木质素荧光研究进展
2022-05-26申琪薛雨源杨涛伟张妍李胜任
申琪,薛雨源,杨涛伟,张妍,李胜任
(太原理工大学化学化工学院,山西 太原 030024)
木质纤维素类生物质主要由纤维素、半纤维素和木质素三部分组成,因其量大、可再生、廉价易得等特点,为全球能源消费从化石资源向可再生碳资源转移提供了巨大潜力。然而,由于结构复杂,木质素的高效利用一直以来都是生物质转化和应用过程中的瓶颈。基于木质素的独特物化性质,开拓其在相应领域的功能化应用,成为推动木质素资源化的一个重要研究方向。
近年来,基于木质素芳香结构、表面活性等独特物化性质的热点研究不断涌现,涵盖大宗化学品和燃料、精细化学品和功能材料等领域。特别地,与纤维素和半纤维素相比,木质素是一种天然高分子荧光材料。近年来,荧光技术因其灵敏度高和非破坏性检测等优势,成为细胞成像与环境检测等领域的重要方法。木质素作为一种天然荧光团,因其生物相容性好和可再生等优点,在荧光检测、示踪及传感器领域极具应用潜力。目前,木质素荧光主要应用于植物细胞壁成像,利用木质素在植物中的特异性和无标记成像方面的优势,在显微镜下观察木材或纸纤维中的木质素;部分科研工作者也探索了木质素作为荧光探针在检测金属离子、小分子化合物和细胞内pH 响应等领域的应用。木质素的荧光特性为木质素的高附加值利用提供了新颖和广阔的前景。然而到目前为止,木质素的荧光机理和调控机制并不清晰,且相关研究结果时间跨度较长,总结凝练不足,制约着这种天然荧光材料的应用前景。
为了更好地推动木质素荧光研究,本文系统地综述了木质素荧光的研究现状,并分别对其研究思路和存在的科学问题进行了总结和凝练。按照研究思路的区别,木质素荧光研究可以分为两大类:第一类是将木质素看作单一荧光团或多荧光团间的线性加和系统,通过对比不同结构模型物与真实木质素的荧光性质,筛选出潜在的荧光团;第二类则以木质素荧光团间的聚集耦合态为对象,通过研究木质素荧光的基本性质和变化规律揭示其耦合作用机制,阐明荧光机理,见图1。最后,本文对木质素荧光领域的研究和应用进行了展望。

图1 木质素荧光研究的发展进程
1 木质素荧光简介
植物组织中含有大量的天然荧光分子,其最大发光波长可以覆盖整个可见光波段,其中木质素是植物中最重要的天然荧光分子之一,在紫外线下会发射出蓝绿色荧光,见图2。因其光致发光的天然属性,木质素荧光早在20世纪50 年代就被用在造纸工业的实时监控和质量检测,之后在荧光共聚焦显微镜的辅助下,原本木质素的自荧光标记更多被用于木材结构分析、植物木质化过程监控和木材/纸浆的光化学反应等植物学研究领域。荧光性质方面,木质素属于多荧光团发光体系,但由于复杂的化学结构和构型构象,其荧光团结构并不清楚,目前推测出的可能荧光团结构包括松柏醇衍生物、联苯、阿魏酸、二苯乙烯和苯基苯并呋喃等。同时,木质素荧光团之间还存在复杂的电荷和能量转移行为,这进一步增加了木质素荧光的解析难度。

图2 植物中常见荧光化合物及其发射波长[9]
2 木质素荧光机理
木质素大分子是由香豆醇、松柏醇和芥子醇三种主要前体以及少量阿魏酸、松柏醛和芥子醛等其他前体通过酶的脱氢聚合和自由基耦合而成的,其连接键包括以-O-4 为主的醚键和少量以-5、-1和5-5为主的碳碳键。木质素分子共轭程度较低,但其荧光波长可以覆盖近紫外和整个可见光波段,其化学结构与整体荧光发射波长不匹配,无法合理解释木质素的长波长荧光来源。为此,通过对比模型物与真实木质素的荧光性质或分析木质素在定向处理前后的荧光变化规律,探寻木质素中可能存在的大共轭结构,是木质素荧光研究的主要内容。本节将从木质素荧光团化学结构的筛选和木质素荧光团间聚集耦合态的研究出发,讨论木质素荧光研究现状。
2.1 木质素荧光团化学结构的筛选
