低鱼粉饲料中添加牛磺酸对许氏平鲉肠道结构、免疫及抗氧化能力的影响
2022-04-29卫力博彭祖想翟浩杰任同军韩雨哲
■卫力博 彭祖想 严 林 高 欣 翟浩杰 王 伟,2 任同军,2 韩雨哲,2*
(1.大连海洋大学水产与生命学院,辽宁 大连 116023;2.辽宁省北方鱼类应用生物学与增养殖重点实验室,辽宁 大连 116023)
许氏平鲉又名黑鲪、黑寨,属近岸底层肉食性鱼类,具有抗病性强、营养丰富和生长快速等优点,是我国北方沿海网箱养殖的重要经济鱼类[1-2]。随着我国水产养殖规模逐渐扩大,许氏平鲉养殖产量逐步上升,对饲料的需求也不断增加,鱼粉作为水产饲料中最重要的蛋白源,由于资源有限,价格上涨等原因已经严重影响了其养殖效益[3]。为了节约饲料成本,通常以成本低廉且资源丰富的植物蛋白源(豆粕、菜粕、花生粕等)进行鱼粉替代,但植物蛋白源除了存在蛋白酶抑制剂、硫代葡萄糖苷等大量内源性抗营养因子外,还缺乏某些在鱼类生长过程中所需的重要氨基酸,如牛磺酸、蛋氨酸等[4-5]。
牛磺酸,化学名称2-氨基乙磺酸,是含硫氨基酸代谢的最终产物,作为动物体含量最丰富的游离氨基酸,具有诱食、促生长、参与胆汁酸调节与渗透调节、提高免疫和抗氧化能力等功能[6-8]。鱼类具有一定的牛磺酸合成能力,但普遍较弱,无法满足自身需求,特别是肉食性鱼类[9]。目前研究已表明,通过在低鱼粉饲料中添加牛磺酸可改善植物蛋白源导致的生长缓慢和绿肝综合征等现象,起到节约鱼粉的效果。Takagiet等[10-11]报道在低鱼粉高植物蛋白饲料中添加牛磺酸,能够显著改善鲉鱼和红鲷的生长性能并降低了绿肝综合征的发病率。刘兴旺等[12]研究表明,在植物蛋白替代鱼粉条件下补充牛磺酸可促进大菱鲆生长,并缓解植物蛋白所导致的脂肪代谢异常现象。目前许氏平鲉饲料研究主要集中在动植物蛋白源替代不同比例鱼粉上[13-15],而关于补充功能性添加剂来提高低鱼粉饲料利用率和缓解低鱼粉饲料所带来的负面效果上的研究甚少[16]。本试验通过在低鱼粉饲料中添加牛磺酸,探究其对许氏平鲉肠道结构、免疫和抗氧化能力的影响,以期为许氏平鲉低鱼粉饲料中牛磺酸的使用提供理论参考。
1 材料与方法
1.1 试验用鱼
试验用许氏平鲉购自大连飞鲲水产养殖有限公司,选取规格均一、健康无伤的平均体质量为(36.25±0.04)g 的许氏平鲉120 尾,暂养1 周以适应实验室养殖条件,期间投喂对照组基础饲料。
1.2 试验饲料
