卵形鲳鲹生肌调节因子基因家族的鉴定及在胚胎中的表达
2022-04-27余艳玲罗洪林冯鹏霏潘传燕宋漫玲张永德
余艳玲,罗洪林,罗 辉,冯鹏霏,潘传燕,宋漫玲,肖 蕊,张永德,*
(1.广西水产科学研究院 广西水产遗传育种与健康养殖重点实验室,广西 南宁 530021; 2.西南大学 动物科学学院,重庆 400715)
脊椎动物的运动系统由骨骼与肌肉组成,骨骼肌在产生运动、支撑骨骼、维持姿势、保护身体和产热等方面发挥着至关重要的作用。骨骼肌的生长发育是一个由多种转录因子参与调控的复杂过程,在此过程中,生肌调节因子(myogenic regulatory factors, MRF)在骨骼肌的发育及调控中发挥至关重要的作用。MRF被认为是调节肌肉发育分化的分子开关,其功能与活性可以决定整个细胞谱系的命运。MRF家族由4种肌肉特异性蛋白质组成,包括生肌决定因子(myogenic differentiation,)、生肌调节因子6(myogenic regulatory factor 6,6)(或称4)、生肌调节因子5(5)和肌细胞生成素(myogenin,)。它们在肌肉发育谱系的多个点起作用,通过调节前体细胞的增殖、不可逆转的细胞周期停滞来协同建立骨骼肌表型,然后调节肌节和肌肉特异基因的激活,以促进分化和肌节的组装。在发育过程中,5和被视为成肌决定因素,控制胚胎中肌细胞的确定和分化,以及肌细胞的生长发育和再生,Myf6与MyoG是肌源性分化因子,在Myf5和MyoD的下游发挥作用,它们有助于成肌细胞的分化,而Myf6在决定和分化水平上都发挥作用。
MRF家族蛋白质主要包含3个结构域,第一个是与螺旋-环-螺旋结构域(helix-loop-helix, HLH)相连的基本域,合称为bHLH,第二个是半胱氨酸/组氨酸结构域,位于靠近bHLH的N端,第三个是富含丝氨酸/苏氨酸的结构域,位于C末端附近,而bHLH结构域被认为是肌生成激活的主要贡献者。HLH蛋白质有7类,MRF被认为是Ⅱ类蛋白质,因为其表达仅限于单一组织类型(骨骼肌)。bHLH结构域可以结合DNA中称为E-box(CANNTG)的特定序列,与E-box的结合受到表观遗传的调控。HLH结构域允许与普遍存在的Ⅰ类bHLH E蛋白质(包括E12、E47、HEB/HTF4和E2-2/ITF-2)进行同二聚或异二聚,从而进行肌生成。
骨骼肌的基本结构是在胚胎发生过程中形成的。尽管MRF基因在胚胎肌发生过程中仅在肌细胞中表达,但在蛋白质水平以及表达的时间和阶段上存在内在差异,反映了MRF在肌细胞分化中的潜在特异性。这些差异突出了每个MRF基因在蛋白质水平上对信号传导及其相互作用伙伴的特异性反应,迄今为止,这方面的研究还十分薄弱。