根据木质素组成单元的结构特点,Konschin等先以单苯环模型物苯乙烯为主体,研究了甲基、甲醇基、甲氧基和酚羟基等助色团取代基对苯乙烯发光波长的影响。结果显示,苯乙烯的荧光发射波长在294nm左右,在位引入助色团对其荧光几乎没有影响;取代基在3位时,最大发光波长有微小的红移;取代基在对位时,发光波长移动最为明显,红移可达10nm 以上,其中酚羟基的助色作用最强,光谱红移达到17~18nm,但仍然无法与真实木质素荧光相提并论。在此基础上,Lundquist等较为全面地分析了不同共轭程度木质素模型物的荧光光谱,包括低共轭的单苯环和二聚体模型物,中等共轭程度的联苯、二苯醚、松柏醇衍生物、羰基/羧酸基取代的苯丙烷/丙烯模型物,以及共轭程度更高的二苯乙烯类模型物,见图3。结果显示,在荧光强度方面,二苯乙烯结构和羰基/羧酸基取代的芳基丙烷对木质素影响最大,其中二苯乙烯类模型物表现出远高于其他所有模型物的荧光量子产率,具有苯并呋喃结构的二苯乙烯类模型物更是达到0.61;羰基/羧酸基取代的芳基丙烷模型物则表现出极低的荧光量子产率,分析认为这与羰基/羧酸取代基的荧光淬灭效应有关。在发射波长方面,松柏醇衍生物和二苯乙烯类模型物的最大荧光发射波长与磨木木质素最为接近。虽然Lundquist 等筛选了大量不同结构的模型物,但仅测试了它们在二氧六环或二氧六环/水混合液[50%∶50%(体积分数)]中的荧光光谱,并未对它们的荧光性质进行深入研究。
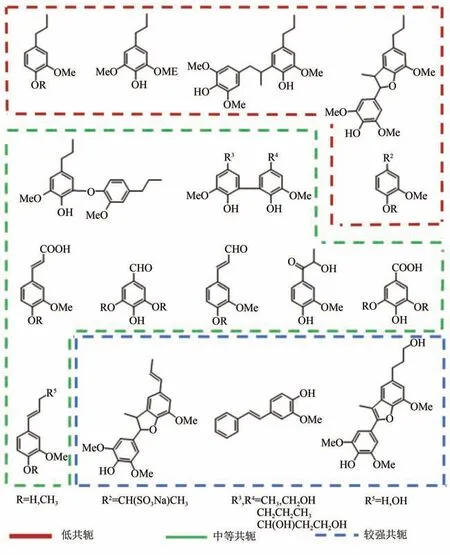
图3 不同共轭程度的木质素模型物[23]
之后,研究人员对几种重要模型物的荧光性质进行了深入分析,见图4。Lang 等继续对苯丙烯酸和二苯乙烯这两类对木质素荧光强度影响最大的结构进行研究,与Lundquist 等的不同之处是将溶剂从二氧六环/水混合液换成了甲醇。结果显示,苯丙烯酸类模型物的荧光光谱变化不大,强度较小,最大荧光发射波长仍在430nm左右。值得注意的是,虽然二苯乙烯类模型物的最大荧光发射波长仍然在390nm左右,但其在甲醇中的荧光强度急剧下降,甚至比苯丙烯酸类模型物的还要弱。文献作者并未对该现象进行讨论,本文作者认为该现象主要与二苯乙烯类模型物的聚集诱导发光效应有关。二苯乙烯类模型物属于典型的聚集诱导发光型荧光团,在二氧六环中掺入50%(体积分数)的水会使其发生聚集,从而显示出较高的荧光发射,不过作者当时并没有意识到聚集态对二苯乙烯类分子荧光的影响。Beyer 等则对-羰基取代的-O-4 型二聚体模型物的荧光性质进行了深入研究。结果显示,-O-4 连接键会阻断两个苯环之间的π-π 共轭,模型物荧光表现为两个子结构的荧光加和,其中苯乙酮结构由于羰基的淬灭效应导致荧光强度很弱,模型物分子的荧光光谱主要为愈创木酚结构的荧光发射,其最大发射波长在340nm 左右。同时,文献作者发现模型物荧光强度会随溶剂极性或质子氢浓度的增加而增大,分析认为该现象主要与两个子结构之间的分子内荧光能量转移受限有关,即当分子内荧光能量转移变弱时,苯乙酮结构对荧光的淬灭程度下降,从而导致整体荧光增强。除溶液体系外,Castellan等还报道了松柏醇、-羰基取代的-O-4型二聚体、苯基苯并呋喃和联苯二酚4种模型物在纤维素表面的固体薄膜荧光性质,其最大荧光发射波长分别约为340nm、355nm、385nm 和395nm。4 种模型物的发光波长均比溶液状态有所增加,这主要与模型物的聚集态差异有关。特别的是,-羰基取代的-O-4 型二聚体的薄膜和溶液荧光有显著差别,该模型物由于羰基的荧光淬灭效应在溶液中荧光强度极弱,但其在薄膜状态下则显示出明显的荧光发射,文献作者猜测该现象可能与-O-4连接键在薄膜状态下发生部分裂解有关。其实,不仅是模型物,羰基对木质素的荧光淬灭效应在原本木质素中也得到证实。Castellan 等研究了杉木薄片在硼氢化钠还原前后的荧光行为。结果表明,杉木薄片经硼氢化物还原后,其在320~500nm处的蓝色荧光强度显著增强,结构表征显示荧光强度的增大幅度与杉木薄片中的羰基含量呈负相关。