南极磷虾蛋白营养与功能特性及食用安全性研究进展
2022-05-13刘柯欣林松毅胡胜杰张秀敏
刘柯欣,林松毅,2,3,胡胜杰,张秀敏,孙 娜,2,3,*
(1.大连工业大学食品学院,辽宁 大连 116034;2.国家海洋食品工程技术研究中心,辽宁 大连 116034;3.海洋食品精深加工关键技术省部共建协同创新中心,辽宁 大连 116034)
南极磷虾(Euphausia superba)是地球上生物量最大的单种生物资源之一,总生物量约107~108t[1]。据南极海洋生物资源保护委员会统计,近10 年世界各国的南极磷虾总捕捞量达2.23×106t,其中2018年捕捞量为3.13×105t[2-3]。南极磷虾是挪威、乌克兰、韩国、日本、智利、俄罗斯等世界各国开发的重要经济战略资源。我国对南极磷虾资源也极为重视,于2009年将其列为“国家经济战略资源”,并发布探捕令,每年以上万吨的捕获量进入我国市场。
南极磷虾因富含优质蛋白,被认为是地球上最大、也是最后一个潜在动物性蛋白资源库[4]。国内外针对南极磷虾蛋白的基础研究和应用研究取得了积极进展,但是南极磷虾蛋白尚未得到充分的开发利用,市面上的产品仍以虾粉、虾干等用于生产饲料的初级产品为主,缺乏高值化精深加工产品。因此,本文系统综述了南极磷虾蛋白的营养特性及其评价、功能特性及其改性、功能活性及其作用机制和食用安全性问题,以期为南极磷虾蛋白资源的高值化利用提供理论依据,为我国优质动物性蛋白资源的补充供应、新型功能性食品的开发和南极海洋生物资源的合理利用提供参考。
1 南极磷虾蛋白的营养特性及其评价
1.1 南极磷虾蛋白的营养特性
南极磷虾含有丰富的蛋白质且氨基酸种类齐全,具有较高的营养价值和广阔的开发前景。南极磷虾的蛋白质占其干质量的60%~65%,是一种完全蛋白质[5]。据报道,南极磷虾蛋白质的生物价高于其他肉类蛋白质和牛奶蛋白质(如酪蛋白),但低于鸡蛋蛋白质[6]。南极磷虾不同产品中的蛋白质含量存在较大差异,研究显示在普通虾粉、脱脂虾粉、脱氟虾粉、虾肉、虾糜这5种南极磷虾产品中,脱氟虾粉的蛋白质含量最高,营养价值最高[7];另外,相比于南极磷虾蛋白,磷酸化后的南极磷虾蛋白相对含量更高,在90%以上[8]。
南极磷虾蛋白含量及其氨基酸组成易受多种因素的影响。一是受捕捞季节的影响。南极磷虾的捕捞季节在3~8月,其蛋白质含量在5月份最低,3月份最高,其他月份没有很大差异;氨基酸含量随季节变化,一般氨基酸总量、游离氨基酸总量、谷氨酸含量和天冬氨酸含量均在3月份达到最高水平,6月份最低[9]。二是受生长阶段的影响。研究发现,体长小于40 mm未成熟磷虾个体的蛋白质含量显著高于体长大于40 mm的成熟个体或高龄个体,这可能是因为未成熟个体吸收的营养物质主要以蛋白质和脂肪的形式储存在生物体内,而成熟个体吸收的营养物质则转化成精子和卵子[10]。三是受加工方式的影响。不同加工方式处理的南极磷虾蛋白质损失情况也有所不同,油炸加工的蛋白质损失最大,其次为水煮,烘烤加工的蛋白质损失最小[11];南极磷虾经微波加热比水浴加热更有利于滋味的形成,微波加热测得的游离氨基酸含量、必需氨基酸含量和鲜味氨基酸含量高于水浴加热[12]。
1.2 南极磷虾蛋白的营养评价
对南极磷虾蛋白进行营养评价主要是通过必需氨基酸种类和数量的评分或通过喂食不同蛋白质所得大鼠体质量增加率来考量的[13-15]。