卵形鲳鲹()是我国海水养殖的名贵鱼类,也是适合我国近、远海集约化养殖的重要品种之一,近些年在我国南部沿海地区发展迅速。随着卵形鲳鲹产业的发展,种质退化现象逐渐显露,主要表现为生长速率下降、群体整齐度低、畸形、抗病力低、病害发生频繁、鱼苗死亡率高等。因此,开展卵形鲳鲹生长相关基因的研究,对其产业的健康持续发展意义重大。MRF家族基因是动物肌肉发育中的重要调控因子,了解MRF基因在胚胎发育中的表达,对了解卵形鲳鲹肌肉发育特征及生长发育具有重要意义。然而,目前对卵形鲳鲹MRF基因家族尚不了解,其在胚胎发育过程中发挥的作用及其表达模式尚不清楚。本研究基于全基因组序列,对卵形鲳鲹的MRF基因家族进行了鉴定,并对其在胚胎发育各阶段中的表达情况进行了检测,旨在为进一步研究MRF家族基因在卵形鲳鲹胚胎发育过程中的作用及其肌肉组织的发生发展等奠定基础。
1 材料与方法
1.1 材料
卵形鲳鲹受精卵取自深圳南澳镇大鹏湾海域人工养殖群体,待其产卵与自然受精后,即刻捞起受精卵,置于1 000 L的孵化桶孵化,孵化温度为26~31 ℃。显微镜下观察确定胚胎所处的发育阶段,对受精卵(OSP)、8细胞期(8-Cell)、16细胞期(16-Cell)、64细胞期(64-Cell)、高囊胚期(HBS)、原肠早期(EGS)、原肠中期(MGS)、原肠末期(LGS)、胚体形成期(EFS)、眼囊期(OPVS)、耳囊期(OTVS)、心脏跳动期(HPS)、晶体出现期(FELS)共13个发育阶段的胚胎分别取样,每个阶段取样品3份,液氮保存。
RNAsimple提取试剂盒购自天根生物科技(北京)有限公司;HiScriptⅢ RT SuperMix、ChamQ SYBR qPCR Mix、AceTaqMaster Mix均购自南京诺唯赞生物科技有限公司。
1.2 MRF基因家族的鉴定与理化特性分析
卵形鲳鲹基因组由本单位测序组装获得(PRJNA574895)。MRF基因家族采用两种方法鉴定,一种是从NCBI数据库获取了鲹形目MRF家族30个蛋白质序列,这些序列被用作查询序列以进行BLASTp搜索(E值<10),使用blastp程序对卵形鲳鲹所有蛋白质序列进行同源分析。然后,手动去除重复的基因,并获得原始候选基因。另一种方法是从Pfam数据库(http://pfam.xfam.org/)下载MRF基因家族序列特征文件(PF01586),采用hmmer 3.1软件,根据序列特征文件PF01586识别卵形鲳鲹MRF家族基因的蛋白质序列,其中e值小于-20。获得的候选序列通过SMART(http://smart.embl.de/)、NCBI CDD(https://www.ncbi.nlm.nih.gov/cdd/)与pfam数据库手动确认结构域,确定候选蛋白质序列。