同时,作者认为该现象也说明植物细胞壁荧光来自木质素而非碳水化合物,因为即使碳水化合物可以发射蓝色荧光,也最可能来源于糖单元上的羰基或醛基,但这些富电子基团被还原后,木材的荧光不降反升,说明荧光并非来自碳水化合物。木质素荧光强度除了受羰基化合物等荧光淬灭剂影响外,Davidson 等在研究漂白剂对纸浆残留木质素荧光影响时提出自吸收效应也是影响木质素荧光强度的主要原因之一。作者发现随着双氧水漂白次数的增加,纸浆残留木质素荧光强度不降反升。根据纸浆吸收光谱的变化规律,认为木质素荧光强度的增加主要与420~450nm 处的黄色吸光团被漂白剂破坏有关。

图4 对木质素荧光影响较大的模型化合物[24-26,28-30,34]
由于木质素大分子内包含多种不同结构的荧光团,单一荧光团的荧光光谱往往无法与真实木质素完全匹配。Tylli等采用反卷积分峰方法对纸浆残留木质素的宽荧光光谱进行了分峰处理,并利用26 种共轭程度较强的模型物荧光光谱进行线性加权拟合,通过谱图匹配性和模型物结构的相对含量来识别荧光团。结果显示,除肉桂酸、香豆素、黄酮类和二苯乙烯四种结构的最大发射波长可以与木质素小于450nm的短波长荧光发射相匹配外,其他所有结构的发射波长均太短。然而,发射波长大于450nm的模型物并没有找到,作者猜测木质素长波长发射与其在氧化偶联过程中生成的延伸共轭体系有关。Albinsson 等则进一步研究了顺-反异构和苯并呋喃单元对二苯乙烯类模型物荧光性质的影响。结果表明,二苯乙烯模型物的顺-反异构体荧光性质表现出很大差异,其中反式结构具有较强荧光,最大发光波长在383nm处,而顺式结构在室温下不显示任何荧光。根据二者的单晶结构差异,作者将其荧光差异归结为反式结构的平面性好,而顺式结构的分子内激发态弛豫现象严重。具有苯并呋喃单元的二苯乙烯类模型物由于有化学键连接,其分子平面性进一步增强,从而表现出更强的荧光发射。最后,根据磨木木质素和上述模型物的荧光光谱形状对比结果,作者得出含有苯并呋喃单元的二苯乙烯类模型物荧光与真实木质素的匹配性更高。
Gardrat 等报道了不同酚羟基醚化基团对5-5键模型物荧光性质的影响。结果表明,未取代的联苯二酚型模型物的荧光量子产率最高,最大发射波长最长,分别达到0.33 和380nm。当酚羟基被甲基、苯甲基等基团醚化后,模型物荧光量子效率显著降低,最大发射波长也向短波长移动。作者认为发射光谱的蓝移主要由酚羟基和烷氧基的助色作用强弱不同导致,不过荧光量子效率的显著差别则很难解释。作者继续合成了八元环醚化结构的5-5键模型物,尽管最大发射波长仍然蓝移,但分子的荧光量子效率只是略有下降。与甲基醚化后的结构相比,联苯二酚型模型物两个子结构之间存在氢键作用,相对运动受限,八元环醚化结构则通过化学键限制了两个子结构的自由运动。因此,作者提出模型物的荧光量子产率大小与其内部能量转换过程的不同振动模式有关。根据近年来发展起来的聚集诱导发光理论,认为上述现象主要与5-5 键的分子旋转受限有关。之后,Machado 等则采用密度泛函理论计算进一步说明了5-5键模型物的分子旋转对其荧光量子产率的影响机制。5-5键模型物的势能曲线显示苯环的相对位置变化会影响分子激发态和三重态之间的带隙大小,部分构象甚至会出现单重态和三重态能量的交叉重叠,此时系间窜越成为5-5 键模型物分子激发态能量耗散的主要原因,见图5。当5-5 键的自由旋转受限后,可以有效抑制系间窜越从而提高荧光量子产率。2013 年,Lähdetie 等则对木质素的长波长荧光来源提供了一种解释方案。作者研究了以-O-4和5-5键为主要连接键的二聚体、三聚体和四聚体模型物在532nm 超长波长激光激发下的荧光性质。结果显示,与非共轭的-O-4键相比,拥有共轭5-5键木质素模型物在532nm波长的激发下仍然有较明显的荧光发射,而且其荧光强度与5-5键的可自由旋转程度呈正相关,即自由旋转程度越高,荧光强度越强。按照常规理论,文中5-5键模型物的分子共轭程度不足以发出大于532nm的长波长荧光,作者猜测模型物的5-5键可以为离域电子共轭提供更多路径,而且分子构象也是影响荧光的关键因素。尽管作者检测到5-5 键模型物存在超长波长荧光发射,但其强度仍然很弱,需采用激光作为激发光源,而木质素在普通光源的激发下就有长波长荧光发射。因此,木质素长波长荧光来源仍然有待探索。上述研究均以小分子模型物的荧光性质为参照,但小分子模型物毕竟与木质素的结构差别大,其结论存在一定局限性。