以豆粕、鱼粉为主要蛋白源,鱼油为主要脂肪源,配制牛磺酸水平分别为0(T1 组,对照组)、0.8%(T2 组)、1.6%(T3 组)、2.4%(T4 组)、3.2%(T5 组)的5种等能低鱼粉饲料。牛磺酸(含量99%)购自湖北远大生命科学与技术有限责任公司。甘氨酸的含量随饲料中牛磺酸含量的增加而减少以保证各试验饲料氮的含量一致。所有饲料原料粉碎过60 目筛,原料按照配方进行逐级混匀,期间添加适量的水使其黏合度适宜,经制粒机制成2 mm 颗粒饲料,于40 ℃烘箱中烘至水分为10%左右,晾至室温后存于密封袋中,于-20 ℃冰箱中保存备用,试验饲料组成及营养水平见表1。
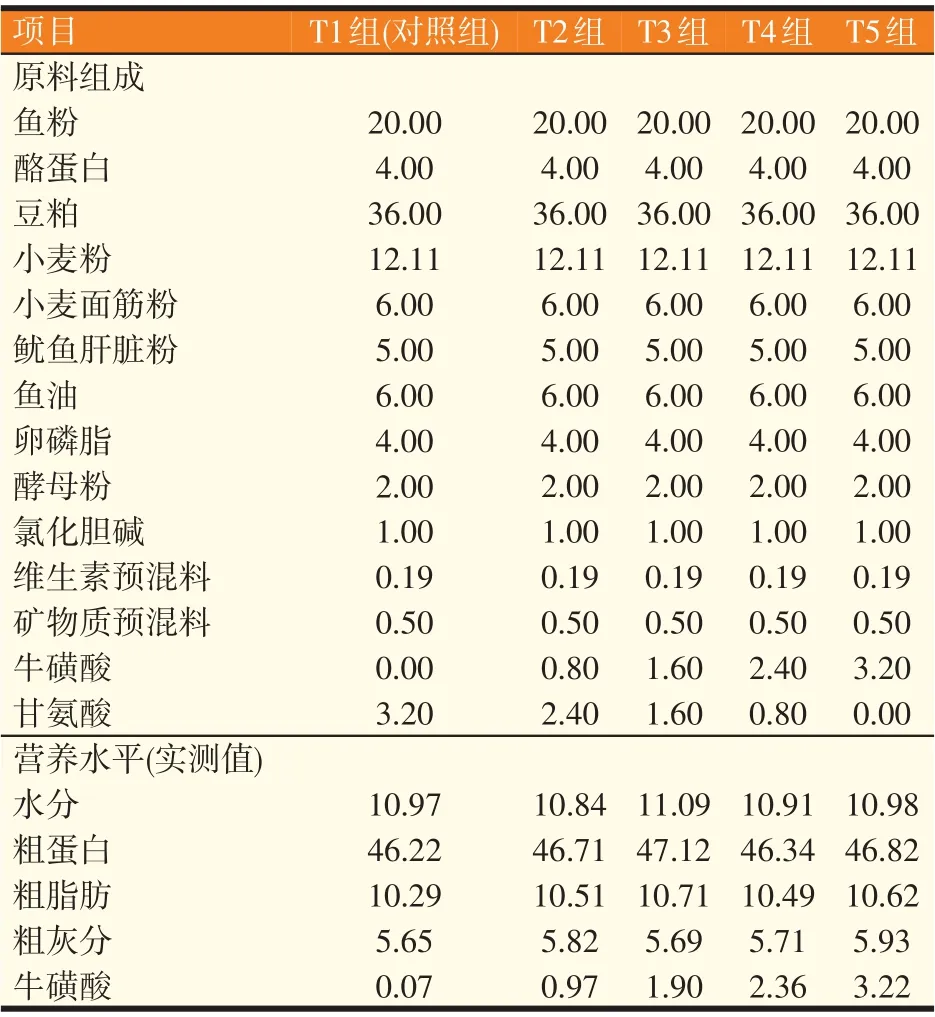
表1 试验饲料组成及营养水平(风干基础,%)
1.3 试验设计与养殖管理
试验开始前,停食24 h,将试验用鱼随机分成5组,每组3个重复,每个重复8尾鱼,共计15个水槽,置于循环水设施中养殖。养殖周期为56 d,每天8:00 和17:00 投喂试验饲料至表观饱食,每日18:30 进行换水,换水量为总水体的1/3。养殖期间,24 h 连续充气,控制水温17~19 ℃,溶解氧浓度大于6 mg/L,pH 7.3~7.8。
1.4 样品采集
试验结束后,所有鱼[平均终末体质量(49.37±1.26)g]进行禁食24 h处理,每个养殖水槽随机选3尾鱼取1 cm 中肠,用4%多聚甲醛固定,用于制作肠道组织切片;剩余试验鱼截取完整前肠和肝脏置于液氮中速冻,后转移至超低温冰箱(-80 ℃)中保存进行酶活性测定。
1.5 酶活性指标测定
肠道消化酶包括淀粉酶(AMS)、脂肪酶(LPS)和蛋白酶(PRO),肝脏免疫酶包括碱性磷酸酶(AKP)、酸性磷酸酶(ACP)和溶菌酶(LZM),肝脏抗氧化指标包括过氧化氢酶(CAT)、总超氧化物歧化酶(T-SOD)和丙二醛(MDA),上述指标测定试剂盒均购自南京建成生物工程研究所有限公司,并参照说明书进行测定。
1.6 肠道组织形态观察
肠道样品经乙醇脱水后置于二甲苯中透明,随后进行石蜡包埋处理,使用切片机横向间隔切片,而后进行苏木精-伊红(HE)染色,中性树胶封片。使用光学显微镜拍照后用图像软件S-Viewer 进行观察测量。
1.7 数据处理
试验数据以“平均值±标准误(Mean±SE)”表示。使用SPSS 21.0软件对试验数据进行单因素方差分析(one-way ANOVA),采用Duncan’s 法进行组间差异分析,设置差异显著为P<0.05。
2 结果与分析
2.1 低鱼粉饲料中添加牛磺酸对许氏平鲉肠道消化酶活性的影响(见表2)
由表2可知,T2、T3和T4组淀粉酶和脂肪酶均显著高于对照组(T1组)(P<0.05),T5组与对照组相比无显著差异(P>0.05)。T2组肠道蛋白酶活性显著高于其他各组(P<0.05),之后随着牛磺酸添加量的增加,蛋白酶活性有所下降。

表2 低鱼粉饲料中添加牛磺酸对许氏平鲉肠道消化酶活性的影响(n=3,U/mg prot.)