采用Protparam(https://web.expasy.org/protparam/)对MRF家族蛋白质的分子量、等电点和疏水性均值(GRAVY)等理化特性进行分析;采用SWISS-MODEL(https://swissmodel.expasy.org/)对蛋白质三级结构进行预测分析。
1.3 基因结构、motif预测及染色体定位分析
通过基因组注释文件获得MRF家族各基因的5′-UTR、3′-UTR、外显子、内含子位置及长度等信息,采用GSDS 2.0(http://gsds.cbi.pku.edu.cn/)绘制基因结构示意图,利用MEME 5.1.1(http://meme-suite.org/tools/meme)在线工具对MRF家族基因的基序(motif)进行预测分析,基序的最短长度设为10,最大长度为100,利用TBtools 0.66软件作图。通过基因组注释文件提取基因在染色体中的位置,采用Samtools 1.10软件分析获得各染色体的长度,Mapchart 2.32软件进行染色体绘图。
1.4 序列比较及系统进化树的构建
采用Mega X软件的ClustalW程序对卵形鲳鲹MRF家族基因全长氨基酸序列进行比对,计算基因间的遗传距离,并应用邻接法(NJ)构建系统聚类树,选择Poisson模型,采用“bootstrap”1 000次自举检测表明各分支的置信度。从Ensembl(http://asia.ensembl.org/index.html)数据库下载高体鰤()、黄尾鰤()、大菱鲆()、半滑舌鳎()4个物种的基因组序列文件,采用与卵形鲳鲹同样的方法进行基因家族鉴定,然后采用最大似然(ML)法对高体鰤、黄尾鰤、大菱鲆、半滑舌鳎与卵形鲳鲹5种鱼类的MRF家族基因编码的氨基酸序列进行比对,构建ML系统聚类树,采用Poisson模型,对空位采用完全删除(complete deletion),并用“bootstrap”1 000次自举检测表明各分支的置信度。
1.5 RNA分离与qRT-PCR分析
通过天根生物RNAsimple提取试剂盒分离卵形鲳鲹不同发育阶段的胚胎组织总RNA。通过2%琼脂糖凝胶电泳检测RNA的完整度,采用NanoDrop One检测RNA的浓度。对完整性较好的RNA,根据南京诺唯赞Hiscript Ⅲ反转录试剂盒对提取的总RNA进行反转录,合成cDNA第一链。以18S rRNA作为内参基因,采用诺唯赞2×ChamQ SYBR qPCR Master Mix进行实时荧光定量PCR(qRT-PCR)分析。