图5 5-5键模型物的S0、S1、T1结构模型和部分电子能级的势能曲线[33]
2006 年,Radotić 等首次以松柏醇脱氢寡聚物代替小分子模型物为研究对象,采用反卷积技术分析其荧光光谱和寿命来推测发光基团。首先,当激发波长大于360nm后,该寡聚物也具有可见光荧光发射,其最大发射波长约在440nm,而且随着激发波长的增加,荧光发射波长向长波方向移动,说明寡聚物也属于多荧光团发光体系。其次,反卷积结果显示,寡聚物根据发光波长划分的荧光团最少包含四种,其波长位置分别位于425~430nm、483~491nm、501~507nm 和535~540nm;根据荧光寿命划分,寡聚物也含有四个动力学组分,其发光寿命分别为0.07ns、0.50ns、1.9ns和7.7ns,且荧光寿命越短,组分的发射波长能量越高,所占权重越大。最后,作者认为虽然寡聚物的化学结构较真实木质素简单,但其长波长荧光团结构依然无法判断,猜测其不同荧光团可能是分子结构差异,也可能是分子所处化学环境的差异。2007 年,Djikanović 等继续采用对数正态分布模型对松柏醇、阿魏酸和松柏醇寡聚物三种木质素模型物的荧光光谱进行反卷积处理,分析其荧光团数量。尽管松柏醇和阿魏酸均是单苯环小分子模型物,但拟合结果显示松柏醇只含有一个荧光基团,而阿魏酸却含有两个荧光基团,概率密度计算显示其最大发射波长分别位于390nm和420nm。作者猜测阿魏酸会因氢键作用在有机溶剂中形成二聚体,从而显示出两个荧光团的发射光谱。松柏醇寡聚物的结果则更为复杂,其反卷积结果显示荧光光谱来自多个不同的荧光基团,分别位于397nm、420nm、440nm、460nm、480nm和505nm,作者认为这主要与松柏醇在脱氢耦合过程中形成的多级共轭体系有关,但具体结构不清晰。同时,Djikanović 等合成的松柏醇脱氢寡聚物的荧光性质与Radotic 等的结果存在显著差异,说明即使木质素合成前体结构完全相同,合成条件相似,但由于聚合位点的随机性和无序性,寡聚物的荧光性质也不尽相同,这无疑进一步增加了木质素荧光的研究难度。Djikanović 等研究了松柏醇寡聚物以及分别掺杂了10%(质量分数)阿魏酸和对香豆酸的两种二元寡聚物的荧光性质。结果显示,在360nm波长激发下,三种寡聚物均可以发出可见光区域的荧光。与纯松柏醇寡聚物相比,掺杂了阿魏酸的荧光光谱发生了蓝移,掺杂了对香豆酸的荧光光谱发生了红移。根据聚合物结构分析,这主要与掺杂单体的活性聚合位点有关,对香豆酸酚羟基邻位没有被甲氧基取代,脱氢耦合过程中生成了共轭程度更高的荧光团,所以荧光光谱表现为红移,阿魏酸则恰恰相反。然而,当阿魏酸的掺杂量提高到20%(质量分数)时,寡聚物荧光光谱发生较大变化,在535nm左右出现一个较强的荧光发射峰,结构分析显示寡聚物中形成了更长的C=C 键共轭结构域,可以形成更大的共轭大分子,所以荧光光谱红移。和小分子模型物相比,寡聚物的荧光性质与真实木质素相似,属于多荧光团体系,且聚合后产生了长波长荧光团。然而,虽然寡聚物的聚合单体结构和组分含量可控,但单元之间的连接方式多样无序,亦无法精准判断不同波长荧光团的化学结构。
