‘红香酥’梨致腐性真菌的鉴定及其孢子萌发特性
2022-05-20张阳高聪聪程玉豆张晓宇张立新关军锋
张阳,高聪聪,程玉豆,张晓宇,张立新*,关军锋*
(1.河北省农林科学院生物技术与食品科学研究所,河北 石家庄 050051;2.河北省植物转基因中心重点实验室,河北石家庄 050051;3.山西农业大学食品科学与工程学院/农产品贮藏保鲜研究所,山西 太原 030031)
‘红香酥’梨是由‘库尔勒香梨’和‘鹅梨’杂交培育而成的新品种,因其果实品质上乘,果肉白嫩、多汁、香甜,深受消费者青睐,具有较高的食用和商业价值[1]。随着其栽培面积逐渐扩大,加强贮藏保鲜意义重大。近年来,在河北省、山西省等冷库发现,贮藏后期‘红香酥’梨果实腐烂时有发生,对贮藏企业造成了一定的经济损失。因此,为了更好地采取有效措施减轻或控制‘红香酥’梨采后腐烂,进行梨果实贮藏期间致腐病原物的鉴定和发病规律研究十分必要。
研究表明,果实贮藏病害主要以真菌为主,其中半知菌亚门真菌占据大多数,少量归属鞭毛菌亚门、接合菌亚门和子囊菌亚门[2]。目前关于梨果实贮藏期常发病害主要包括以下几种类型:由半知菌亚门真菌Penicillium expansum导致的青霉病[3-4],半知菌亚门真菌Alternaria alternate导致的黑斑病[5]、半知菌亚门真菌Botrytis cinerea导致的灰 霉 病[6]和 子 囊 菌 亚 门 真 菌Botryosphaeria do⁃thidea导致的轮纹病[7]等。然而,关于‘红香酥’梨果实采后病害研究尚不够系统。本研究在河北省代表性冷库中进行‘红香酥’梨果实病害调查和取样,采用组织分离法对典型病斑进行真菌的纯化,通过致病性检测明确了病原菌,进一步运用形态学和分子学手段对病原菌进行分类学鉴定,确定其分类地位,最后研究病原菌的生物学特性,为‘红香酥’梨果实采后病害的防治提供参考。
1 材料与方法
1.1 病果的采集与病原菌分离纯化
于2021年3 月采自河北省石家庄市赵县(114°58'28″ E,37°47'3″ W)、晋 州(115°3'4″ E,38°1'44″W)和辛集市(115°13'22″ E,37°56'13″ W),从各地冷库中采集‘红香酥’梨腐烂果样品后,在取样袋2 h 内运回实验室进行分离试验。参考方中达《植病研究方法第三版》[8]中的组织分离法,对典型病斑进行病原菌分离。从病斑处切取边长约为5 mm的小块方形果实组织(果皮带果肉),在70%酒精中浸泡几秒钟,转移至0.1%的酸性升汞中浸泡3 min 用无菌水洗3次,随后置于PDA 培养基。培养3~5 d 后,从菌落边缘挑取菌丝体转移到新的PDA 培养基上进行纯化培养,如此纯化2~3次,经显微镜检查证实无杂菌污染后即可确认为是单一的纯菌落。将纯化后的菌落接种到PDA 斜面培养基中,培养2 d 后置于4 ℃进行保存备用。
1.2 柯赫氏法则验证病原菌
使用10 mmol L-1PBS 缓冲液(生工生物工程股份有限公司,上海)将分离纯化的菌株重悬混匀,用四层纱布过滤后得到孢子悬浮液,使用PBS缓冲液将孢子浓度调整为5×106mL-1。以大小一致、颜色均匀、无物理损伤的‘红香酥’梨果实为材料,在2%次氯酸钠溶液中消毒2 min,用去离子水冲洗3次,然后在干净的工作台中风干,用无菌针在每个果实赤道针刺出2个3 mm×3 mm 的方形伤口。取5 μL 孢子悬液(5×106mL-1)接种在每个伤口处,采取保鲜膜保湿的方法,在恒温(25 ℃)、恒湿(95%)、黑暗条件下培养5 d。待果实发病后,按照1.1 中组织分离法从发病部位再次分离纯化出与首次分离中相同的真菌,即为病原菌。