15 μL反应体系中包含ChamQ Universal SYBR qPCR Mix 7.5 μL,上下游引物(10 μmol·L)(表1)各0.3 μL,cDNA模板1.2 μL,ddHO 5.7 μL。每个样品进行3次生物学重复,每种MRF基因的PCR产物均通过测序以确认序列的准确性。采用SPSS 19.0软件进行单因素方差分析(ANOVA),通过Duncan多重比较分析各组织间的差异显著性。

表1 卵形鲳鲹MRF家族基因qRT-PCR引物信息
2 结果与分析
2.1 MRF基因家族的鉴定与理化特性分析
通过对卵形鲳鲹MRF基因家族的鉴定,共获得了5个基因。经基因注释分析,确认5个基因分别为1、2、5、6与。根据氨基酸序列,对MRF家族蛋白质的理化特性进行了预测分析(表2),卵形鲳鲹MRF家族蛋白质长度介于231~297 aa,蛋白质相对分子质量介于25 542.27~32 206.38 u,其中ToMyf6与ToMyoD1分别对应最小和最大的蛋白质。所有MRF蛋白质的等电点在5.33~6.33,其中ToMyoD1与ToMyoG蛋白质分别具有最小和最大的等电点,5个蛋白质均为酸性蛋白质。GRAVY的值介于-0.591~-0.849,表明这些蛋白质均为亲水性蛋白。5个蛋白质的不稳定性指数介于65.35~89.12,表明这些蛋白质均属于不稳定蛋白质。三级结构预测结果显示(图1),卵形鲳鲹MRF家族蛋白质均呈现典型的螺旋-环-螺旋结构,两螺旋结构一短一长,呈约140°的夹角。
2.2 基因结构及motif预测分析
对卵形鲳鲹MRF家族5个基因的结构分析发现(图2),5个基因都由外显子、内含子和 UTR 区组成,但序列信息中5与6缺少5′-UTR,1缺少3′-UTR。除6可能包含4个外显子与3个内含子外;其余4个成员均包含3个外显子和2个内含子。
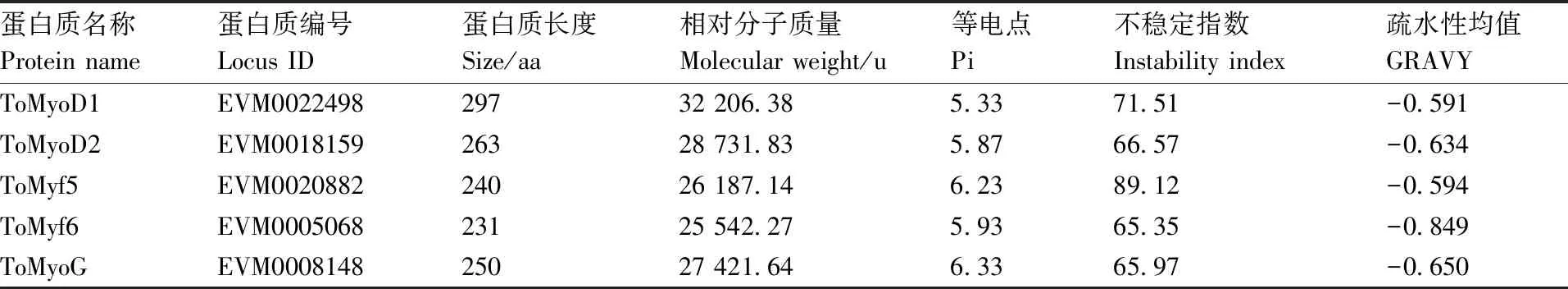
表2 卵形鲳鲹MRF家族蛋白质的理化特性
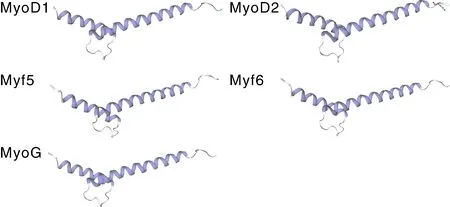
图1 卵形鲳鲹MRF家族蛋白质的三级结构Fig.1 Tertiary structure of MRF family proteins in T. ovatus
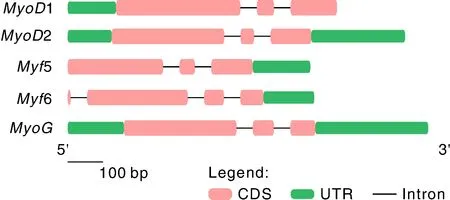
图2 卵形鲳鲹MRF家族基因的结构Fig.2 Gene structure of the T. ovatus MYF family genes