除模型法之外,以植物细胞壁为对象,通过分析原本木质素在定向处理前后的荧光变化探究木质素荧光团结构也是研究策略之一。Harris 等采用紫外荧光显微镜对比了草本植物细胞壁在氨气碱化处理前后的荧光行为。结果显示,细胞壁在氨气处理后荧光强度增强,而且未完全木质化的细胞壁荧光颜色还会由蓝色变为绿色。根据草本植物细胞壁的结构特点,这主要是由于未完全木质化的细胞壁中存在单酯键连接的阿魏酸和对香豆酸,碱性条件下苯酚结构电离为酚氧负离子,电子云密度增大,从而使荧光发射从蓝色红移到绿色,说明草本植物细胞壁中木质素荧光来源可能与阿魏酸和香豆素酯有关。Morales 等以可能发射出蓝绿色荧光的四种甜菜叶内源性化合物,阿魏酸、对香豆酸、山柰酚和槲皮素为对象,从稳态、瞬态和温敏荧光等方面探究了甜菜叶片蓝绿色荧光的来源。结果表明,山柰酚和槲皮素的荧光强度很弱,可以首先排除。与对香豆酸相比,阿魏酸的荧光强度相对最高,且最大荧光发射波长可以达到420nm,与甜菜叶片的蓝绿色荧光最匹配。同时,阿魏酸的荧光寿命与甜菜叶片蓝绿色荧光的主要动力学荧光团(70%)一致,并且二者的温敏荧光变化也相似。因此,作者认为阿魏酸是甜菜叶片蓝绿色荧光的主要来源。在此基础上,Lichtenthaler 等通过对比不同类别、不同生长环境和碱水解处理前后的植物叶片蓝绿色荧光强度变化,并与其阿魏酸含量相关联,探究植物叶片蓝绿色荧光来源。结果显示,单子叶禾木植物叶片(小麦、玉米等)的阿魏酸含量远高于双子叶禾木植物叶片(狐尾草、马齿苋等),与此同时,二者的蓝绿色荧光强度表现出相同的大小规律。南瓜、向日葵等植物甚至不含阿魏酸,只含有少量咖啡酸,其蓝绿色荧光强度更低。采用碱水解可以脱除这些植物细胞壁上的阿魏酸,对于本身阿魏酸含量极低的植物,其蓝绿色荧光大小几乎不变,而本身阿魏酸含量较高的植物,其蓝绿色荧光强度显著下降。作者根据上述结果,进一步验证了细胞壁上化学键连接的阿魏酸是植物叶片蓝绿色荧光的主要来源。
研究人员对木质素荧光团化学结构的筛选开展了大量工作,但迄今为止依然没有定论,分析认为主要由下面三方面原因造成。第一,不同来源木质素的荧光差异导致筛选出的荧光团结构不同。木质素是由多种苯丙烷前体无序聚合而成的芳香族大分子,具有天然的异质性和化学结构的复杂性,其各组分含量和子结构连接方式均存在差异。因此,不同来源/批次木质素可能筛选出不同结构的模型物。第二,木质素自身荧光性质研究亟待加强。研究人员往往从荧光强度和发射波长两方面筛选与真实木质素荧光相近的模型物作为其荧光团,此时木质素自身荧光性质测量的“准确性”至关重要:一方面,木质素结构中存在羰基等荧光淬灭剂,对木质素真实荧光强度的判断造成干扰;另一方面,木质素发光波长与其紫外吸收光谱部分重叠,自吸收效应不仅会影响木质素荧光强度,甚至会改变其光谱形状。第三,将木质素看作单一荧光团或相互作用可忽略的多荧光团系统,存在一定局限性。木质素多种荧光团随机分布在三维网络结构中,并非单独存在。荧光团间可能存在的电荷及能量转移会使电子能级结构发生重排,从而影响木质素整体的荧光光谱。上述文献表明,甚至是二苯乙烯类、阿魏酸、5-5 键等小分子模型物的荧光性质也与其聚集状态密切相关,木质素敏感的微结构变化可能进一步放大荧光团间相互作用的影响。
2.2 木质素荧光团间聚集耦合态
前面介绍的研究工作多从单分子荧光团或荧光团间的简单线性加和来理解木质素发光机制,但木质素子结构的差别不仅体现在化学结构上,还体现在相对方向、间隔距离等微结构的化学构象上,部分荧光团甚至在其微结构的空间限域作用下发生电子能级的耦合与重排。
