小菜蛾Btk抗性品系对四种Bt杀虫晶体蛋白抗性发展的研究
2022-04-22马明勇张政兵
符 伟,唐 涛,王 培,王 勇,尹 丽,马明勇,张政兵*
(1. 湖南省农业科学院植物保护研究所,长沙 410125;2. 湖南省植保植检站,长沙 410005)
小菜蛾Plutella xylostella(L.)是十字花科作物上一种世界性重要害虫[1],全世界每年用于防治小菜蛾的费用高达40~50亿美元[2,3]。目前,我国对小菜蛾的控制策略仍以化学防治为主。然而,化学农药的大量连续使用,导致其对化学药剂的抗性水平上升,而与之相应的是田间防治效果逐渐下降[4]。因此,用以苏云金芽胞杆菌(Bacillus thuringiensis,Bt)为代表的生物农药防治小菜蛾近年来日益受到关注和应用。
苏云金芽胞杆菌是一种从土壤或死亡昆虫体内分离出来的革兰氏阳性细菌。它在芽胞形成时期产生一种或多种杀虫晶体蛋白(Insecticidal crystal proteins,ICPs),可对鳞翅目等多种昆虫产生毒杀活性[5]。在Bt制剂及转Bt基因作物的应用与推广之初,不仅对田间害虫的有效控制成绩斐然;同时有利于提高作物的产量和质量,且减少了化学农药的使用频次与使用量,从而对农林业生产和环境保护发挥了积极的作用[6]。然而,随着时间的推移及广泛应用,部分害虫已对Bt产生了一定程度的抗药性。Tabashnik等[7]在长期使用Bt制剂的美国夏威夷田间首次发现小菜蛾对Bt 制剂产生抗药性。戴承镛等[8]首次报道了我国部分地区小菜蛾对Bt制剂的抗药性问题,其中华南地区(广州及深圳一带)对Bt制剂的抗性比扬州地区高10~25倍,而泰国曼谷地区的抗性又比华南地区高40%左右。Pardo-Lopez等[9]总结了Bt Cry毒素在鳞翅目昆虫中产生抗药性的多种机理,特别值得注意的是一种昆虫可能形成一套抗性机制对同种或是不同种Cry毒素都产生抗性。
苏云金芽胞杆菌的不同杀虫晶体蛋白由不同种类的Cry基因控制,而不同种类的Cry杀虫晶体蛋白对小菜蛾的杀虫活性不一致,而且在 Bt杀虫晶体蛋白的选择压力下,其抗性发展亦有所差别。王崇利等[10]报道广东惠州小菜蛾田间种群对Cry1Ab和Cry1Ac蛋白的抗药性达到高抗水平,但对Cry1Aa 和Cry2Aa仍然保持较高的敏感性。目前,用于防治小菜蛾的Bt菌株主要为库斯塔克亚种(Bt var.kurstaki,Btk),大多数含有对鳞翅目害虫有特异杀虫活性的Cry1Aa、Cry1Ab和Cry1Ac,其中Cry1Ac是杀虫毒力最高的基因[11]。已有研究表明,基因叠加策略可为害虫 Bt抗性治理策略提供新途径,即利用无交互抗性的杀虫晶体蛋白组合来拓宽与增强转基因作物的抗虫谱,从而使得靶标害虫极难对转基因作物产生抗性[12]。
小菜蛾田间种群对 Bt制剂及不同杀虫晶体蛋白抗性水平屡有报道,但田间害虫产生抗性影响因素较为复杂。为此,本研究中使用Btk制剂汰选获得小菜蛾抗性品系(F80代和F100代),采用叶片浸渍法测定了Cry1Ab、Cry1Ac、Cry1Ah和Cry1Ca杀虫晶体蛋白对小菜蛾抗性品系的室内毒力,旨在明确主效基因为Cry1Ac的Btk制剂选择压力下小菜蛾对Cry1Ac的抗性发展趋势,以及与其他Cry1类杀虫晶体是否存在交互抗性,从而为Bt多价杀虫晶体蛋白的应用与小菜蛾的抗性治理提供理论基础。
1 材料与方法
1.1 供试昆虫
小菜蛾相对敏感品系(SS):将2003年采自广州的田间品系于中国农业科学院蔬菜花卉研究所养虫室内用无虫甘蓝苗继代饲养,期间未接触任何杀虫剂。小菜蛾Btk抗性品系:该种群由2005年采自上海市郊的甘蓝种植田间的小菜蛾蛹和幼虫建立的,随后采用Btk可湿性粉剂浸渍甘蓝苗叶片后喂食小菜蛾,连续汰选80代和100代后,分别获得F80代和F100代供试试虫,二者对Btk的抗性倍数分别大约为1800倍和1900倍。