2.2 低鱼粉饲料中添加牛磺酸对许氏平鲉肠道结构的影响(见图1、表3)
由图1可知,对照组许氏平鲉中肠皱襞高度较低,皱襞破损严重,而牛磺酸添加组许氏平鲉肠道皱襞无明显损伤脱落情况,皱襞整齐排列,由表3 可知,T2、T3组皱襞高度和皱襞宽度显著高于对照组(P<0.05)。
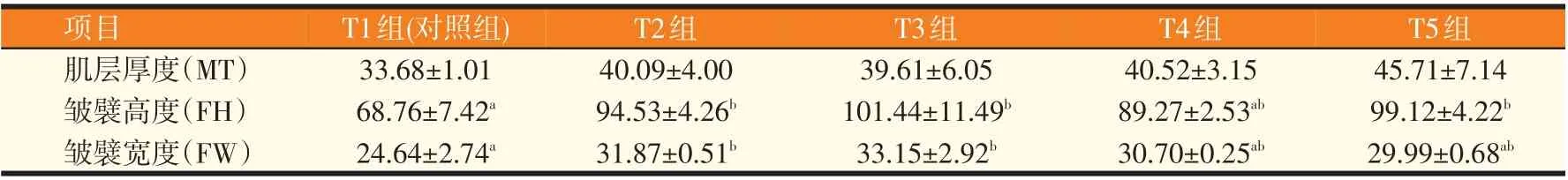
表3 低鱼粉饲料中添加牛磺酸对许氏平鲉肠道形态的影响(n=3,μm)
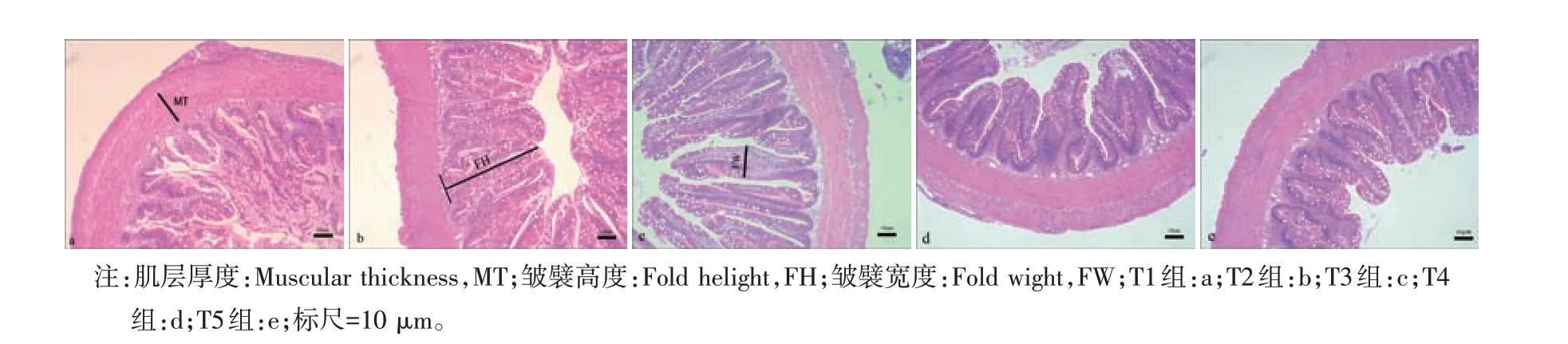
图1 许氏平鲉肠道组织结构
2.3 低鱼粉饲料中添加牛磺酸对许氏平鲉肝脏免疫指标的影响(见表4)
由表4可知,与对照组相比,T2和T3组碱性磷酸酶、溶菌酶活性显著提高(P<0.05);各牛磺酸添加组的酸性磷酸酶活性显著高于对照组(P<0.05),且随着牛磺酸含量的升高有下降趋势,但差异不显著(P>0.05)。

表4 低鱼粉饲料中添加牛磺酸对许氏平鲉肝脏免疫指标的影响(n=3)
2.4 低鱼粉饲料中添加牛磺酸对许氏平鲉肝脏抗氧化能力的影响(见表5)
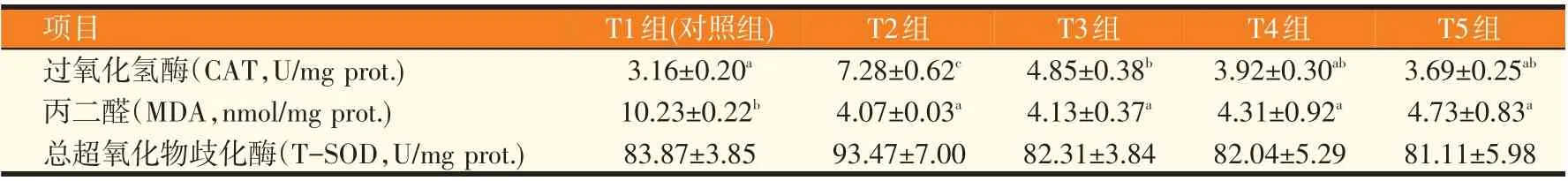
表5 低鱼粉饲料中添加牛磺酸对许氏平鲉肝脏抗氧化能力的影响(n=3)
由表5可知,T2组过氧化氢酶活性显著高于其他各组(P<0.05),之后出现下降趋势。牛磺酸添加组丙二醛含量相较于对照组显著下降(P<0.05),且随着牛磺酸添加量的增加有上升趋势。各组间总超氧化物歧化酶活性无显著差异(P>0.05)。
3 讨论
3.1 低鱼粉饲料中添加牛磺酸对许氏平鲉肠道健康的影响
鱼类肠道消化酶是反映鱼体消化生理机能的重要指标,可以直接影响鱼体对营养物质的吸收和利用情况[17]。试验结果表明在低鱼粉饲料中添加适宜牛磺酸可增强许氏平鲉肠道脂肪酶、淀粉酶和蛋白酶活性,提升鱼体对营养物质的消化吸收。研究表明牛磺酸在鱼体内会与胆酸、鹅脱氧胆酸等游离胆酸合成牛磺胆酸和牛磺鹅脱氧胆酸,结合胆汁酸的增加进而会激活鱼体内胆盐刺激脂酶来分解脂肪[18-20],对脂肪酶活性具有促进作用;淀粉酶活性的提升一方面与牛磺酸提升机体α-淀粉酶基因的相对表达量有关[21],另一方面牛磺酸会刺激胰岛素的释放来血糖降低,从而间接促使淀粉酶活性升高进行葡萄糖分解;此外适量牛磺酸还会刺激鱼体蛋白酶分泌,促进蛋白质的消化吸收[22]。
