白三烯受体拮抗剂治疗支气管哮喘的药物基因组学研究进展▲
2022-05-25杨一民洪菲萍石慧莉王珊珊黄永莲
杨一民 洪菲萍 石慧莉 王珊珊 王 颖 黄永莲 李 溦
(1 福建省厦门市中医院儿科,厦门市 361009,电子邮箱:yym216@126.com;2 福建省厦门弘爱医院新生儿科,厦门市 361000;3 福建中医药大学,厦门市 350108)
【提要】 支气管哮喘患者可能存在基因位点单核苷酸多态性,从而影响白三烯受体拮抗剂(LTRA)治疗效果,包括影响白三烯作用通路的基因(5-脂氧合酶基因、白三烯A4水解酶基因、白三烯C4合成酶基因、半胱氨酰白三烯基因)、影响LTRA的药代动力学的基因和其他基因的多态性等,这些研究结果推动了支气管哮喘精准治疗的开展。本文对近年来LTRA治疗支气管哮喘的药物基因组学的研究进展进行综述,以期为支气管哮喘个体化治疗及相关临床研究提供参考。
支气管哮喘(简称哮喘)是由环境和遗传等因素共同作用引起的一种疾病。白三烯是哮喘发作过程中的重要炎症介质。2014年修订的全球哮喘防治创议(Global Initiative for Asthma,GINA)指南[1]明确指出,白三烯受体拮抗剂(leukotriene receptor antagonist,LTRA)是除糖皮质激素外,唯一可单独长期应用的哮喘控制药物。抗白三烯治疗的作用机制主要有抑制5-脂氧合酶与半胱氨酰白三烯1型受体结合、竞争性抑制白三烯与受体结合两种途径,前者代表药物为齐留通,后者代表药物为孟鲁司特钠。2020年版GINA指南仍把LTRA作为哮喘治疗的重要控制药物之一[2],然而LTRA治疗哮喘患者的效果个体化差异较大,有研究显示,35%~78%的哮喘患者接受LTRA治疗无效[3],这种疗效差异性被认为是由于患者部分基因遗传变异所导致的[4],即不同个体对相同药物的活化、代谢、清除能力不同,这种现象被称为药物基因组学[5]。目前关于LTRA的药物基因组学研究主要侧重于LTRA代谢途径所涉及的基因变异。本文就近年来关于LTRA治疗哮喘的相关药物基因组学的研究进展作一综述,以期为哮喘患者拟定个体化的治疗方案提供参考。
1 影响白三烯作用通路的基因
1.1 5-脂氧合酶基因 当嗜酸性粒细胞受到刺激可释放花生四烯酸,在细胞内与5-脂氧合酶代谢生成5羟过氧化二十碳四烯酸(5-hydroperoxyeicosatetraenoic acid,5-HPETE),同时5-脂氧合酶也能催化5-HPETE生成白三烯A4。5-脂氧合酶为白三烯合成途径上的限速酶,由5-脂氧合酶基因编码合成,5-脂氧合酶基因位于人体10号染色体(10q11,2)上,包含14个外显子,其启动子区的SP1结合部位(-GGGCGG-)与转录子的合成有关,可影响启动子转录活性。5-脂氧合酶基因最常见的等位基因型特点为在5-脂氧合酶核心启动子位点含有5个SP1(-GGGCGG-)串联性重复,其被视为野生型等位基因,其余还包括变异型等位基因,纯合型等位基因、杂合型等位基因、纯合型等位基因、杂合型等位基因[6]。5-脂氧合酶基因启动子区域的变异会导致半胱氨酰白三烯(cysteinyl leukotrienes,CysLTs)合成减少,进而影响白三烯的合成;同时,使用LTRA能够减少机体白三烯的合成,进而抑制5-脂氧合酶基因的表达[7]。其中孟鲁司特主要与支气管平滑肌中的CysLTs受体选择性结合,竞争性阻断CysLTs,进而阻断细胞对CysLTs的反应,齐留通则通过抑制花生四烯酸与5-脂氧合酶在细胞内的代谢过程进而抑制CysLTs的合成,具体机制见图1。
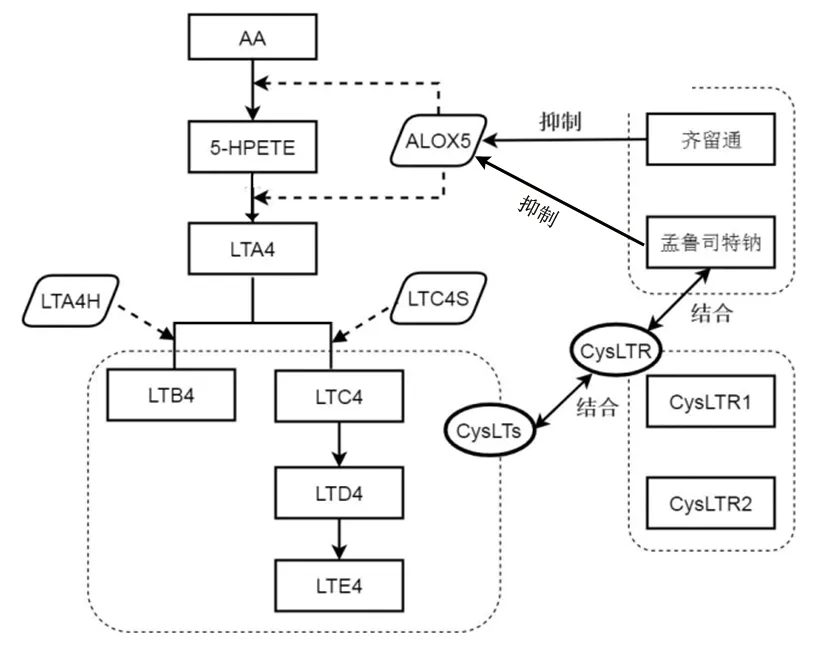
图1 白三烯代谢途径和白三烯拮抗作用的作用机制途径