1.3 菌株的形态特征鉴定
将纯化的菌株在PDA 培养基上活化,培养7 d后,挑取菌落边缘少许菌丝体放在加有一滴水的载玻片上,加盖玻片后在显微镜(OLYMPUS BX51,日本)下进行观察。观察孢子形态和产孢结构,拍照记录,使用ImageJ 软件(National Institutes of Health,Maryland,美国)测量孢子大小。
1.4 菌株的分子生物学鉴定
在分离得到4株病原真菌(Y001、Y002、Y003、Y004)的基础上,取0.1 g 活化的真菌菌丝,使用液氮研磨成粉末,使用真菌基因组DNA 快速抽提试剂盒(生工生物工程股份有限公司,上海)提取真菌的总DNA。根据文献报道并结合NCBI数据库中序列的收录情况,挑选了不同真菌中各自常用的保守基因作为构建该菌进化树的候选序列,例如真菌Penicillium中EF-1α 序列的报道不够充分,很多重要的种中没有相应的序列信息,因此选用BenA 基因进行建树,具体引物使用信息如下:使用引物ITS1/ITS4(扩增ITS 部分序列)和EF1-728F/EF1-986R(扩增EF-1α 部分序列)对菌 株Y001、Y002 和Y003 的DNA 进 行 扩 增[9],使用通用引物ITS1/ITS4 和Bt2a/Bt2b(扩增BenA部分序列)对菌株Y004 的DNA 进行扩增[10],引物由生工生物工程股份有限公司(上海)合成,序列见 表1。PCR反应体系为50 μL:2×T5 Super PCR Mix(擎科生物技术有限公司,天津)25 μL,菌株DNA为1 μL,引物各1 μL,ddH2O 23 μL。PCR体系为:98 ℃预变性3 min;98 ℃变性10 s,55 ℃退火10 s,72 ℃延伸10 s,30个循环;72 ℃延伸32 min;4 ℃保存。使用1%的琼脂糖检测PCR 扩增结果,120 V 电泳15 min,将条带大小正确的DNA样品送至生工生物工程股份有限公司(上海)进行双向测序。

表1 引物信息Table 1 Primer information in this study
测序结果经NCBI 网站在线BLAST 分析后,将菌株初步鉴定到属水平。接着将各菌株的ITS与EF-1α 或BenA基因序列合并,使用软件Mega5.1 构建系统进化树,使用邻接法(Neighbor-Joining),自展值(Bootstrap)为1000。
1.5 分生孢子萌发特性
1.5.1 分生孢子体内萌发观察
使用无菌刀片在经消毒的果实表面切一个5 mm 左右的圆形伤口,滴加5 μL 的孢子悬液(5×106mL-1)。将接种好的果实进行保鲜膜保湿,保持果实周围温度为25 ℃、湿度为95%。分别在接种后第0、24、48、72 h,使用超景深显微镜(DVM6A,Leica,德国)观察分生孢子在果实接种处的萌发状态,并拍照记录。
1.5.2 温度对分生孢子萌发的影响