南极磷虾蛋白含有人体必需的8种氨基酸[16],必需氨基酸总量占总蛋白质含量的20%以上,氨基酸评分在0.85~1.00之间,虽然低于参考蛋白鸡蛋蛋白的氨基酸评分,但南极磷虾蛋白提供的必需氨基酸量超过了健康成年人所需量[5],也高于大豆浓缩蛋白含量,与乳蛋白含量相近。其中,谷氨酸含量最高,其次是天冬氨酸,组氨酸含量最低,且其含量也符合婴儿对氨基酸的需求。研究发现,南极磷虾蛋白、蛋白分离物[17]及磷酸化蛋白[8]中赖氨酸含量也相对较高,对于以谷物食品为主的我国居民而言,摄入南极磷虾有助于补充赖氨酸[18]。另外,南极磷虾蛋白中所含的支链氨基酸(缬氨酸+亮氨酸+异亮氨酸)与芳香族氨基酸(苯丙氨酸+酪氨酸)的比值接近正常人和哺乳动物支链氨基酸/芳香族氨基酸,说明南极磷虾蛋白中所含的氨基酸比例平衡[19],合理摄入南极磷虾有利于维持机体的营养与代谢平衡。联合国粮食及农业组织(Food and Agriculture Organization of the United Nations,FAO)/世界卫生组织(World Health Organization,WHO)/联合国大学(United Nations University,UNU)以婴儿对氨基酸的需要量为最低限度提出:必需氨基酸与总氨基酸含量的比值在40%左右、必需氨基酸与非必需氨基酸含量的比值达到60%以上的蛋白质为优质蛋白[7,20]。南极磷虾蛋白的必需氨基酸/非必需氨基酸含量比值、蛋白分离物必需氨基酸含量和磷酸化蛋白的氨基酸组成均符合FAO/WHO/UNU提出的理想模式[8,17,20],由此可见,南极磷虾蛋白是一种优质蛋白。
通过动物实验对蛋白质价值进行营养评价比氨基酸评分更准确一些,因为前者包括消化和吸收方面,能更好地反映人体的消化吸收程度。Gigliotti等[14]用南极磷虾浓缩蛋白和酪蛋白分别饲喂小鼠,结果表明,浓缩蛋白的消化率、蛋白质消化率校正后的氨基酸评分(protein digestion corrected for amino acid score,PDCAAS)和蛋白质功效比(protein efficiency ratio,PER)与酪蛋白相当。一些研究数据还表明,饲喂南极磷虾蛋白质组大鼠的肝脏质量、胴体干物质和血红蛋白含量与饲喂酪蛋白组相比没有显著性差异[13]。Suzuki等[15]还进行了人体试食实验,利用南极磷虾蛋白和全蛋蛋白质喂食5名成年男子,结果表明,煮熟的南极磷虾蛋白质净利用率是全蛋蛋白质的89%。通过以上氨基酸评分和动物实验可以得出,南极磷虾蛋白是高质量蛋白的良好来源。
不同产品形式南极磷虾的氨基酸组成如表1所示。
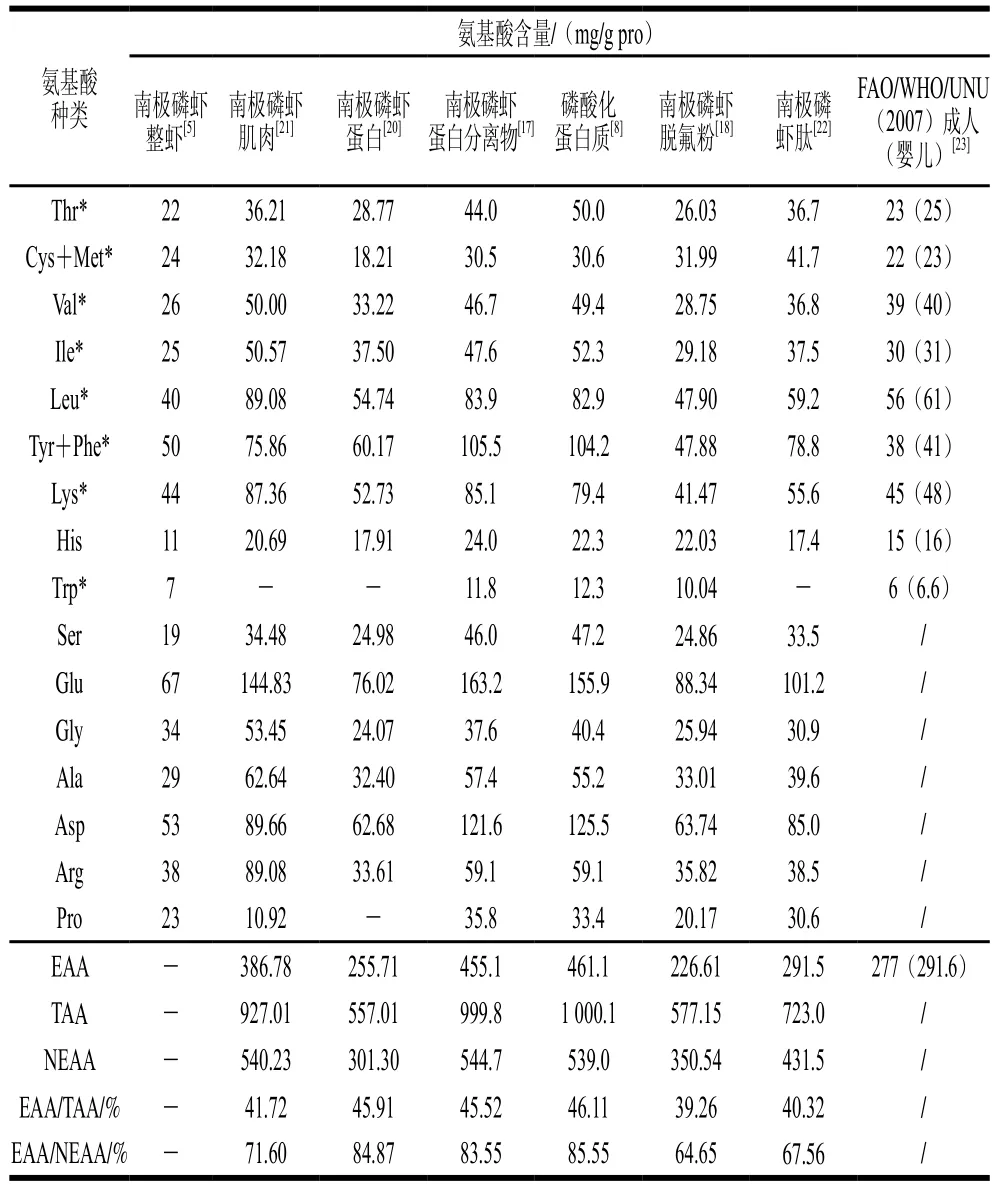
表1 不同产品形式南极磷虾的氨基酸组成Table 1 Amino acid composition in different types of Antarctic krill products
2 南极磷虾蛋白的功能特性及其改性
2.1 南极磷虾蛋白的功能特性
在对南极磷虾蛋白的研究和开发应用过程中,功能特性尤为重要。南极磷虾蛋白的功能特性主要包括溶解性、乳化性、凝胶性、稳定性等,这些性质对于南极磷虾蛋白的应用具有重要意义。
2.1.1 溶解性
蛋白质的溶解性与等电点(pI)有关,通常表现为等电点附近溶解度最低,偏离等电点溶解度升高[24]。南极磷虾蛋白的等电点为pH 4.6,此时溶液中分子间作用力减弱,蛋白质分子因没有相同电荷的排斥而凝聚沉降。南极磷虾蛋白在pH 4.6时显示最低的溶解度,仅为0.61%;在pH 2.6~4.6和pH 4.6~6.6范围内,南极磷虾蛋白溶解度急剧增加,主要是因为在高于或者低于pI的酸碱度条件下,蛋白质分子根据酸碱度带正或负电荷,分子间的静电排斥作用有助于南极磷虾蛋白溶解;pH>6.6时,有90%的南极磷虾蛋白发生溶解[17]。南极磷虾蛋白质良好的溶解性是其他功能特性的基础。
相比于南极磷虾虾肉和虾糜,南极磷虾蛋白的溶解度是最高的[25]。南极磷虾肌肉外部组织中含有较多的水溶性肌浆蛋白,提取蛋白质需经多次酸沉碱溶,肌浆蛋白随之进入上清液后沉淀,因此赋予了南极磷虾蛋白较好的溶解性。