其实早在1978 年,Lundquist 等就报道了真实木质素荧光团间的耦合现象。作者在研究云杉磨木木质素的基本荧光性质时发现,其荧光发射光谱和激发光谱的形状几乎不随波长(小于400nm)的改变而变化,说明虽然木质素属于多荧光团体系,但其荧光行为却与只含有一个发色团的分子相似,见图6(a)。推测木质素的荧光团间存在相互作用,形成了无辐射能量转移系统,短波长荧光团部分能量通过该系统转移到末端的长波长荧光团,从而表现出荧光能量的一体性。为了验证木质素荧光团间的能量转移行为,2003 年,Barsberg 等以典型的电子受体对苯醌为掺杂剂,研究了木质素在掺杂前后的荧光变化,见图6(b)。结果显示,对苯醌使木质素荧光急剧淬灭,结合二者的吸收光谱性质及其混合物的吸收光谱变化规律,作者认为该淬灭现象不是由对苯醌的自吸收效应导致的,而是木质素荧光团将其激发态能量转移到对苯醌,由于对苯醌不发光而导致荧光淬灭。其次,由于对苯醌的吸收光谱仅与木质素荧光光谱部分重叠,但木质素荧光却表现出整体下降趋势,即木质素多荧光团荧光能量对外表现出能量的一体性,这间接证明木质素自身荧光团间也存在激发态能量转移。最后,作者提出木质素的荧光团并非某个确定的化学结构,而是子结构通过分子间相互作用力诱导产生激发态电荷转移后形成的准分子或激态复合物,换句话说,木质素的真实发光基团往往是其荧光团间的聚集耦合态。2015 年,Djikanović 等意外通过木质素寡聚物的热力学性能研究验证了木质素荧光团间的耦合行为。在对比不同比例松柏醇、阿魏酸和对羟基香豆酸三种单体脱氢耦合制备的木质素寡聚物的荧光变化时发现,不同寡聚物在碱性水溶液中具有形状相似的可见光荧光发射,但荧光性质与聚合单体组成比例关联性较小,特征不明显。有趣的是,在与高分子热力学性能的关联对比中,发现不同寡聚物的最大荧光发射波长与其玻璃化转变温度表现出良好的关联性,玻璃化转变温度越高,寡聚物的最大荧光发射波长越小。作者认为上述结果说明寡聚物荧光发射波长与其分子胶团的紧密度有关,玻璃化转变温度越高,胶团的紧密度越高,分子内聚集越严重,导致荧光发射波长红移。该研究从寡聚物角度间接验证了木质素荧光团间存在相互作用。2019年,Qiu等在木质素-罗丹明构建的荧光能量转移系统中再次验证了木质素荧光团间的能量转移行为。在荧光能量转移系统中[见图6(c)~(e)],供体荧光团的荧光寿命会因为发生非辐射能量转移而明显减小。结果显示,虽然木质素的荧光光谱强度因发生能量转移而整体降低,但只有与罗丹明吸收光谱重叠区域的木质素荧光团的荧光寿命明显缩短,非重叠区域的荧光团寿命几乎没有变化,说明只有光谱重叠区域的木质素荧光团与罗丹明发生了非辐射能量转移。作者认为非重叠区域的荧光团是通过木质素胶团内固有的能量转移机制,将荧光能量间接地转移到罗丹明,从而保持木质素荧光能量的一体性。

图6 不同条件下的荧光光谱、平均荧光寿命及能量转移途径示意图[23,29,42]
众所周知,荧光团间的距离会影响其相互作用的强弱。木质素在提取过程中会产生羟基、羧基甚至磺酸基等亲水基团,这使木质素大分子聚集行为对环境变化异常敏感,拥有大分子弱聚电解质特有的多尺度聚集行为。该复杂聚集行为为木质素荧光团间聚集耦合态的研究提供了更大的空间。2010年,Yan等报道了木质素磺酸盐在水溶液中的浓度聚集荧光行为。结果显示,当木质素磺酸盐浓度低到0.001g/L时,其激发光谱形状与紫外吸收光谱基本一致,最大激发波长在280nm处。随着浓度的增加,280nm处的激发强度越来越弱,最大荧光激发波长向长波长移动,浓度大于0.2g/L后移动到350nm处,浓度大于0.5g/L后在400nm附近出现一个较强的肩峰,见图7(a)。根据木质素磺酸盐微结构的浓度聚集行为,作者认为浓度诱导的木质素聚集行为会使芳香环之间发生或增强耦合作用,从而导致激发光谱的红移。同年,Deng等报道了碱木质素在四氢呋喃中的浓度聚集荧光行为。结果显示,在一定浓度范围内,虽然木质素荧光发射光谱的形状和最大发射波长随浓度增加不发生变化,但对应的荧光激发光谱则发生明显红移,见图7(b)。根据分子激发态耦合理论,可以认为上述光谱红移现象说明碱木质素苯丙烷单元间的聚集行为属于J-聚集。之后,Deng等继续采用该方案研究了不同来源木质素的浓度聚集荧光行为,发现了相似的激发光谱红移现象,提出木质素苯丙烷单元间的J-聚集属于木质素的固有性质。