肠道皱襞高度、宽度以及肌层厚度会直接影响肠道的完整性和稳固性,对鱼体健康极为重要。低鱼粉高植物蛋白饲料因存在大量抗营养因子和适口性差等原因会对鱼类肠道造成损伤,而本试验结果显示通过添加牛磺酸可以明显修复许氏平鲉肠道的损伤状态,提升肠道皱襞高度及宽度。据相关报道牛磺酸不仅作为结肠上皮细胞中含量最丰富的游离氨基酸,与上皮细胞增殖和肠道结构的变化相关[23],还会通过调节TLRs/NF-κB 信号通路,下调肠道促炎因子与上调抗炎因子的表达,缓解高植物蛋白引起的肠道炎症,保持肠道健康生长[24]。但随着牛磺酸添加量的上升,许氏平鲉肠道消化酶活性、皱襞高度及宽度有下降趋势,在青鱼[25]、花鳗鲡[26]的研究上也得到了类似结果,其原因可能是鱼类摄入过量牛磺酸会导致肠道处于高酸度环境,抑制肠道拟杆菌门等一些参与营养代谢的优势菌群生长[27]和肠道α-淀粉酶基因的相对表达[21],进而对肠道健康造成严重损伤。
3.2 低鱼粉饲料中添加牛磺酸对许氏平鲉免疫水平的影响
牛磺酸作为机体内淋巴细胞和吞噬细胞中含量最丰富的游离氨基酸,具有促进巨噬细胞产生白介素、促进免疫球蛋白分泌和增强中性粒细胞吞噬杀菌能力等多种生物学功能[28]。溶菌酶可水解致病菌细胞壁的肽聚糖,致其内容物流出,加速细菌溶解[29];碱性磷酸酶存在机体各组织中,可通过改变细菌表面结构,加速吞噬细胞对其进行吞噬降解[30];酸性磷酸酶是巨噬细胞溶酶体的标志酶,而溶酶体通过水解致病菌来达到保护机体的作用[31]。本试验中,饲料中添加0.8%牛磺酸会显著增强许氏平鲉肝脏溶菌酶、碱性磷酸酶和酸性磷酸酶的活性,表明适量的牛磺酸对增强机体免疫具有重要作用,此外Zhang 等[32]在探究牛磺酸对黄颡鱼急性氨中毒的缓解作用时也有相同的发现,牛磺酸可以有效遏制试验鱼体内溶菌酶活性和细胞吞噬指数下降的现象。