2.1.2 凝胶性
凝胶性是蛋白质在食品应用中的一个重要功能特性,南极磷虾肌球蛋白对其凝胶性的发挥起着非常关键的作用。南极磷虾分离蛋白在不添加盐或其他添加剂且质量浓度超过100 g/L的情况下能形成凝胶,这种凝胶在重复加热/冷却循环条件下表现出热可逆行为[17]。在此基础上,Wang Yanchao等[26]通过研究热诱导南极磷虾蛋白凝胶的流变行为和微观结构来评价其增稠作用,为开发食品增稠剂提供了理论基础。经过热处理的南极磷虾蛋白在pH 5.5~8.6的范围内可以形成流体凝胶,其中,在pH值为5.5和7.2时的凝胶强度明显高于其他pH值下的凝胶强度;在pH 7.2时,液体凝胶的表观黏度明显高于其他pH值条件下形成的凝胶。但是,南极磷虾蛋白的凝胶性易受其内源性蛋白酶的影响,由于南极磷虾蛋白内源性蛋白酶活性非常强,自身很容易被水解,从而产生自溶,失去其功能特性,尤其是凝胶特性,这阻碍了南极磷虾商业化加工和重组磷虾蛋白产品(如虾滑等)的开发[27]。
2.1.3 乳化性和乳化稳定性
植物蛋白作为天然乳化剂在食品工业领域引起极大的关注[28-30]。事实上,南极磷虾蛋白中含有的必需氨基酸比植物蛋白多,是一种很有开发前景的食品级乳化剂资源。南极磷虾蛋白的疏水相互作用、二硫键和β-折叠结构对蛋白质聚集体的形成和稳定有重要影响。当pH值远离等电点时,分子间排斥力的增加大大阻碍了蛋白质聚集,使分子尺寸更小、灵活性更高,促进蛋白质在油-水界面的吸附。在此基础上制备的南极磷虾蛋白稳定乳化剂颗粒粒径小于20 μm,与已有的蛋白质或生物聚合物稳定的乳化剂相比,贮存30 d后仍能表现出较好的稳定性[31]。由此可见,新型南极磷虾蛋白稳定乳化剂的构建可以提高南极磷虾蛋白资源在食品工业中的高附加值利用。
2.1.4 起泡性和泡沫稳定性
蛋白质的起泡性在食品领域有着极为重要的应用,研究发现南极磷虾蛋白具有一定的起泡能力和泡沫稳定性。南极磷虾蛋白的起泡性和泡沫稳定性高于大豆分离蛋白,低于乳清分离蛋白,且均在质量浓度为40 mg/mL时最高[32]。相比于南极磷虾虾糜和虾肉,南极磷虾蛋白具有较好的溶解性,溶液中有效蛋白浓度较大,吸附到气-液界面的蛋白质分子较多,有利于界面性质的发挥,因此南极磷虾蛋白的起泡性最好。但南极磷虾蛋白的泡沫稳定性低于虾肉,所以南极磷虾蛋白的起泡性和泡沫稳定性没有必然联系[25]。
2.1.5 吸水性和吸油性
吸水性是南极磷虾蛋白很重要的功能特性之一,同时南极磷虾蛋白对油脂的吸收能力在很大程度上影响着以南极磷虾为原料食品的外观、色泽和口感。南极磷虾蛋白的吸水性很弱,但吸油性优于乳清浓缩蛋白和大豆分离蛋白[32],这主要与蛋白质分子表面所含基团的性质有关。磷虾蛋白吸水性在等电点处最低,主要是因为等电点处蛋白质发生聚集沉淀,表现出最弱的吸水能力[24]。
南极磷虾蛋白的功能特性总结如表2所示。

表2 南极磷虾蛋白的功能特性Table 2 Functional properties of Antarctic krill proteins
2.2 南极磷虾蛋白的改性方法
南极磷虾蛋白在常温下溶解度很低、凝胶性也较弱,限制了其在食品行业中的应用。因此,对南极磷虾蛋白进行改性可提高其功能性,扩大其应用范围。
2.2.1 极磷虾蛋白的物理改性
近些年,物理改性因其绿色、安全、成本低等优点广泛应用于改善蛋白质功能特性方面[32,37-39],但是目前研究报道涉及到对南极磷虾蛋白的物理改性方式很少,主要集中在运用超声波对南极磷虾蛋白进行改性。