在无菌载玻片上铺一层水琼脂(1%,w/v),待其凝固后滴加少量孢子悬液,并用玻璃棒轻轻地涂抹均匀。将制作好的带菌载玻片置于带有湿润滤纸的培养皿中保持湿度,随后在5、10、15、20、25、30、35 ℃的培养箱中恒温培养。每2 h 使用显微镜观察1次孢子萌发情况,并随机选取100个孢子统计萌发率,共观察10 h,重复3次。
1.5.3 湿度对分生孢子萌发的影响
按照浓硫酸湿度控制法,设置5个湿度条件分别为70%、80%、90%、100%、100%+ddH2O[11]。带菌载玻片的制作方法同上,随后在不同湿度的密封盒中恒温培养(25 ℃)。6 h 后显微镜观察孢子萌发情况,并随机选取100个孢子统计萌发率,重复3次。
1.5.4 光照对分生孢子萌发的影响
使用载玻片萌发法检测光照对分生孢子萌发的影响,使用LED日光灯为光源,光照条件分别为全光照、光暗交替(光照黑暗间隔时间为0.5 h)、全黑暗3个条件。带菌载玻片的制作方法同上,随后在不同光照条件培养箱中恒温(25 ℃)、恒湿(95%)培养。每2 h 使用显微镜观察1次孢子萌发情况,并随机选取100个孢子统计萌发率,共观察时间8 h。重复3次。
1.6 统计分析
使用GraphPad Prism 8 软件(GraphPad Inc.,CA,USA)对孢子萌发实验进行统计分析和作图,采用双因素方差分析(ANOVA)检验不同处理的差异显著性。
2 结果与分析
2.1 菌株分离及柯赫氏法则验证
通过组织分离法对果实腐烂部位进行病原菌的分离和纯化,共分离纯化出27个真菌菌株。经过对菌落形态的差异分析,最终选定了4种具有代表性的菌落形态不同的真菌进行后续研究。将纯化后的真菌回接至梨果实后,观察发现病斑与初始病斑基本一致,从病斑处再次分离得到了相同的病原菌。
发病过程观察结果表明,在高湿度条件下,所有菌株均会在发病部位产生大量的菌丝(图1)。菌株Y001 和Y002 均导致果实表面形成深褐色圆形病斑,病斑凹陷明显,病、健部位明显分离,菌株Y001 的病斑扩展速度相对于菌株Y002 的病斑扩展速度较慢,在发病后期,2种病斑表面长有黑褐色霉状物,且Y002 的菌丝量明显较多。菌株Y003初期在果实上导致浅褐色病斑,病斑凹陷明显,病、健部位明显分离,扩展速度较快,后期在病斑表面长有大量白色菌丝。菌株Y004 初期在果实上导致浅褐色圆形病斑,且病斑表面凹陷,边缘明显,扩展十分迅速,病斑表面长有白色小疣状发霉颗粒,后变为蓝色。因此,这4 株真菌均为导致‘红香酥’梨果实腐烂的病原物,分别被命名为Y001、Y002、Y003 和Y004。
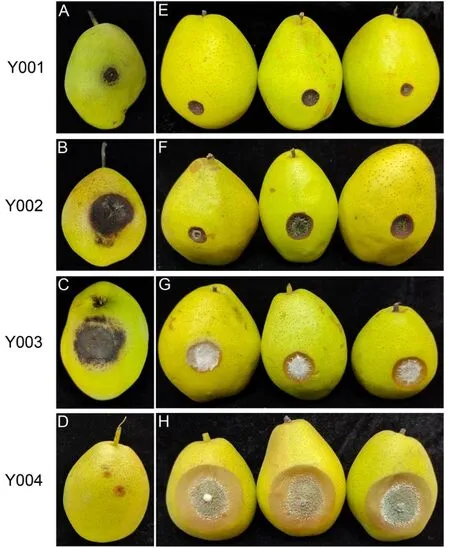
图1 ‘红香酥’梨果实真菌病害特征Fig.1 Disease symptoms of pathogenic fungi in‘Hongxiangsu’pear fruit
2.2 病原菌的形态学特征
经单孢子培养和显微观察菌丝及孢子形态,对病原菌的形态学特征进行鉴定(图2),菌株Y001 在平板上的菌落形态呈圆形,黄绿色菌丝,边缘比较整齐;分生孢子为纺锤形,黄褐色,有分隔且缢缩,大小为(14.170~37.060)μm×(5.798~10.254)μm;分生孢子梗为黄褐色,分生孢子着生在梗的末端。菌株Y002 的菌落为圆形,最初菌落颜色为白色,后变为墨绿色,呈绒毛状,边缘比较整齐;分生孢子为椭圆形或纺锤形,黄褐色,分隔处 缢 缩,大 小为(5.042~8.345)μm×(3.507~5.582)μm;分生孢子梗为黄褐色,分生孢子着生在其末端。