除上述聚集荧光淬灭机理外,近年来,聚集诱导发光和聚集空间共轭等荧光基础理论发展迅速,为解释木质素复杂的荧光行为提供了新思路。2016 年,Qiu 等基于木质素在混合溶剂中的自组装行为,首次报道了木质素的分子内聚集诱导荧光增强现象。混合溶剂中良溶剂对木质素有很好的溶解性,非良溶剂则很难溶解木质素,通过增加非良溶剂的比例可以精确控制木质素的聚集状态。作者先以磺化碱木质素为对象,研究了其在不同比例水/乙醇体系中的聚集荧光行为[见图7(c)],结果显示,随着不良溶剂乙醇含量的增加,木质素发生聚集,其荧光发射强度随之增大,当乙醇体积分数达到90%时,荧光强度增大了15 倍左右。为了排除溶剂极性变化对木质素荧光的影响,作者又研究了磺化碱木质素在十六烷基三甲基溴化铵的静电相互作用诱导下的聚集荧光行为[见图7(d)],结果显示,随着十六烷基三甲基溴化铵含量的增加,木质素荧光依然表现出典型的聚集荧光增强效应。然而,当实验对象换为碱木质素时,其在混合溶剂中却表现出聚集荧光淬灭现象。通过对碱木质素和磺化碱木质素聚集行为的深入分析对比,发现虽然从宏观尺度看碱木质素可以很好地溶解在有机溶剂中,但由于缺乏静电斥力,碱木质素胶团内部聚集严重,可以说碱木质素在有机溶剂中的初始聚集状态与磺化碱木质素在90%(体积分数)乙醇中的相似。因此,随着非良溶剂水的加入,刚性木质素荧光胶团间的聚集荧光行为与传统的聚集荧光淬灭效应一致。随后,作者将碱木质素的良溶剂换为碱溶液,引入静电斥力,此时的碱木质素也表现出聚集荧光增强效应。结合木质素的化学结构和微结构特点,作者推断木质素胶团内聚集导致的荧光团的分子内运动受限和羰基等富电子基团的聚集空间共轭效应是其聚集荧光增强效应的主要原因。

图7 不同条件下木质素的荧光激发光谱和荧光光谱[43-44,46]
除上述两种木质素外,2018年,Li等在酶解木质素体系中报道了聚集荧光增强效应。在研究酶解木质素在乙醇中的浓度聚集荧光行为时发现,当浓度达到0.02mg/mL时,酶解木质素的荧光强度由原先的线性增加转为跳跃式增强,见图8(a)。根据紫外吸收光谱的相应变化,作者猜测酶解木质素的浓度聚集荧光增强效应主要与木质素苯丙烷单元的J-聚集有关。作者继续研究了酶解木质素在碱溶液/乙醇(良/非良)混合溶剂中的聚集荧光行为[见图8(b)],结果显示酶解木质素表现出典型的聚集荧光增强效应。认为碱溶液会使酶解木质素的酚羟基等官能团发生电离,破坏木质素分子内原本的J-聚集,加入非良溶剂乙醇后,J-聚集效应再次增强,木质素荧光强度与J-聚集的相关性再次说明木质素苯丙烷单元间的J-聚集行为是诱导木质素聚集荧光增强的原因。2020 年,Qiu 等基于传统的共轭发光和聚集诱导发光理论,分别从木质素胶团间/胶团内等不同聚集维度和溶剂极性、pH、离子强度、阳离子添加剂等不同溶剂环境角度,对木质素的聚集荧光行为和机理进行了较为系统的研究。结果表明,浓度诱导的木质素胶团间聚集不会形成激态缔合物或因碰撞发生激发态能量耗散,对木质素荧光不产生实质影响,该现象被认为与松散木质素胶团所形成的空间位阻效应相关。常见的木质素浓度聚集荧光淬灭和荧光发射/激发光谱红移现象主要由自吸收效应导致[见图8(c)],而且吸收光谱中波长的相对吸光度越大,对应的荧光淬灭浓度拐点越低,从而造成木质素最大荧光激发波长随浓度增大向长波长移动的现象,见图8(d)。因此,前期研究结果中根据浓度诱导木质素荧光光谱红移得出的J-聚集结果需要进一步验证。松散木质素胶团内聚集则会诱导荧光增强,与前期文献结果一致,然而当胶团紧密度超过一定临界值后,木质素会由聚集荧光增强型荧光团转变为聚集荧光淬灭型荧光团,该现象可能与木质素松散胶团压缩导致的空间位阻效应减弱有关。上述胶团间和胶团内聚集荧光规律与木质素在不同pH 和离子强度等溶剂环境显示的复杂聚集荧光行为一致。

图8 木质素在不同条件下的光谱图[11,47]