但当牛磺酸添加量在1.6%~3.6%时,许氏平鲉肝脏免疫酶活性出现下降趋势,这一结果与青鱼[25]、草鱼[33]、红鳍东方鲀[34]的研究上相同,说明过多的牛磺酸会对鱼体免疫应答产生抑制作用,相关研究表明过量的牛磺酸会上调鱼类的促炎细胞因子,肝脏健康程度下降,导致机体免疫酶活性降低[35]。此外肠道健康与鱼体健康水平息息相关,过多的牛磺酸摄入时肠道受损,同样会使机体的免疫水平下降[36]。
3.3 低鱼粉饲料中添加牛磺酸对许氏平鲉抗氧化能力的影响
相关研究表明,牛磺酸会与机体内次氯酸结合生成牛磺氯胺,而牛磺氯胺会激活Nrf2/ARE 信号通路,从而上调抗氧化酶的相对基因表达[37],此外牛磺酸作为线粒体蛋白质合成的调节器,具有增强电子传递链活性并保护线粒体免受过量超氧化物影响的作用,进而提升机体的抗氧化能力[38]。本试验研究中,饲料中添加牛磺酸提高了过氧化氢酶活性并显著降低了丙二醛的含量,表明牛磺酸促进了鱼体对氧自由基的清除能力,降低了脂质过氧化物的产生,缓解了许氏平鲉摄食高植物蛋白饲料后机体的氧化损伤程度。Rosemberg 等[39]在探究牛磺酸对乙醇急性诱导斑马鱼大脑氧化应激后的保护效果时也有相同发现,150 mg/L 和400 mg/L 牛磺酸可有效抑制脂质过氧化并提升过氧化氢酶、超氧化物歧化酶的活性。此外本试验中当牛磺酸添加量超过0.8%时过氧化氢酶活性有降低趋势,丙二醛含量有上升趋势,表明适宜的牛磺酸含量对机体抗氧化能力具有提升作用,而含量过高可能会产生抑制作用,其原因可能是过高牛磺酸水平会使鱼类肝脏出现肝窦扩张现象,加速肝细胞与血液之间的物质交换,降低血清中白蛋白、球蛋白含量,打破了肝脏代谢平衡,使肝脏健康受损,机体的抗氧化能力下降[40]。
4 结论
本试验通过在低鱼粉饲料中添加牛磺酸饲养许氏平鲉56 d后发现,适量牛磺酸可提升许氏平鲉肠道脂肪酶、蛋白酶和淀粉酶活性并改善其肠道结构,提高肝脏免疫及抗氧化能力,但过量牛磺酸会对鱼体产生不利的影响,根据肠道健康指标,建议许氏平鲉低鱼粉饲料牛磺酸最适添加范围为0.8%~1.6%。