注:AA为花生四烯酸;5-HPETE为5羟过氧化二十碳四烯酸;ALOX5为5-脂氧合酶;LTA4、LTB4、LTC4、LTD4、LTE4为白三烯A4、白三烯B4、白三烯C4、白三烯D4、白三烯E4;LTA4H为白三烯A4水解酶;白三烯C4S为LTC4合成酶;CysLTs为半胱氨酰白三烯;CysLTR为半胱氨酰白三烯受体。
研究发现,5-脂氧合酶启动子中的串联碱基重复变异与基因活性表达降低有关[8]。刘占利等[9]研究发现,哮喘患儿全血中5-脂氧合酶mRNA表达增加,他们认为5-脂氧合酶基因启动子SP-1结合位点能够调控5-脂氧合酶mRNA的表达。已有研究证实人体5-脂氧合酶基因位点rs2115819(G>A)与孟鲁司特治疗支气管哮喘的疗效有关,与A等位基因的携带者(AA、AG)相比,GG基因型哮喘患者在接受LTRA治疗后的1 s用力呼气容积(forced expiratory volume in one second,FEV1)改善更多,孟鲁司特治疗GG基因型的哮喘患者临床疗效更佳,复发率更低,研究者认为孟鲁司特对5-脂氧合酶基因位点G等位基因的表达影响更大[10]。也有研究表明,LTRA会降低5-脂氧合酶基因的表达,同时减少花生四烯酸的释放,进而影响白三烯的合成过程[11]。
周奇兴等[6]研究发现,携带5-脂氧合酶基因启动子野生型等位基因的哮喘患者对LTRA治疗有效,而携带变异型等位基因的患者则无效。一项针对270例哮喘控制不良儿童的研究发现,5-脂氧合酶基因启动子区基因序列为纯合型的患儿,其体内白三烯合成增加,尿液LTE4含量增多,这部分患儿使用LTRA的疗效较差;进一步研究发现,这部分患儿86%是非裔美国人,提示非洲祖先基因的单核苷酸多态性可能影响了哮喘儿童对LTRA的药物反应[12]。也有学者提出不同的观点,他们认为5-脂氧合酶基因启动子的多态性导致了孟鲁司特疗效的差异,与野生型纯合子相比,变异型等位基因携带者在使用孟鲁司特治疗后恶化的风险降低了73%,这表明变异等位基因上调了5-脂氧合酶活性[13]。Drazen等[14]认为使用ALOX5抑制剂ABT-761可改善野生型纯合子与杂合子哮喘患者的FEV1。Telleria等[15]研究发现,5-脂氧合酶基因中CC型和CT型哮喘患者在接受孟鲁司特治疗后FEV1明显改善,而TT型患者无明显改变。由此可以推测,LTRA对具有变异型纯合子5-脂氧合酶基因的哮喘患者疗效较差,而对具有野生型5-脂氧合酶基因的哮喘患者疗效较好。此外,一项回顾性研究发现,14.8%哮喘控制不良儿童的5-脂氧合酶启动子区rs59439148位点存在变异,此类患儿的尿LTE4水平升高,FEV1降低,哮喘恶化几率更高,这可能与LTE4水平升高,增加CysLTs的暴露有关[16]。
1.2 LTA4H基因 由LTA4H基因编码的LTA4H水解酶能催化LTA4转化为LTB4,LTB4是中性粒细胞催化因子之一,可激活中性粒细胞参与哮喘的发生与发展。LTA4H基因位点rs2660845(G>A)的变异型与哮喘患者的最大呼气峰流速(peak expiratory flow,PEF)、FEV1有显著关联,与G等位基因相比,携带A等位基因的患者在使用LTRA后PEF、FEV1显著提高[17]。有研究发现,LTA4H基因位点携带G等位基因的哮喘患儿哮喘加重率是AA型基因的4~5倍,研究者认为G等位基因可调控白三烯的合成,加重了炎症反应[18]。Maroteau等[19]研究表明,LTA4H的遗传变异以及哮喘发作时间的不同可能会导致孟鲁司特钠治疗效果的差异,LT4H基因位点中携带至少一个G等位基因的患儿病情恶化风险更高,而AA型基因患儿血液中LTB4浓度较前者低,研究者认为A等位基因可降低LT4H活性,影响孟鲁司特作用于CysLTs受体,通过LTB4途径影响白三烯的合成,进而引起哮喘病情恶化的风险降低。还有研究发现,在哮喘治疗期间LTA4H rs2660845带G等位基因的哮喘患者哮喘恶化的风险是纯合子的4~5倍,可能与G等位基因下调LTA4H的活性有关,导致LTA4偏离LTA4H途径,从而增加CysLTs的形成[13]。