即使与纤维素、半纤维素等通过共价键和分子间作用力固定在植物细胞壁中,原本木质素的荧光对其聚集状态变化也具有响应性。2010 年,Donaldson 等采用共聚焦荧光显微镜研究了原本木质素随木材压缩程度变化的荧光行为,见图9(a)、(b)。结果显示,随着压缩程度的增强,原本木质素在紫外线激发下的荧光强度增大,且最大发射波长红移,从紫色移动到蓝色,说明原本木质素中的苯基丙烷单元会在挤压下聚集形成更大的共轭体系。随后,Donaldson 等采用时间相关单光子计数的瞬态荧光技术继续研究了木材压缩程度对原本木质素荧光寿命的影响。结果显示,与正常木材相比,压缩木材中的原本木质素的平均荧光寿命显著降低,从0.473ns下降到0.418ns。作者将二者的荧光寿命区别归结于化学结构和分子环境两方面原因。化学结构方面,压缩木材中的原本木质素共轭程度大,拥有更长的C=C/C―C键交替体系,这些平行取向的共轭荧光团随着聚集程度的增强容易导致荧光寿命降低;分子环境方面,由于正常木材和压缩木材生长环境的差异,细胞壁中与原本木质素结合的非荧光多糖结构不同,从而导致二者荧光寿命产生差别。2017年,Auxenfans等研究了蒸汽爆破预处理对木质素荧光的影响[见图9(c)、(d)],作者发现蒸汽爆破预处理会导致原本木质素荧光强度明显降低,然而组分含量分析显示荧光损失并非源于木质素含量的降低。作者认为高强度的预处理过程会导致木质素-芳基醚键含量降低,空间聚集结构遭到破坏而导致荧光降低,即木质素聚集程度与其荧光强度呈正相关。不过,由于细胞壁结构的限制,难以系统性地揭示木质素微结构聚集行为对其荧光的影响规律。

图9 不同样品的共聚焦荧光图像和三维荧光图[48,50]
在木质素微结构聚集行为研究和聚集诱导发光理论的基础上,木质素荧光团间聚集耦合态的研究近年来取得了一些进展,从宏观层面基本明确了木质素荧光团间的相互作用以及胶团间和胶团内两种木质素基本聚集行为对其荧光的影响规律和作用机制。因此,单独分析木质素荧光团的化学结构往往意义有限,木质素真实发光基团是这些荧光团间的聚集耦合态。在此基础上,认为还有以下两方面问题需要思考:第一,木质素存在多尺度复杂聚集行为,木质素聚集行为对荧光的影响规律需从不同角度和维度进一步完善,构建木质素物化性质-溶液行为-荧光变化的关联模型;第二,木质素聚集荧光行为仅从宏观层面反映了聚集行为变化对荧光团间耦合作用的影响规律,需进一步从微观分子层面揭示木质素荧光团间的耦合机制,才可能取得更多突破。
3 结语
本文从木质素荧光团化学结构筛选和木质素荧光团间聚集耦合态研究两方面总结并讨论了木质素荧光机理的研究现状,并对其中的一些问题进行了梳理凝练。在木质素荧光团化学结构筛选方面,相关研究主要通过对比模型物荧光性质或分析木质素定向处理前后的荧光变化规律来确定木质素荧光团结构,不同来源木质素往往筛选出的荧光团结构存在差异。这一现象尽管与木质素的异质性和复杂化学结构有关,但更主要是对木质素自身荧光性质研究不透彻,忽略了木质素荧光团间相互作用的影响。在木质素荧光团间聚集耦合态研究方面,相关研究主要通过调控木质素聚集状态或添加荧光受体来分析木质素荧光机理。目前,虽然对木质素基本聚集荧光行为的作用规律和影响机制有了一些认识,但仍然需结合木质素的多尺度复杂聚集行为,从不同角度和维度完善二者的关联模型,并进一步从分子层面揭示木质素荧光团间的耦合机制。
总的来说,木质素荧光的复杂性不光在于其化学结构,还应考虑木质素微结构聚集行为引起的激发态能量转移。由于无法直接监测电子的跃迁过程,目前只能基于理论和经验对木质素荧光机理进行推导和判断。尽管研究人员已经做了一些工作,但木质素荧光的研究仍然处于起步阶段,有待进一步拓展和深入,期待未来木质素荧光机理研究随着观测技术和研究理论的进步取得较大发展。除了荧光机理复杂外,木质素荧光量子效率低、应用领域范围较窄也制约着这种天然荧光材料的应用前景。可喜的是,近年来木质素碳点研究发展迅速,可能是实现木质素荧光特性功能化的突破口。