采用SMART、NCBI CDD与pfam三大数据库对卵形鲳鲹MRF家族5个基因编码的蛋白质进行结构域确认,结果显示,5个蛋白质均存在BASIC结构域和HLH结构域(图3-a),其中HLH结构域较为保守,而MYF5结构域仅在1、2与5中保守,而在6与中却不保守。为了进一步鉴定其保守 结构域,采用MEME工具分析了MRF家族5个蛋白质的氨基酸序列,总共获得了6个保守的基序(motif)(图3-a、b),每个基因包含3~5个基序。其中基序1包含该家族基因典型的HLH结构域,与基序4一样均在5个基因中出现,基序2、基序3仅在1与2中出现,基因缺少基序5,基序6仅在5与中出现。
2.3 MRF家族基因的染色体定位分析
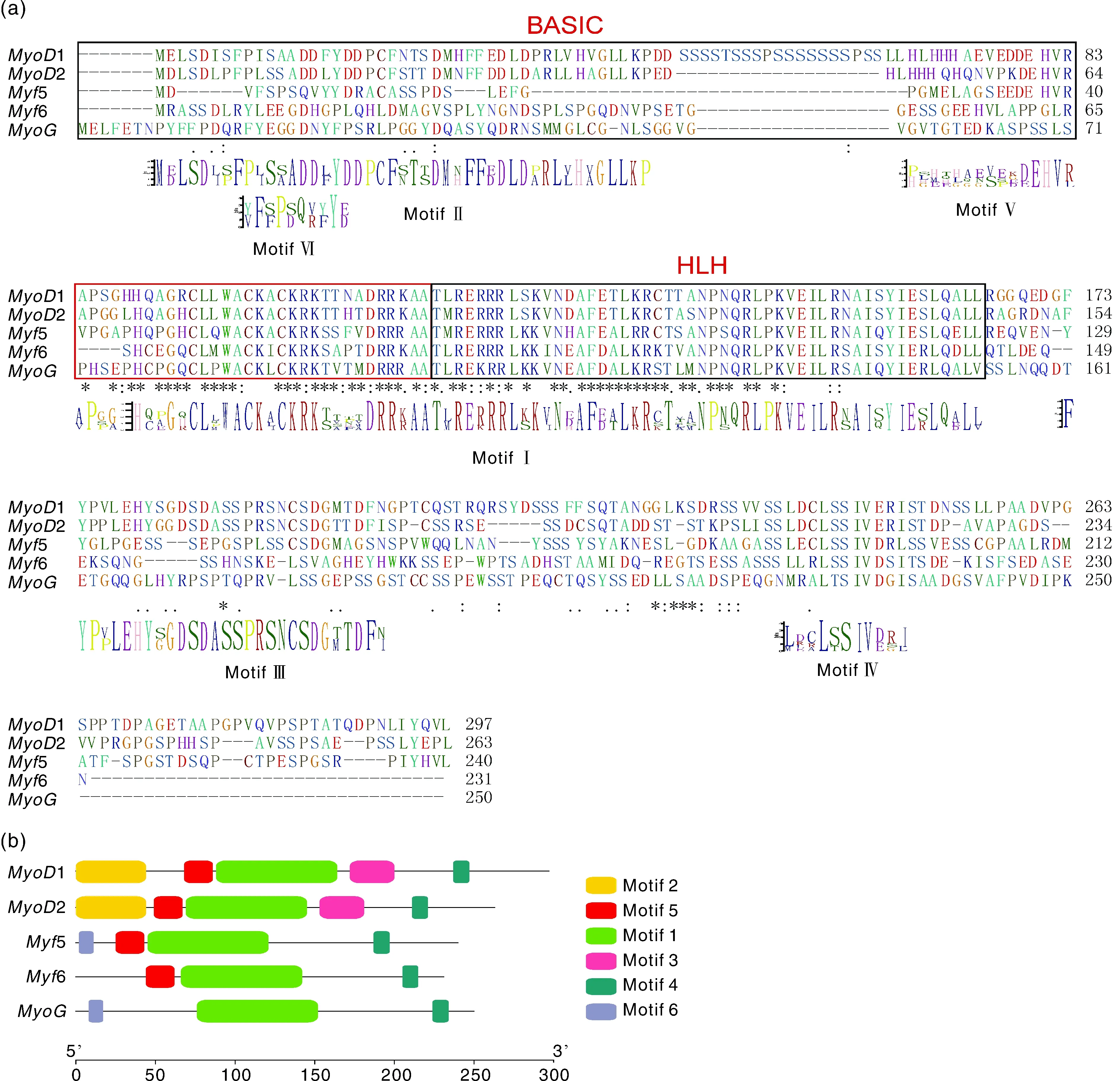
a,MRF家族成员的结构域及保守基序位置;b,MRF家族成员的保守基序组成。a, Domain and conserved motif position of MRF family genes; b, Conserved motif composition of MRF family genes.图3 卵形鲳鲹MRF基因家族的结构域及保守基序Fig.3 Domain and conserved motif of MRF gene family in T.ovatus
染色体定位分析显示,MRF家族基因定位于卵形鲳鲹4对染色体上(图4),其中2基因定位于1号染色体,基因位于2号染色体,1基因位于9号染色体,而5与6基因则均位于22号染色体,两基因位置相连,中间未检测到其他基因的存在。
2.4 序列比较及系统进化树的构建
对卵形鲳鲹MRF家族5个基因进行序列比对,构建NJ聚类树(图5),结果显示,5个基因的同源性在38.89%~68.34%,其中1与2基因的同源性最高,为68.34%,其遗传距离也最近,为0.351 8。而与1、6
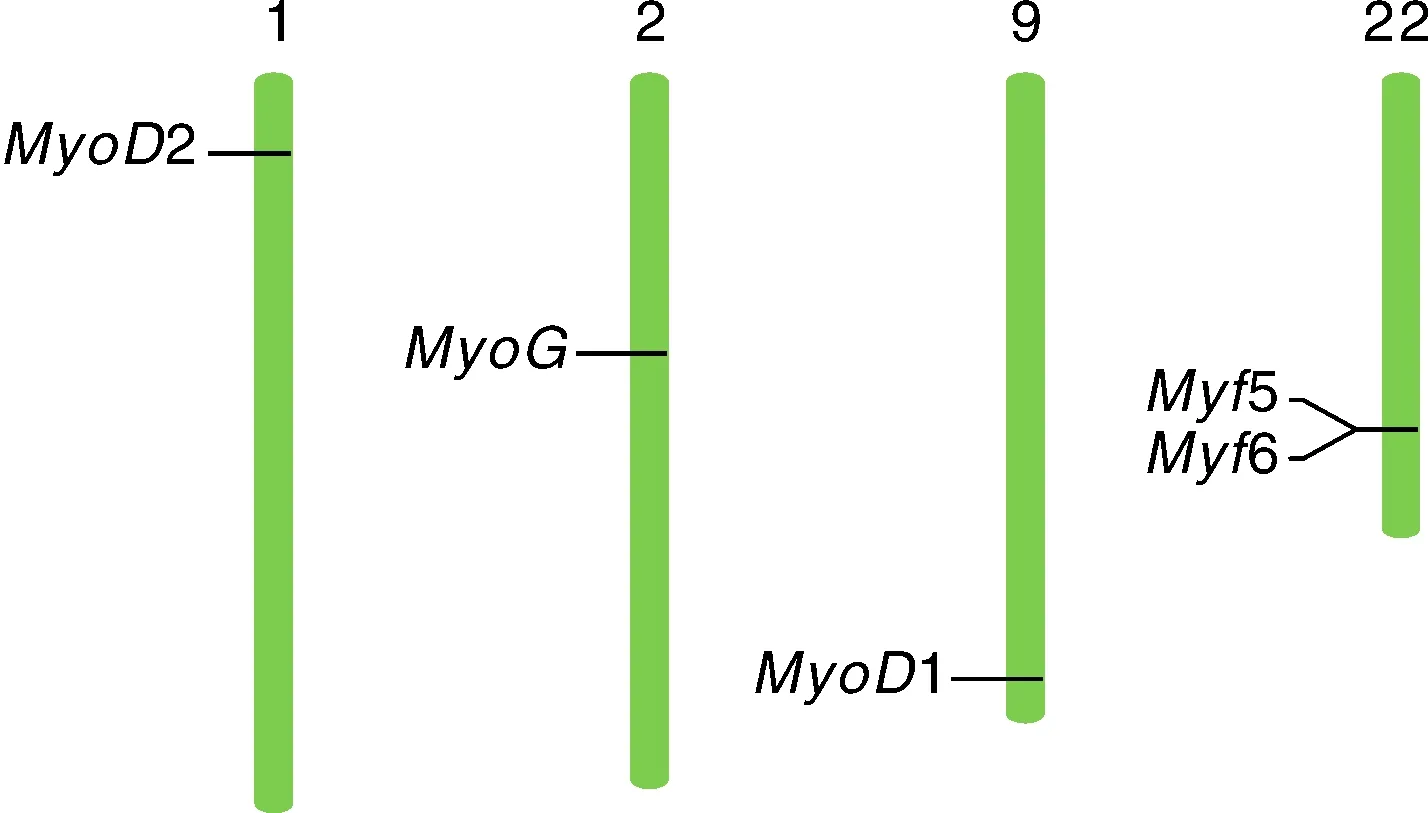
图4 MRF家族基因在卵形鲳鲹染色体上的分布Fig.4 Chromosome distribution of MRF family genes in T. ovatus
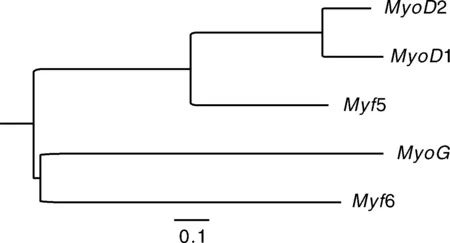
图5 卵形鲳鲹MRF家族基因NJ聚类树Fig.5 MRF family gene NJ clustering tree of T. ovatus
两个基因的同源性最小,均为38.89%,与1之间遗传距离最大,为1.032 2。聚类树显示,1、2与5聚为一支,而6与聚为另一支。
对高体鰤、黄尾鰤、大菱鲆与半滑舌鳎4种鱼类MRF家族基因进行鉴定,共获得20个基因,每种鱼类5个基因,经基因注释分析,确认5个基因分别为1、2、5、6、(表3)。将卵形鲳鲹与上述4种鱼类MRF家族基因编码的氨基酸序列进行比对分析,构建ML系统发育树。25个基因共聚为3个大分支(图6),其中1与2基因聚为一支,5独立成为一支,两分支具有较近的遗传距离,而6与聚为一支。