超声处理能够改变南极磷虾蛋白的功能特性,通过改变蛋白质的空间结构,暴露亲水基团,提高溶解度和吸水性,但会导致吸油性降低[36]。溶解能力改善的同时,南极磷虾蛋白的乳化性也得到提升。在没有超声辅助的情况下,含2%南极磷虾蛋白的稳定乳液在4 ℃下储存30 d后显示出明显的乳状液分层现象,然而由超声处理的蛋白质稳定乳液的乳化程度显著降低,特别是在处理20 min和30 min之后[35]。此外,超声诱导南极磷虾蛋白乳液产生的无序和不规则结构也有助于乳液的稳定性。超声处理还改变了磷虾蛋白质的表面疏水性,促进了蛋白质分子在水-空气界面的展开和界面膜的形成[36],提高了溶解度,减小了粒径,有利于α-螺旋和β-折叠的转变,促进了磷虾蛋白的表面疏水基团和二硫键的形成,有助于增强磷虾蛋白质的起泡能力、泡沫稳定性[35]。
2.2.2 南极磷虾蛋白的化学改性
南极磷虾蛋白的化学法改性研究应用较多,其方法简单、效果显著、易实现工业化[40],研究重点主要为磷酸化、糖基化等。
对食品蛋白质的功能特性来说,磷酸化被证明是一种很有效的方式。南极磷虾蛋白磷酸化后,其溶解性、吸水性、乳化性、乳化稳定性、起泡性、泡沫稳定性均有所提高,且变化趋势相同,基本遵循“接近等电点时低,偏离等电点时高”的规律,但是也有例外,即磷酸化后的南极磷虾蛋白吸油率低于未磷酸化的蛋白,泡沫稳定性在等电点附近时最高,而偏离等电点时降低[24]。
糖基化改性可以提高南极磷虾蛋白质的功能特性,并保持性质稳定[41]。南极磷虾蛋白糖基化对其功能特性的影响主要取决于两个因素:一个是糖基供体,另一个是糖基供体与南极磷虾蛋白的混合时间。采用葡萄糖对南极磷虾蛋白进行糖基化改性可以提高其溶解性、乳化性和乳化稳定性,其分别在葡萄糖与蛋白反应45、60、15 min时达到最高;采用D-半乳糖对南极磷虾蛋白改性60 min可以提高其吸水性,但是对起泡性的改善效果不是很明显;采用D-木糖改性能够提高南极磷虾蛋白的吸油性,改性30 min时吸油性最大[32,42]。由此可见,可以根据不同的应用领域或目的,选择合适的改性时间和糖基供体对南极磷虾蛋白进行改性,以获得高质量产品。
南极磷虾蛋白与多糖结合可以改善其凝胶特性和持水能力。用等电点溶解/沉淀法提取的南极磷虾分离蛋白由于内源性蛋白酶的存在,形成的凝胶质地非常差,显示出不均匀、多孔和海绵状的微观结构。κ-卡拉胶的加入显著提高了南极磷虾蛋白的硬度、黏性、弹性和咀嚼性,其存在于南极磷虾蛋白的间隙中,主要负责凝胶网络的形成,与水结合形成氢键,提高持水能力进而形成空隙均匀规则的凝胶[27]。
2.2.3 南极磷虾蛋白的生物改性
酶法改性被认为是一种比较优异且有潜力的蛋白质生物改性方式,能够改善南极磷虾蛋白的溶解性、吸水性、起泡性和泡沫稳定性,而改性后磷虾蛋白的吸油性、乳化性和乳化稳定性均有一定程度的下降[32]。酶法改性过程中,南极磷虾蛋白的水解度不断增加,水解产生的较小肽暴露出极性基团,能与水形成氢键,使亲水性提升,溶解度和吸水性增加;小肽在界面上快速移动和吸附,但在降低界面张力方面效率较低,不能像大分子蛋白一样在界面上展开和重新定向,所以酶法改性后南极磷虾蛋白的起泡性和泡沫稳定性较高,乳化性和乳化稳定性较低[43]。
南极磷虾蛋白改性方式及其效果总结如表3所示。

表3 南极磷虾蛋白改性方式及其效果Table 3 Modification of Antarctic krill proteins and its effects
3 南极磷虾蛋白衍生肽的功能活性及其作用机制
南极磷虾蛋白衍生肽一般通过酶解反应获取,且不同氨基酸组成的肽链具有不一样的功能活性[44]。目前,已从南极磷虾蛋白中获取多种生物活性肽,其功能活性包括抗氧化活性、促矿物质吸收活性、抗骨质疏松活性、降血压及降血糖活性、抗炎活性等。
3.1 抗氧化活性