1.3 LTC4S的基因 由LTC4S基因编码的LTC4合成酶参与了白三烯合成过程,其能催化LTA4转化成LTC4,LTC4能脱酸形成LTD4,LTD4可再进一步形成LTE4。LTB4、LTC4、LTD4和LTE4都含有半胱氨酸残基,故之称为CysLTs。
LTC4S基因位于5号染色体长臂的末端(5q35),其启动子区A-444位存在C替代A的情形,是LTC4S基因多态性的一种。王婷婷等[20]发现,在新疆维吾尔族支气管哮喘患儿中,LTC4S基因A-444C位点三种基因型CC、AC和AA的分布频率分别为6.8%、49.0%和44.2%,A、C等位基因频率分别为68.7%、31.3%,与韩国、澳大利亚及中国东北地区汉族支气管哮喘儿童存在显著差异。接受LTRA治疗后,与LTC4S基因A-444C位点AA纯合子相比,杂合子发生哮喘恶化的风险降低76%[13]。
有研究认为LTC4S基因rs730012位点携带C变异等位基因的哮喘患者对LTRA的反应优于AA型患者[21]。袁姝华等[22]在临床上使用吸入糖皮质激素联合LTRA治疗中度持续哮喘患儿的研究中发现,LTC4S基因rs730012位点上AC型与CC型患儿的50%肺活量时FEF改善率和75%肺活量时FEF改善率均优于AA型,且75%肺活量时FEF的改善更明显。蔡畅等[23]研究发现,LTC4S基因AC/CC型的哮喘患者接受LTRA治疗后,FEV1的提高及晨尿LTE4的下降有明显变化,而LTC4S基因AA型患者则无改变,这说明LTRA对携带C等位基因的哮喘患儿的疗效更佳。但也有研究得出相反结论[24]。LTC4S基因rs730012位点多态性还与哮喘严重程度有关,C等位基因频率增加提示哮喘病情严重或发展为重度哮喘的可能性大[25]。还有研究提示LTC4S基因位点A>C的变异与呼吸道合胞病毒(respiratory syncytial virus,RSV)毛细支气管炎的发生及病情严重程度有关[26],董明瑛等[27]则认为其二者无相关性,但RSV毛细支气管炎患儿LTC4S位点多态性可能与LTRA疗效有关,且AC/CC基因型患儿疗效优于AA型。
有研究表明,前列腺素D2能够募集和刺激嗜酸性粒细胞并诱导LTC4的合成,LTC4S-444C和前列腺素D2受体-441C等位基因的组合效应可能对增加总嗜酸性粒细胞计数具有累加效应,导致较高水平的LTC4,破坏哮喘儿童对LTRA反应性[28]。 潘明鸣等[24]研究北京哮喘患者的LTC4S基因位点,提示基因型为AA/AC/CC的携带者分别占为65.4%、30.5%、3.8%,这与吴永红[29]研究的西南地区的成人哮喘患者LTC4S基因型分布频率结果相似。而在北京正常成年人群中LTC4S基因变异型为C-444的出现频率为19%,与日本相近,低于美国和澳大利亚[24]。但2017年关于我国东北地区哮喘患儿的一项研究发现,LTC4S基因rs730012基因型、等位基因分布与健康对照组无明显差异,即该基因多态性与哮喘的发病率无关[25],这与2011年关于韩国哮喘儿童的一项研究结果一致[28]。这些研究结果的差异可能与人种、遗传背景、环境差异等有关。
1.4 CysLTs基因 CysLTs通过参与气道重构导致哮喘的发生。LTRA通过与支气管平滑肌等部位的CysLTs受体结合,竞争性阻断CysLTs的作用,有效控制哮喘症状,改善患者肺功能。CysLTs受体主要由半胱氨酰白三烯受体(cysteinyl leukotriene receptor,CYSLTR)1和CYSLTR2基因编码,这两个基因都会在肺巨噬细胞和气管平滑肌中大量表达[30]。哮喘患者CysLTR1 mRNA高表达可能与哮喘急性发作存在相关性[31]。在CYSLTR1的基因位点rs773347588上,变异型(CT或TT)比野生纯合型(CC)的患者CysLTs表达量更多,治疗效果更好;而在CYSLTR2的基因位点rs912277和rs912278的变异使得一部分患者在使用LTRA治疗时CysLTs浓度增加,表明CYSLTR2多态性导致部分患者CysLTs的浓度增加,从而哮喘临床表现明显,抗白三烯治疗反应更好[32]。通过实验检测发现哮喘患者CysLTs受体基因mRNA水平与尿LTE4水平、PEF水平有相关性,经过LTRA治疗,尿LTE4水平下降,PEF水平升高[33]。