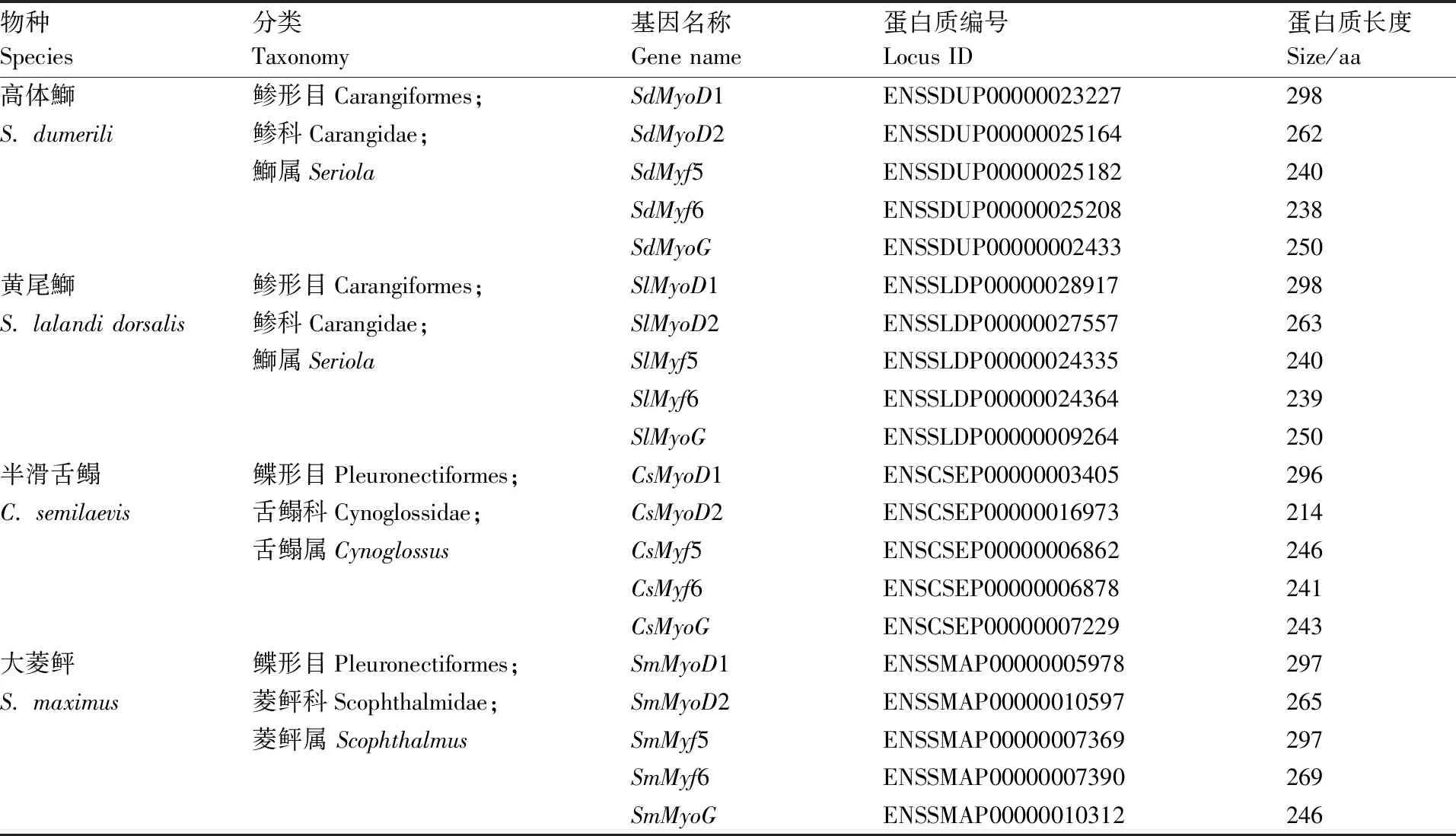
表3 四种鱼类MRF家族基因信息
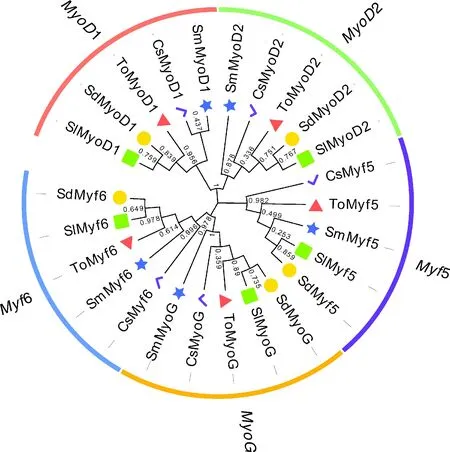
图6 5种鱼类MRF家族基因编码的蛋白质的ML系统进化树Fig.6 Phylogenetic tree of MRF family proteins in five fish species
2.5 不同胚胎发育时期MRF家族基因的表达
利用qRT-PCR研究卵形鲳鲹MRF家族基因在13个胚胎发育时期的组织表达情况,经基因测序确认所分析的基因即为MRF家族基因。其中1、2、5与6基因在卵形鲳鲹胚胎发育过程中的表达模式较相似,从受精卵到原肠中期的7个时期表达量较低,而从原肠末期到孵化期的表达量迅速升高,且均显著高于前7个时期(<0.05)。的表达量在卵形鲳鲹胚胎发育过程中呈现先升高后降低的趋势,在受精卵到胚体形成期的前9个时期表达量极低,而在眼囊期、耳囊期与心脏跳动期3个时期表达量迅速升高,到晶体出现期其表达量又有所降低,但仍然高于前9个胚胎发育期(<0.05)(图7)。
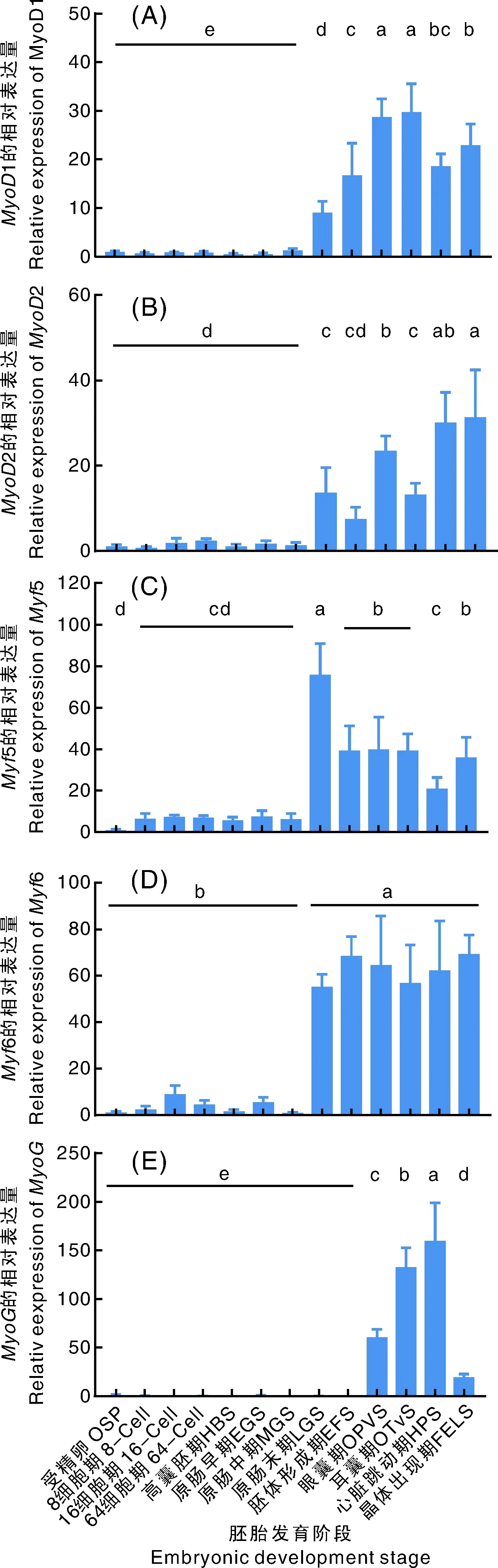
没有相同小写字母表示差异显著(P<0.05)。The bars without the same letters meant the significant difference (P<0.05).图7 MRF家族基因在卵形鲳鲹不同胚胎发育阶段的表达Fig.7 Expression pattern of MRF family genes in different developmental stages of T. ovatus embryos
3 讨论
脊椎动物的肢体和躯干肌肉源自最早在体节中建立的成肌祖细胞,体节的背侧域细胞发育成为生皮肌节,生皮肌节最后发育成真皮、骨骼肌以及血管平滑肌、内皮细胞等。所有这些事件主要受MRF即、5、6和的控制,此过程在所有脊椎动物中是高度保守的,其结构也存在高度的相似性;但是鱼类的肌肉生长还具有某些明显的特征,如生长不定、生长速率和达到的最大尺寸存在种属差异等。此外,鱼类MRF的表达容易受温度等环境变量和鱼类的生长阶段及性别等生物学特性的调节,饲料中高浓度的大豆浓缩蛋白质替代水平或饥饿也会改变生肌调节因子家族的时空表达模式。因此,了解与肌发生相关的MRF的分子结构和功能对于深入了解鱼类肌肉发育的特征是必要的,并对提高鱼类肌肉的生长、肉质和总产量方面具有重要意义。