菌株Y003 菌落为近圆形,菌落初为白色,后逐渐变为粉红色,边缘整齐;分生孢子为两端稍尖的镰刀形或卵圆形,无色单孢子,无分隔,大 小为(10.802~18.458)μm×(3.878~5.166)μm;分生孢子梗无色,分生孢子着生在其侧面。菌株Y004 的菌落呈圆形或不规则形状,初为白色,后逐渐变绿,边缘十分整齐;分生孢子为球形,无色单孢子,大小为(3.347~4.029)μm×(3.451~4.175)μm;分生孢子梗为帚状,无色,分生孢子在其顶端串生。

图2 ‘红香酥’梨果实病原真菌的形态特征Fig.2 Morphological characteristics of pathogenic fungi in fruits of‘Hongxiangsu’pear
2.3 病原菌的分子生物学鉴定
在形态学鉴定结果的基础上,提取真菌菌株Y001、Y002、Y003 和Y004 的总DNA,根据文献报道不同真菌所适合的引物进行PCR 扩增并测序。测序结果经NCBI 网站在线BLAST 分析,初步鉴定到菌株的属水平。进一步将各菌株测得的2个基因序列进行合并,使用NJ 方法构建系统进化树(图3),菌株Y001 和Y002 与Alternaria alternate、菌株Y003 与Fusarium proliferatum、菌株Y004 与Penicillium expansum菌株聚在一起。结合图1 的形态学鉴定结果,基本明确了4 株菌株的分类学地位。

图3 基于ITS 和EF-1α/BenA 基因序列的‘红香酥’梨果实病原真菌系统发育树Fig.3 Phylogenetic tree of pathogenic fungi from fruits of‘Hongxiangsu’pear based on ITS and EF-1α/BenA gene sequences
2.4 孢子体内萌发观察
在果实伤口注射分生孢子,利用超景深显微镜观察4个菌株在‘红香酥’梨果实中的孢子动态萌发规律。结果发现,在放大约400~700 倍时,4株菌在接种第24 h 后分生孢子均无明显差异迹象,而在接种第48 h 后可以明显看到大量的分生孢子萌发产生的菌丝,在接种第72 h 后菌丝量增长迅速,几乎掩盖住整个伤口。菌株Y003 和Y004在放大约700 倍时,可以明显看到其特异的分生孢子梗等部位(图4)。
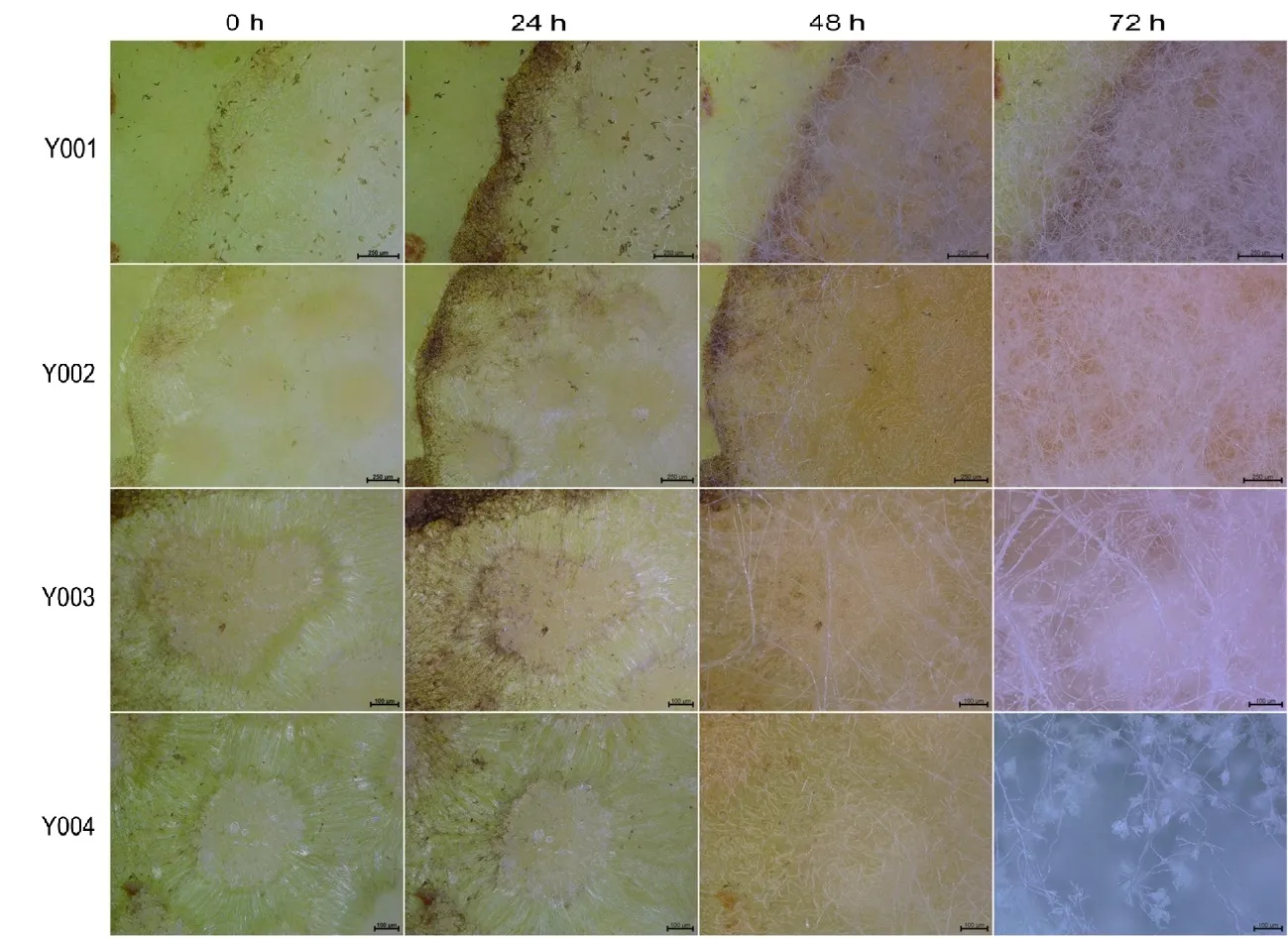
图4 超景深显微镜观察病原真菌孢子在‘红香酥’梨果实上的萌发Fig.4 Observation on spore germination of four pathogenic fungi in‘Hongxiangsu’pear fruit by super depth of field microscope
2.5 温度、湿度和光照对分生孢子体外萌发的影响