迄今为止,抗氧化肽是研究人员探索最多的一种功能性肽,抗氧化活性与氨基酸组成[45]或序列[46]有关,如含有疏水性氨基酸[47]、芳香族氨基酸[48]、-Glu-Leu-[49]、His、Pro[50]等会提升肽的抗氧化性。如表1所示,南极磷虾肽含有较高含量的芳香族氨基酸(苯丙氨酸、酪氨酸)、谷氨酸、脯氨酸等[22],因此,南极磷虾肽具有潜在的抗氧化活性。目前已从南极磷虾蛋白中分离获取了多种具有抗氧化活性的多肽。郑景如等[51]采用酶解-超滤-Sephadex G-25凝胶层析制备南极磷虾抗氧化肽组分,发现3~10 kDa南极磷虾肽组分的1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-picrylhydrazyl,DPPH)自由基清除能力最高,分子质量低于3 kDa南极磷虾肽组分的2,2’-联氮-双-(3-乙基苯并噻唑啉-6-磺酸)(2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid),ABTS)阳离子自由基清除率、抗氧化能力指数、超氧化物歧化酶和过氧化氢酶活力显著高于其他肽组分。同样地,张元元[44]利用不同分子质量的南极磷虾多肽分别进行体内、体外抗氧化实验,结果表明不同分子质量多肽具有不同程度的抗氧化能力,其中分子质量不超过5 ku的多肽抗氧化能力最佳。另外,Lan Chen等[52]采用连续酶水解工艺从南极磷虾蛋白中制备出高F值(支链氨基酸与芳香族氨基酸的物质的量比大于20)寡肽,具有高自由基清除活性、还原能力和抑制脂质过氧化的能力。若将南极磷虾肽应用在功能性食品和食疗食品的开发中,有望提高食品的抗氧化性能。
3.2 促矿物质吸收活性
南极磷虾肽富含谷氨酸、天冬氨酸、精氨酸等,易与钙离子[53-54]、锌离子[55]、铁离子[56-57]螯合形成肽-钙、肽-锌、肽-铁复合物,可以明显提高金属离子在胃肠道环境下的溶解度和稳定性,增加金属离子吸收利用率,改善人体矿物质元素缺乏情况。
Hou Hu等[54]从南极磷虾蛋白水解物中纯化出Val-Leu-Gly-Tyr-Ile-Gln-Ile-Arg,研究发现钙离子螯合至Gln、Ile和Arg残基的氨基或羰基上,与氨基氮原子和水分子相比,钙离子更容易与带负电荷的羧酸基团形成配合物。锌可以与南极磷虾肽的羧基氧、氨基氮[55]和羰基等[58]官能团螯合,螯合过程中肽与锌的相互作用破坏了肽表面原有的致密结构,螯合后肽锌复合物的粒径、ζ电位增加,分子质量低于1 000 Da的肽含量降低,分子质量高于1 000 Da的肽含量增加。利用螯合后的肽锌复合物、硫酸锌和葡萄糖酸锌分别进行模拟胃肠道消化实验,在胃消化阶段,三者的锌溶解度基本一致,进入肠消化阶段时,硫酸锌的溶解度急剧下降,其次为葡萄糖酸锌,而肽锌复合物溶解度保持在最高水平。由此可见,南极磷虾肽锌复合物在胃肠道消化过程中具有良好的稳定性[55]。铁离子主要结合到南极磷虾肽Asp和Glu的羧基或Ser和Thr的磷酸基团上,使肽结构发生折叠,以纳米级球形颗粒通过胃肠道,并在模拟胃肠道消化过程中表现出显著的转运活性[57]。研究人员进一步筛选出具有更高铁结合能力的南极磷虾九肽(DTDSEEEIR),Fe3+螯合到九肽的羧基和氨基上形成的簇状九肽-铁螯合物粒径较小,为新的晶体形态[59]。在胃肠道消化过程中,南极磷虾九肽能够保持一定程度的稳定性((93.89±1.37)%),且能够显著增强铁离子在肠道中的吸收[59-60]。综上所述,南极磷虾肽通过特定基团或残基与钙、锌、铁3种金属离子螯合形成矿物螯合肽,在模拟胃肠道消化过程中能够保持一定程度的稳定性,提高矿物质生物利用度,具有成为新型矿物质补充剂的潜力。
3.3 抗骨质疏松活性