本研究采用Blast与hmm模型两种方法分别对卵形鲳鲹、高体鰤鱼、黄尾鰤鱼、大菱鲆与半滑舌鳎5种鱼类MRF家族基因进行了鉴定,各鉴定出5个MRF家族基因,分别为1、2、5、6和,说明在这些鱼类中MRF成员的数量是高度保守的。然而人类与小鼠等高等动物MRF家族基因成员中可能缺少2基因。在这些物种的进化过程中,2基因可能发生了丢失,或被其他基因所替代。
一般相似度较高的序列,可能发挥相似的蛋白质功能。对卵形鲳鲹5个基因的结构分析结果显示,除6基因可能为4个外显子,其余4个基因均由3个外显子和2个内含子组成,基因组成高度相似。结构域分析显示,MRF家族基因编码的蛋白质存在3个结构域:BASIC结构域、HLH结构域和MYF5结构域。在所有的MRF基因编码的蛋白质中,均包含BASIC结构域和HLH结构域,说明它们在进化上存在较高的保守性。其中HLH结构域是最保守的结构域,它定义了MRF家族,该结构域的氨基酸序列在5个MRF基因之间同源性最高,序列间差异最小;但是MYF5结构域仅存在于1、2与5中,而6与却不含有该结构域。MRF家族基因成员之间结构的差异,说明其功能虽有相似性,但也存在差异,这种多样性推动了多基因家族的进化,各基因的具体功能差异有待进一步研究。卵形鲳鲹MRF家族基因结构与其他脊椎动物存在高度的相似性,说明MRF家族基因在脊椎动物的进化过程中是高度保守的,其功能也可能是高度保守的。
卵形鲳鲹具有24对染色体,对卵形鲳鲹MRF基因在染色体上的定位分析表明,MRF家族基因定位于1号、2号、9号与22号染色体上,其中5与6均定位于22号染色体,且位于同一基因座,形成5-6簇,且具有广泛重叠的共有调控元件,说明这两个基因可能具有密切的进化关系。1与2基因序列具有高度的相似性,其基因的同源性为68.34%,暗示其在进化中存在较密切的关系,基因功能也可能高度相似,但2个基因却位于不同的染色体上(9号与1号染色体)。因此,1与2可能是由独立的复制产生的,或可能在进化的过程中整个基因组发生复制产生,但在某些物种内2基因发生了丢失,或被其他基因所替代。
对5种鱼类MRF家族成员的序列比对及聚类分析结果发现,25个MRF家族成员聚为3大分支,其中1与2基因聚为一支,5独立成为一个分支,两分支遗传距离较近,而6与聚为第三分支。这些进化分支具有较高的邻近匹配性,进化支内的关系也被发现具有很高的匹配率。该结果表明,每个亚家族中的MRF基因可能经历了相同的进化事件并具有相似的功能。在脊椎动物中,MRF家族基因被分为两个主要功能组:和5参与肌肉细胞的决定,发挥“早期”的功能,和6主要参与分化,一种被“早期”MRF激活的“晚期”功能,而与5较近的遗传距离,以及6与较近的遗传距离,均与这些基因的功能分类相似。