通过检测不同温度、湿度和光照条件下分生孢子的萌发率,以明确病原真菌分生孢子的生物学特征(图5)。通过测定不同温度下分生孢子随时间的萌发率,菌株Y001 分生孢子在第4 h 时开始大量萌发,并且温度为25、30 ℃时萌发率最高,第8 h 分生孢子的萌发接近100%。因此,Y001 的孢子最佳萌发温度为25~30 ℃。菌株Y002 的分生孢子也是在第4 h 时开始大量萌发,且在20、25和30 ℃时萌发率最高,第6 h 萌发率接近100%。因此,Y002 的孢子最佳萌发温度为20~30 ℃。菌株Y003 在第4 h 时,20、25、30 和35 ℃均略有萌发且25 ℃和30 ℃萌发最多,第6 h 时,孢子在25、30和35 ℃大量萌发,在第8 h 达到最高。因此,Y002的孢子最佳萌发温度为25~30 ℃。菌株Y004 在前6 h 时均未萌发,第8 h 时温度高于20 ℃的孢子均有少量萌发,第10 h 时大量萌发,且温度为25 ℃时萌发率最高。因此,Y004 的孢子最佳萌发温度为25 ℃。

图5 温度对‘红香酥’梨4种病原菌孢子萌发率的影响Fig.5 Effect of temperature on spore germination rate of 4 pathogenic fungi in‘Hongxiangsu’pear fruit
通过浓硫酸控制湿度法,检测恒温培养8 h 后不同湿度条件对分生孢子萌发的影响。结果如图6所示,当环境湿度为70%时,除菌株Y001 的分生孢子有微量萌发外(7%),其余3 株真菌的分生孢子均不萌发;当湿度为80%时,除菌株Y004 不萌发外,其余3个菌株均有萌发(32%~49%);当湿度为90%时,4个菌株均有萌发,其中菌株Y002 的萌发最高(79%),Y004 的萌发率最低(30%);当湿度为100%时,菌株Y003 的萌发率达到了90%,Y004 的萌发率为40%;当湿度为100%+ddH2O时,除了菌株Y004 的萌发率略低外(77%),其余3个菌株的孢子萌发率均接近100%。

图6 湿度对‘红香酥’梨4种病原菌孢子萌发率的影响Fig.6 Effect of humidity on spore germination rate of 4 pathogenic fungi in‘Hongxiangsu’pear fruit
当使用全光、全暗和光暗交替3种光条件处理分生孢子时,除了菌株Y003 外,其余菌的分生孢子萌发不受光照的影响(图7)。菌株Y003 在受光照6 h 后开始大量萌发,而当光照时间达到8 h 时萌发率可达58%。光暗交替同样可以促进菌株Y003 分生孢子的萌发,在第8 h 时萌发率达到60%,这说明,光照可促进菌株Y003 的孢子萌发。

图7 光照对‘红香酥’梨果实中4种病原菌孢子萌发的影响Fig.7 Effect of light on spore germination of 4 pathogenic fungi in‘Hongxiangsu’pear fruit
3 讨论
果实病害是长期以来困扰果农和贮藏企业的重要问题。本研究分离并鉴定出了4 株导致‘红香酥’梨果实贮藏期的病原物,分别为2个链格孢属真菌Alternaria alternateY001 和A. alternateY002,1个镰刀菌属真菌F. proliferatumY003 和1个青霉菌属真菌P. expansumY004。