2 影响LTRA药代动力学的基因
生物个体内的白三烯水平受肠内载体介导转运功能的影响,这种转运受组织离子通道载体家族2B1基因(SLCO2B1基因)编码,其变异位点rs12422149能够影响细胞液中白三烯的水平,有研究证实此变异位点与孟鲁司特治疗后的哮喘症状控制率有关[16]。Mougey等[17]采用体外人结肠腺癌细胞培养模型进行孟鲁司特体外渗透性研究,发现SLCO2B1基因编码的有机阴离子转运蛋白可能是决定孟鲁司特药代动力学和药效学的一个重要因素,同时发现SLCO2B1(G>A)rs12422149变异中,杂合基因型哮喘患者服药后孟鲁司特的血浆浓度明显高于纯合型患者,AG基因型患者对孟鲁司特的反应高于GG基因型。这提示基因多态性可通过影响药代动力学影响LTRA治疗效果。
3 其他位点的基因多态性研究
Dahlin等[34]研究了欧洲、非洲及亚洲哮喘患者使用孟鲁司特治疗的效果,在服用8周后,携带混合谱系白血病基因3(rs6475448)AA基因型的患者最大自主通气量显著高于基因型AG/GG患者。基因ATP结合盒C亚家族成员1编码的多重耐药蛋白1在LTC4跨膜转运中起重要作用,该基因的rs119774位点的多态性与LTRA的疗效相关,在ALOX5基因rs2115819位点上,与AG/AA基因型相比,携带纯合型GG基因患者在接受LTRA治疗后FEV1改善更多[13]。
与健康儿童相比,特异性哮喘儿童基因CA10中的rs967676片段(A>G)纯合子AA分布频率显著提高,rs967676的G等位基因与哮喘的严重程度有关,与AA型等位基因携带者相比,携带有1或2个G等位基因的患儿FEV1/用力肺活量更低,IgE、嗜酸性粒细胞表达更高。经LTRA治疗后钙黏蛋白相关蛋白rs1786929(T>A)TT型等位基因携带者的FEV1/用力肺活量值较杂合子或CC型纯合子高[35]。
5-脂氧合酶激活蛋白也是参与CysLTs合成的关键基因之一,是5-脂氧合酶活化的关键,哮喘患者外周血5-脂氧合酶激活蛋白mRNA的表达明显高于健康人[36]。郭青等[37]研究表明,脑浆型磷脂酶A2Ⅳ基因rs932476位点AA型患儿经孟鲁司特钠治疗后总有效率、γ-干扰素水平及肺功能指标均明显高于GG型患儿,且AA型患儿血清中白细胞介素4水平明显低于GG型患儿,提示该基因位点AA型患儿孟鲁司特钠疗效明显优于GG型患儿。此外,还有学者认为细胞色素P450酶系3A5基因rs776746位点、细胞毒性T淋巴细胞相关抗原4基因rs4553808位点AA基因型和A等位基因以及STAT3基因内含子11 rs2293152C/G的多态性都与LTRA疗效及哮喘的易感性相关[38-40]。
4 结 语
哮喘是一种复杂的多基因疾病,受遗传因素和环境因素的双重影响,目前LTRA的药物基因组学作用机制较为明确,如5-脂氧合酶、LTA4H、LTC4S通过干预白三烯作用通路上酶的代谢影响炎性介质CysLTs的生成,CYSLTR1和CYSLTR2通过干预CysLTs受体的合成影响其在细胞中的表达。但其他哮喘控制药物,如糖皮质激素的药物基因组学的具体作用机制仍未完全明确[41]。尽管近年来在使用药物基因组学改善哮喘疗效方面研究颇多,但由于研究人种、研究方法、研究内容上存在差异,很多临床研究难以重复,临床数据仍然缺乏,未来应结合药物基因组、遗传学、准确的疾病表型进行研究,为哮喘患者提供确最佳的治疗方案。