随着我国人口老龄化不断加剧,越来越多的人受到骨质疏松症的影响,而南极磷虾无论是对原发性骨质疏松还是继发性骨质疏松均有较明显的改善效果。
原发性骨质疏松破坏了钙磷代谢的平衡,导致骨吸收增加、骨小梁和皮质骨的骨量严重损失,而南极磷虾磷酸化肽能以剂量依赖性地抑制骨吸收增加,显著逆转骨质疏松引起的骨吸收标志物的变化,对其骨小梁和皮质骨强度也均有明显改善作用[8]。南极磷虾蛋白水解获得的南极磷虾肽可以促进前胚细胞MC3T3-E1细胞的增殖,通过激活骨形态发生蛋白2(bone morphogenetic protein2,BMP2)/Smads和Wnt/β-连环蛋白分子途径来改善老年骨质疏松症,从而促进骨形成[61]。南极磷虾磷酸化肽还可以通过促进软骨骨痂的转化和骨痂的重塑、调节软骨内骨化相关基因的表达来促进去卵巢诱导的骨质疏松小鼠胫骨骨折的愈合[62],这为南极磷虾蛋白作为营养成分用于骨保护功能性食品的开发提供理论依据。
除了对原发性骨质疏松的改善作用,Han Lihua等[63]还研究了南极磷虾磷酸化肽对地塞米松诱导的继发性骨质疏松症的影响。南极磷虾磷酸化肽通过抑制丝裂原活化蛋白激酶磷酸酶(mitogen-activated kinase phosphatase,MKP)-1和大麻素受体(cannabinoid receptor,CB)1的基因表达,提高关键因子p38、细胞外信号调节激酶(extracellular signal-regulated kinase,ERK)、磷脂酰肌醇-3-激酶(phosphatidylinositol-3-kinase,PI3K)、丝氨酸/苏氨酸特异性蛋白激酶(protein kinase B,AKT)、β-连环蛋白以及成骨核转录因子Runx2和OSX的表达,激活了下游成骨相关的丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)和PI3K/AKT/糖原合成酶激酶(glycogen synthase kinase,GSK-3β)信号通路,显著改善骨转换状态,增加小鼠的骨小梁质量,减少骨丢失和微结构的退化,加速骨形成,逆转了地塞米松对小鼠骨微结构的有害作用。这为南极磷虾生物活性肽作为改善骨代谢的天然活性替代品提供了理论依据。
3.4 降血压、降血糖活性
早在1992年,Kawamura等[64]在脱脂南极磷虾蛋白质的消化水解物中分离并鉴定出有助于控制高血压的血管紧张素转换酶(angiotensin converting enzyme,ACE)抑制剂肽Lys-Leu-Lys-Phe-Val。Hatanaka等[65]通过改进前人的方法,从去皮的南极磷虾尾肉蛋白中制备了有效的ACE抑制肽Val-Trp和Leu-Lys-Tyr,其对自发性高血压大鼠具有抗高血压作用。
南极磷虾蛋白水解物还被认为是糖尿病治疗中二肽基肽酶IV(dipeptidyl peptidase IV,DPP-IV)抑制肽的一种天然有效来源。Ji Wei等[66]通过超滤和色谱法从南极磷虾蛋白水解物中纯化出两种DPP-IV抑制肽(Ala-Pro(AP)和Ile-Pro-Ala(IPA)),并通过分子对接技术将作为配体的AP和IPA与蛋白质靶标的DPP-IV对接,确定了结合位点。Ji Wei等[67]用相同的方法从南极磷虾蛋白水解物中分离出两条对ACE和DPP-IV有双重抑制活性的肽,分别是Lys-Val-Glu-Pro-Leu-Pro和Pro-Ala-Leu,通过二者抑制ACE和DPP-IV活性的能力来调节血压和血糖。因此,南极磷虾蛋白水解产物可用于生产降血压和控制血糖的功能性产品。