链格孢的寄主范围很广,且传播方式多样,侵染后会造成严重危害[12-14]。其中,由链格孢引起的梨黑斑病是世界范围内的重要病害,不仅侵害果树上叶片和果实,还对贮藏期的果实健康造成严重威胁[15-16]。我国梨产区的主要链格孢主要分属6个种:细极链格孢(A. tenuissima)、细链格孢(A.alternata)、乔木链格孢(A. arborescens)、梨黑斑链格孢(A. gaisen)、棉链格孢(A. gossypina)、长柄链格孢(A. longipes),其中A. tenuissima和A. al⁃ternata为优势种[17]。本研究分离鉴定出2 株引起‘红香酥’梨贮藏期病害的链格孢菌Y001 和Y002,经形态学和分子学鉴定均属于细链格孢(A. alter⁃nate),并且试验发现其孢子萌发受温度和湿度影响较大,对光照不敏感(图7)。
镰刀菌在自然界中广泛存在,不仅在小麦、玉米等大田作物上危害严重,而且可侵染多种果蔬作物,包括苹果[18]、香蕉[19]、冬枣[20]、土豆[21]、洋葱和大蒜[22]等。然而,很少有关于镰刀菌引起的梨果实病害报道。近年,在翠冠梨中报道了1个可以引起果实腐烂的镰刀菌,经形态和分子鉴定为层出镰刀菌F. proliferatum[23],与本研究中分离得到的菌株Y003 归为同1个种。因此,镰刀菌在梨贮藏中所引起的病害应引起人们的重视。
青霉菌是果蔬采后贮藏中的常发真菌病害,主要由扩展青霉(P. expansum)、指状青霉(P.digitatum)和意大利青霉(P. italicum)所引起。本研究从‘红香酥’梨中分离纯化出的菌株Y004 经形态学和分子学鉴定为扩展青霉(P. expansum),与其它品种梨果实中报道的青霉菌的种类一致[24-25],进一步说明扩展青霉在采后梨果实病害中分布十分广泛。
分生孢子是子囊菌门真菌寄生在宿主上时产生的无性孢子,孢子萌发在真菌病原物的致病过程中具有重要作用[11,26]。与传统的光学显微镜和电子显微镜不同,本研究通过运用超景深显微镜,直观地观察到4 株病原菌的分生孢子在梨果实体内逐步萌发成为菌丝的动态过程,分生孢子在48 h即可在果实伤口产生大量的菌丝,说明在果实贮藏时期应加强对冷库等贮藏空间的消毒等处理。孢子在萌发过程中受到多种环境因素的影响,明确孢子萌发条件,有利于通过调控贮藏条件来减少病害发生[27-29]。本研究通过控制环境温度的变化检测孢子萌发活性,结果发现高温显著抑制了4种真菌的孢子的萌发,这可能是由于高温导致孢子细胞内大量活性氧自由基(ROS)的累积,进而对体内的大分子蛋白和脂类造成过氧化损伤,从而表现出抑制孢子萌发效果[30]。另外,光照明显影响了F. proliferatumY003 的孢子萌发,这与光照可能是某些真菌体内的生物途径的转录激活与抑制因子有关[31]。这些结果表明,在‘红香酥’梨果实的采后和货架期贮藏中应尽量保持低温、低湿和避光状态,从而降低A. alternata、F. pro⁃liferatum和P. expansum等真菌的孢子萌发活性,减少腐烂发生。
4 结论
组织分离法从河北省代表性冷库的‘红香酥’梨果实上分离出27 株真菌菌株,赫氏法则验证了4株典型病原真菌的致病性;综合其形态学和分子学特征明确了病原菌的分类学地位,4 株真菌分别被鉴定为Alternaria alternateY001,Alternaria al⁃ternateY002,Fusarium proliferatumY003 和Peni⁃cillium expansumY004;超景深显微镜观察发现梨果实上的分生孢子在48 h 内即可萌发出大量的菌丝;温度和湿度对4 株菌的孢子萌发有显著影响,而光照可促进菌株Y003 的孢子萌发。