系统发育分析还表明,高体鰤与黄尾鰤MRF家族5个基因具有最高的序列同源性,遗传距离最近,这两种鱼均为鰤属()鱼类。卵形鲳鲹与高体鰤、黄尾鰤的遗传距离也较近,而与半滑舌鳎和大菱鲆的遗传距离则较远。半滑舌鳎和大菱鲆属于鲽形目(Pleuronectiformes)鱼类,而卵形鲳鲹与高体鰤、黄尾鰤则均属于鲹科(Carangidae)鱼类,具有更近的亲缘关系。MRF家族基因的聚类树状图并不代表这些物种的进化历史,但与这些物种的系统分类学结果高度一致。
MRF家族基因是动物肌肉发育过程中重要的正向调控因子,其在不同发育阶段和组织中的分布存在差异。对卵形鲳鲹13个胚胎发育时期的研究结果表明,的表达量在卵形鲳鲹胚胎发育过程中呈现先升高后降低的趋势,在受精卵到胚体形成期的前9个时期表达量极低,而在眼囊期、耳囊期与心脏跳动期表达量迅速升高,到晶体出现期表达量又有所降低,直到孵化期均维持较低的表达量,但仍显著高于受精卵到胚胎形成期(<0.05),推测其可能在眼囊、耳囊与心脏跳动期发挥主要的生肌调节作用,且在维持肌肉的形态或后续肌肉发育中仍发挥重要作用。对维持肌肉稳态、调节肌细胞融合以决定肌肉纤维的数量和大小至关重要。杨建等对斑马鱼的研究发现,基因在囊胚期和原肠胚期表达量很低,在体节期和咽囊期表达量迅速升高,并在孵化期开始下降,其结果与本研究结果相似。其余4个基因1、2、5与6在卵形鲳鲹胚胎发育过程中的表达模式则较相似,原肠中期前表达量较低,而从原肠末期到孵化期表达量迅速升高,这暗示、5与6基因从原肠末期开始发挥生肌作用。在脊椎动物发育过程中,原肠期是体轴建立和中内胚层形成的重要时期。原肠胚形成过程中胚层的形态发生涉及一个复杂的机制,该机制调节细胞群的增殖、运动和模式化,以及遗传和信号活动中开关的编排,可能驱动胚胎的谱系规范和组织建模。在原肠胚形成过程中,与多能性调控、生殖层形成、转录调控、细胞代谢、转运和离子内稳态相关的基因高度富集,并伴随着组织特异性基因和形态发生驱动因子的激活而向器官发生发展。重要的是,中晚期原肠胚转变过程中发生了转录组的重大改变,表现为多能性相关基因的下调,TGFβ/BMP信号通路相关基因的高表达。本研究在卵形鲳鲹原肠中期到末期的转变过程中,也发现MRF家族4个基因1、2、5与6转录表达的迅速升高。这意味着1、2、5与6基因从原肠末期开始发挥重要的作用,这与骨骼肌在原肠期开始形成在时间上相吻合。然而,对鳜()胚胎中MRF家族基因的研究发现,5、和在神经胚期表达量急剧增加,这可能意味着MRF基因在不同鱼类不同发育阶段的功能与表达存在差异,不同鱼类MRF家族基因的具体功能有待进一步研究。