从南极磷虾蛋白中分离出的肽不仅可以降血压、降血糖,还可能通过精氨酸供应一氧化氮(NO)来影响动脉粥样硬化的发展从而保护内皮功能[68]。Zhao Yuqin等[69]从南极磷虾蛋白水解物中成功纯化了8种新肽,鉴定为Trp-Phe、Tyr-Arg-Lys-Glu-Arg、Tyr-Arg-Lys、Val-Asp、Tyr-Lys-Asp、Phe-Gln-Lys、Phe-Ala-Ser和Phe-Arg-Lys-Glu,其中小分子质量的Trp-Phe和Phe-Ala-Ser对ACE显示出显著的抑制活性,而且肽诱导NO的产生和抑制内皮素-1的表达保护了血管。这些发现均显示了南极磷虾蛋白衍生肽具有预防高血压和动脉粥样硬化的潜力。
3.5 抗炎活性
目前有研究报道,南极磷虾肽对骨关节炎软骨有保护作用,并能够抑制内侧半月板失稳诱导的骨关节炎中软骨基质的损失。南极磷虾肽通过降低关节软骨中缺氧诱导因子(hypoxia-inducible factor,HIF)-2α的水平、调节下游分解代谢基因、降低关节软骨中促炎基因的表达并激活下游合成代谢基因以维持正常骨稳态。南极磷虾肽抑制软骨细胞凋亡的机制有两个,其一是通过阻断关节软骨细胞Fas-FasL信号通路抑制软骨细胞异常凋亡;其二是调节关节软骨细胞中DR3-DR3L凋亡信号通路关键基因的表达[70]。南极磷虾肽有潜力作为改善骨关节炎的功能性食品,并为骨关节炎的临床治疗提供新方向。
南极磷虾蛋白衍生肽的功能活性及其作用机制总结如表4所示。
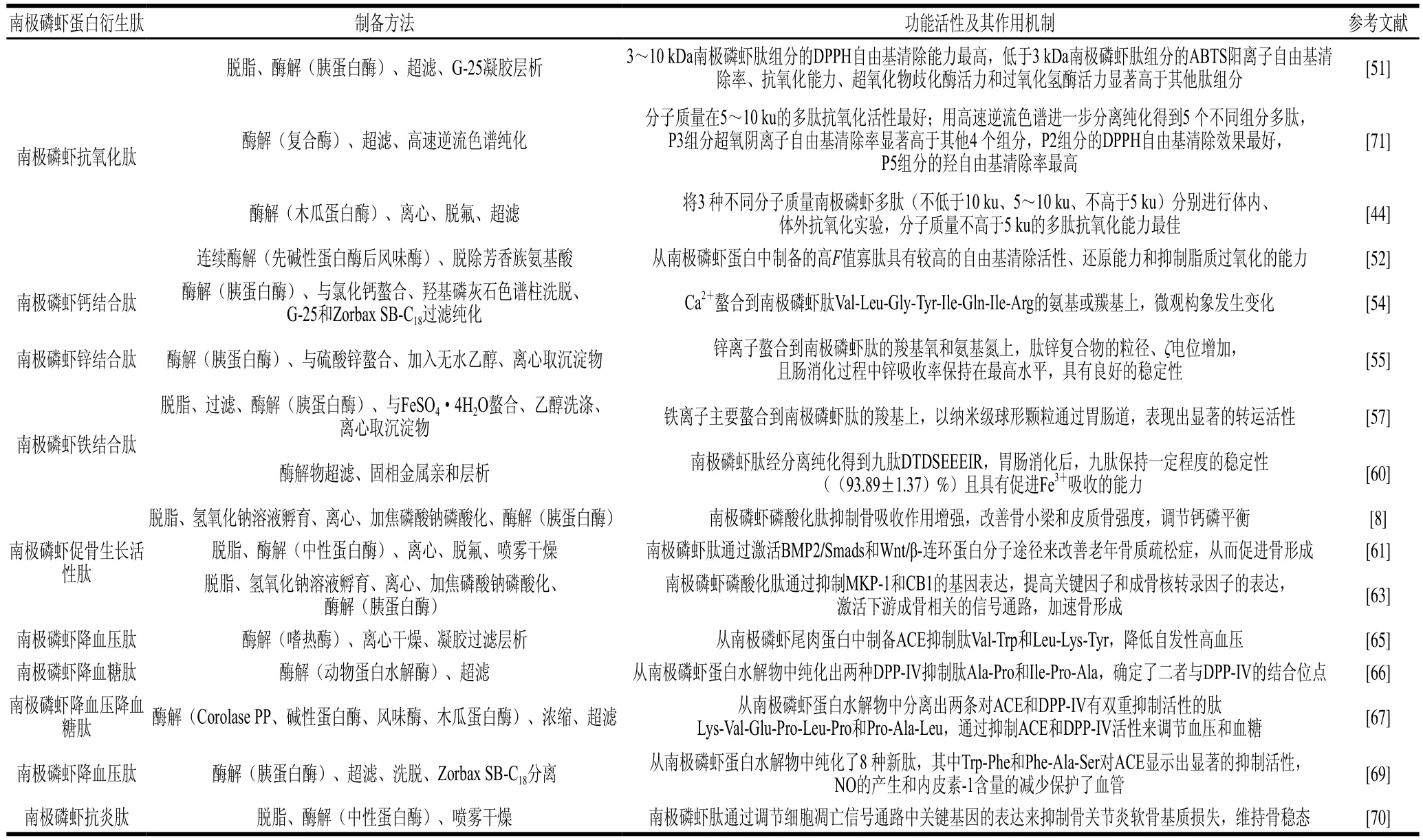
表4 南极磷虾蛋白衍生肽的功能活性及其作用机制Table 4 Functional activities and mechanisms of action of Antarctic krill protein derived peptides
4 南极磷虾蛋白的食用安全性
南极磷虾作为新型食品蛋白质的来源,在食用前必须进行安全性评估。南极磷虾蛋白的安全性问题主要体现在两个方面:一个是含氟问题;另一个是致敏性问题。
4.1 含氟问题
南极磷虾中氟的存在形式主要有水溶态氟、可交换态氟、氧化态氟、有机态氟和残余态氟[72],其中虾肉中残余态氟含量最高,其次为可交换态氟[73]。水溶态氟易被人体吸收利用,可交换态氟虽然也对生物体有较高的有效性,但其在一定条件下可用钙盐除去。南极磷虾中的氟化物主要来自外骨骼、甲壳和头胸部,肌肉中的水平相对较低[74]。南极磷虾整虾的氟含量在1 142~2 400 mg/kg,肌肉氟含量为250 mg/kg,甲壳中含量最高,达到2 566~4 260 mg/kg[75-76]。若捕捞之后及时将南极磷虾的头部和外骨骼与肌肉分离,会有效阻止氟由壳和头胸部向肌肉中转移,降低肌肉中氟的含量。
去除南极磷虾中氟化物的方法正在逐步完善,这为南极磷虾蛋白创造了更广阔的应用前景。Wang Lingzhao等[6]通过在pH 4.6下沉淀磷虾蛋白并用稀盐酸冲洗获得了低氟蛋白提取物,也证实了水洗涤法是一种非常有效的去除南极磷虾蛋白中氟化物的方法。为了节约规模化生产过程中的用水量,Qi Xiangming等[33]使用多级逆流洗涤技术去除了南极磷虾蛋白中98.58%的氟化物。Yin Li’ang等[77]先用食品级柠檬酸[78]、磷酸和盐酸去除氟化物,再用等电点法沉淀蛋白质,获得南极磷虾低氟功能性浓缩蛋白。去氟后的南极磷虾蛋白可作为人类消费的新型食物来源或添加剂,并且使用脱氟技术可大规模生产功能性南极磷虾蛋白产品。
4.2 致敏性问题
据统计学数据显示,虾类过敏是食物过敏发生率占比最高的,在食物过敏人群中有26%的成年人及婴幼儿对虾及其制品过敏,而且亚洲地区虾过敏率高于其他地区[79]。虾类常见过敏原有原肌球蛋白、精氨酸激酶、肌浆钙结合蛋白和肌球蛋白轻链等[80]。其中,原肌球蛋白是具有交叉反应性虾的主要过敏原[81-82],也被认为是南极磷虾的主要过敏原[83]。
