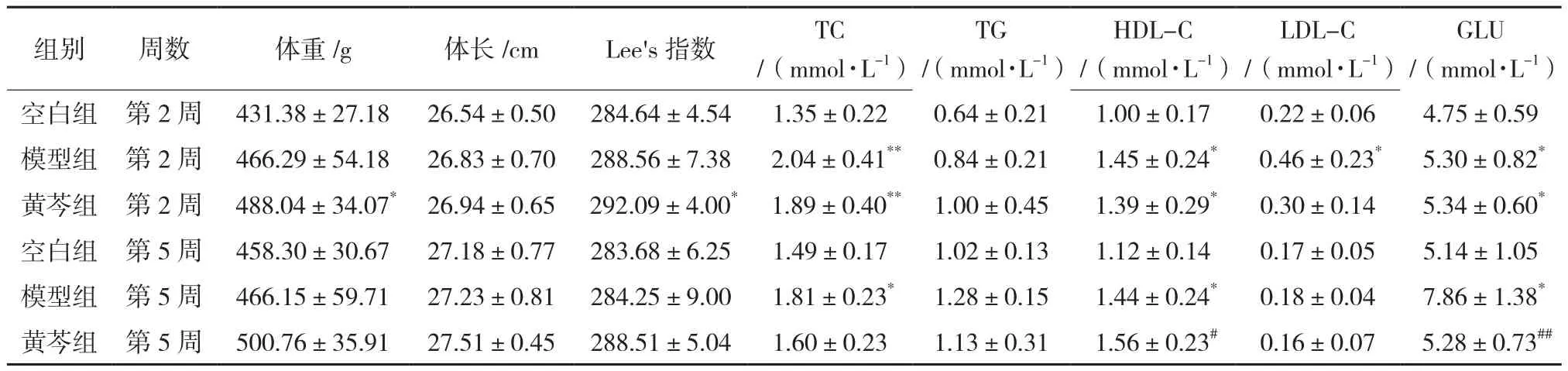
表2 给药5周体征和血糖血脂变化(n=8)
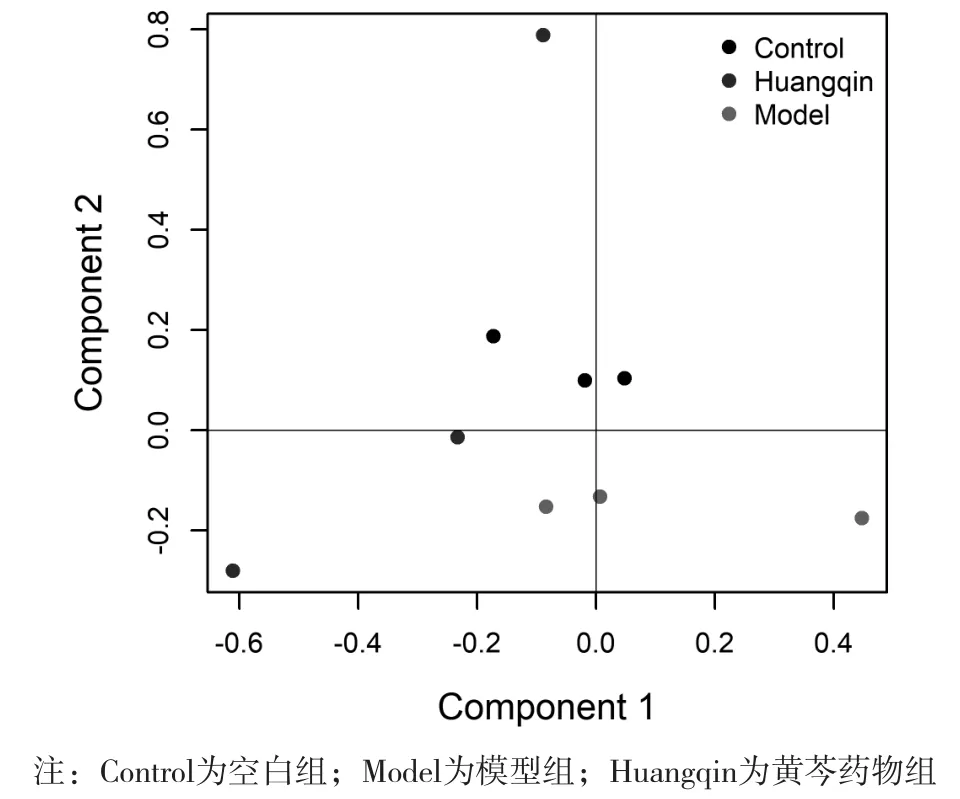
图1 肝脏组织差异基因聚类结果
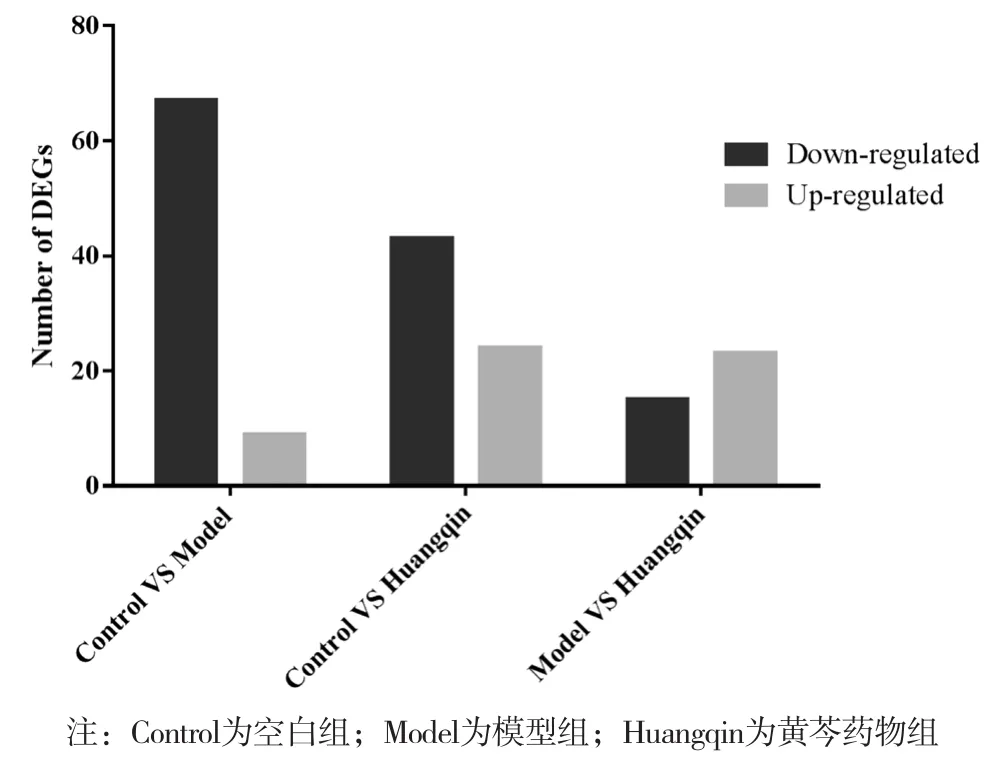
图2 肝脏组织差异基因结果

表3 黄芩对营养性肥胖大鼠肝脏差异基因的影响
3 讨论
已有很多研究团队开了肥胖的相关研究工作[12-13],尤其是近些年来,借助多组学的技术优势[14-17],在肥胖方面取得了较大的进展[18-19],然而临床常用的苦寒药在调控肥胖研究方面并未深入开展,尤其是基于新一代测序技术的转录组学差异基因网络调控方面鲜有报道。本文基于前期建立的营养性肥胖大鼠,以临床常用苦寒药黄芩进行干预来初步探讨其调控血糖血脂的作用。血清中的糖脂结果显示黄芩具有降低血糖和总胆固醇的作用,且降糖的效果优于降脂的效果,一定程度上体现了黄芩对糖脂的调控作用,与已有黄芩提取物对血糖血脂的影响相一致[20-21]。
为了更好了解黄芩对血糖血脂的调控机理,特别是哺乳动物机体最大的实质性代谢性器官肝脏中哪些基因参与了糖脂代谢过程。基于新一代测序技术的转录组学,课题组开展了肝脏组织表达谱测序,结果显示差异基因主要位于生物节律和支链氨基酸合同通路中,这与中科院梁斌研究团队报道的结果相符[3]。表明研究模型与人类肥胖和糖尿病的发生发展过程相符合,且黄芩调节的作用靶点Sds基因,即丝氨酸脱水酶,在大鼠肝脏的活性受到饮食和激素等因素影响[22]。该基因富集于支链氨基酸合成通路,表明黄芩干预营养性肥胖大鼠后,调控支链氨基酸相关的酶和基因得到了加强。相对于模型组,黄芩药物组还显著上调了生物钟相关的3个基因(Per1,Per3和Bhlhe40),表明黄芩可能通过延长时间来转化高脂饲料造成的能量过剩,进而改善血糖和血脂。这一过程可能还涉及到了支链氨基酸合成代谢通路相关基因,但相关的候选基因作用机制还需进一步验证和深入研究,本研究为黄芩及类似苦寒类药物的机制研究提供了新的研究方案。