Motoyama等[83]研究表明,南极磷虾热提取物存在37 kDa的原肌球蛋白条带,且在免疫印迹中与抗原肌球蛋白的单克隆抗体5G5E1发生反应。南极磷虾原肌球蛋白的氨基酸序列与十足类甲壳动物具有高度序列同源性,说明二者的原肌球蛋白之间具有交叉反应性;另一方面,南极磷虾的原肌球蛋白含量与十足类动物几乎处于同一水平,这表明南极磷虾引起过敏反应的可能性与十足类动物相同。研究显示,南极磷虾原肌球蛋白的序列与太平洋磷虾、龙虾和蟹原肌球蛋白的序列非常相似,序列同源性分别为98%、89%、84%;虾过敏受试者血清的免疫球蛋白(immunoglobulin,Ig)E与南极磷虾、龙虾、蟹的肌肉蛋白提取物中38 kDa条带蛋白反应,且重组南极磷虾原肌球蛋白通过免疫印迹被识别为38 kDa。此外,重组南极磷虾原肌球蛋白对IgE的吸收消除了IgE对重组原肌球蛋白和蛋白质提取物的反应性。综上,南极磷虾原肌球蛋白是一种潜在的、具有交叉反应性的过敏原[82]。
由于食用南极磷虾易导致过敏现象的发生,因此降低或消除南极磷虾过敏原的过敏性显得尤为重要。有研究表明,通过酶解技术[84]、体外模拟胃肠道消化[80]、电子束辐照[85]、美拉德反应[86]、超高压结合酶法[87]等方式可以降低虾类过敏原的致敏性。董晓颖等[84]用5种蛋白酶酶解海白虾,其中水解效果最好的是碱性蛋白酶。蛋白酶能使过敏蛋白的肽链断裂,生成小肽或氨基酸,由此可能对过敏蛋白的致敏性有影响;王学丽[80]发现南美白对虾原肌球蛋白在体外模拟胃液消化过程中会产生一些小分子降解产物,且部分抗原表位被破坏,但是原肌球蛋白在胃液中消化2 h仍具有免疫活性;Guan Aiyan等[85]利用电子束辐照大管鞭虾(Solenocera melantho),通过改变原肌球蛋白的结构来降低其免疫反应性,其中以7 kGy辐照效果最显著;Fu Linglin等[86]采用核糖、低聚半乳糖和壳寡糖进行美拉德反应消除中国对虾原肌球蛋白的致敏性,通过诱导原肌球蛋白由α-螺旋转变为β-折叠导致构象改变,进而降低致敏性;超高压处理主要作用于非共价键,破坏蛋白质空间结构,从而掩盖或破坏过敏原表位,降低致敏性。贾莹等[87]研究发现,采用超高压结合酶法对南美白对虾虾仁致敏性的消减效果好于超高压法。但是目前有关减弱南极磷虾原肌球蛋白致敏性的研究很少,有待进一步研究。
5 结 语
南极磷虾蛋白因储量丰富和较高的营养价值受到广泛关注。南极磷虾蛋白质含量高、必需氨基酸种类齐全,具有良好的起泡性、泡沫稳定性、乳化性、乳化稳定性、吸油性等功能特性,可以应用于泡沫型产品起泡剂、食品乳化剂的开发等;此外,目前已发现南极磷虾蛋白衍生肽具有抗氧化、促矿物质吸收、抗骨质疏松、降血压、降血糖、抗炎等功能活性。作为新型食物蛋白资源,南极磷虾蛋白的含氟问题和致敏性问题也是不容忽视的,尤其是南极磷虾原肌球蛋白致敏性的消减方法还未充分开发,未来还需要在此方面进行更加深入的研究。
尽管南极磷虾蛋白的优势很多、应用前景广阔,但我国对南极磷虾的精深加工和高值化利用起步较晚,南极磷虾蛋白的部分功能特性也限制了其大规模工业化生产,其应用也仅仅停留在用南极磷虾粉替代鱼粉作为饲料,市面上并不常见将南极磷虾蛋白及其衍生肽用作膳食补充剂的功能性产品。所以仍需更多的研究来阐明南极磷虾蛋白的化学组成和功能之间潜在的关系,探索提高其溶解性、凝胶性等功能特性的技术,开发南极磷虾蛋白衍生肽功能性产品,完善降低原肌球蛋白致敏性的方法及机制,这将有助于新型南极磷虾蛋白产品高值化、产业化,并开发针对特定市场、特定功能的各种南极磷虾蛋白产品。