1.2 供试药剂
苏云金芽胞杆菌库斯塔克亚种可湿性粉剂由湖北省农业科学院Bt研究开发中心提供,效价为16000 IU/mg,属Bacillus thuringiensissubsp.kurstaki。Cry1Ac原毒素从Bt标准菌株HD-1中提取获得,消化后的Cry1Ab、Cry1Ah及Cry1Ca原毒素由中国农业科学院植物保护研究所提供,用牛清蛋白BSA结合SDS-PAGE进行Bradford定量,存储于-80 ℃待用[13]。
1.3 室内毒力测定
参照Tabashnik和Cushing[14]的叶片浸渍饲喂法。先进行预备试验,然后根据预备试验的结果,以校正死亡率在10%~90%的浓度范围作为正式试验的浓度范围。先用0.1% TritonX-100水溶液稀释药剂,按等比级数稀释法,顺次配制成5~7个系列浓度,以0.1% TritonX-100水溶液作为对照。将直径为5 cm的新鲜甘蓝苗嫩叶蝶在系列浓度的药液中浸10 s,自然晾干后转入培养皿(底部铺一层浸过适量蒸馏水的滤纸片用于保湿)中。每皿接入10头3龄初期幼虫,每处理4次重复。然后,将其置于培养箱中,温度(25±1)℃,光周期16L∶8D,相对湿度70%,72 h后检查幼虫死亡数。最后,用Abbott公式计算各处理的校正死亡率,并运用DPS软件计算供试药剂的LC50值、95%置信限、毒力回归方程(y=a+bx)相关系数及卡方值。抗性倍数(resistance ratio,RR)=抗性品系的LC50/相对敏感品系的LC50。
2 结果与分析
2.1 对小菜蛾相对敏感品系的室内毒力
Cry1Ab、Cry1Ac和Cry1Ca对小菜蛾相对敏感品系有较好的杀虫活性,其中以Cry1Ac最好,LC50为0.71 mg/L;Cry1Ah的活性较差,LC50高达40.03 mg/L(表1)。

表1 四种Bt杀虫晶体蛋白对小菜蛾敏感品系的室内毒力Table 1 Toxictiy of four Bt-insecticidal crystal proteins against Bt-susceptible strain ofP. xylostella
2.2 对小菜蛾Btk抗性品系的室内毒力
不同Bt杀虫晶体蛋白对Btk抗性种群毒力有较大的差异。Cry1Ca对F80代及F100代抗性品系的活性均最高,LC50约12.07~21.37 mg/L。Cry1Ac对F80代的活性中等(LC50为47.75 mg/L),而对F100代的活性较低(LC50为75.72 mg/L)。Cry1Ab对F80代的活性最低(LC50为65.43 mg/L)而对F100代的活性较低(LC50为72.18 mg/L)。Cry1Ah对F80代及F100代的活性均低,LC50约65.21~100.18 mg/L(表2)。
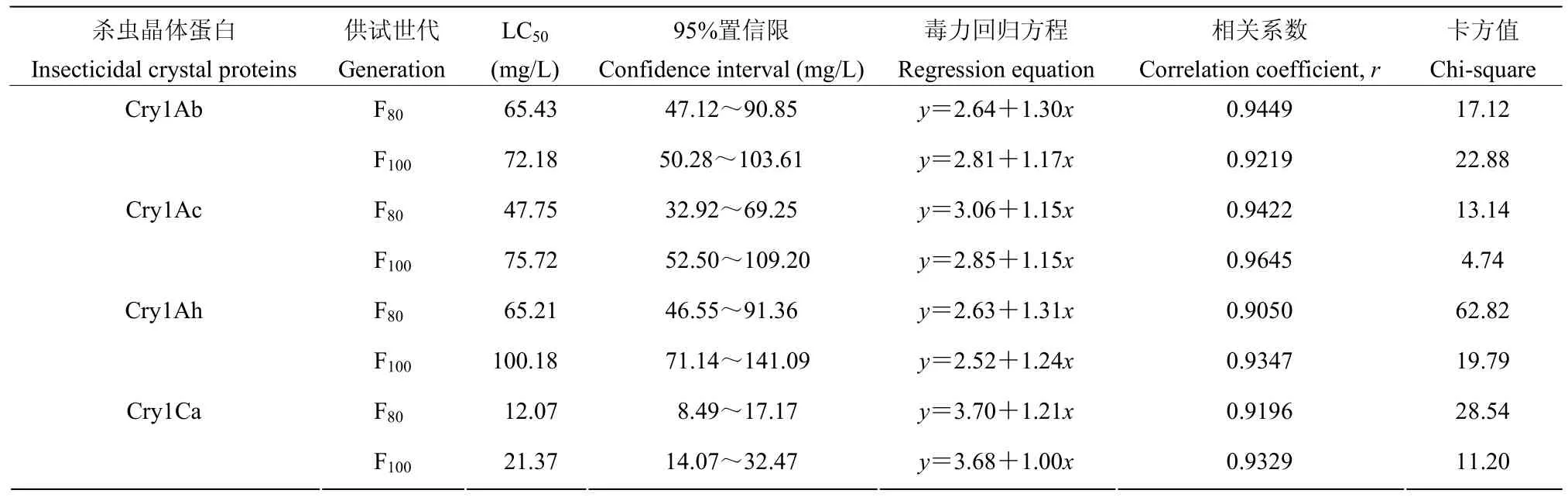
表2 四种Bt杀虫晶体蛋白对小菜蛾Btk抗性品系的室内毒力Table 2 Toxicity of four Bt-insecticidal crystal proteins against Bt-resistant strain ofP. xylostella
2.3 小菜蛾Btk抗性品系对四种杀虫晶体蛋白的抗性发展规律
经Btk制剂汰选80至100代后,小菜蛾Btk抗性品系对Cry1Ac的抗性发展较快,其抗性倍数高达67.3~106.8倍;对Cry1Ab的抗性发展速度次之,其抗性倍数达60.0~66.1倍;对Cry1Ca和Cry1Ah的抗性发展缓慢,其抗性倍数分别为3.3~6.0倍和1.6~2.5倍(图1)。

图1 小菜蛾Btk抗性品系对四种Bt杀虫晶体蛋白的抗性演变Fig. 1 The resistance development of four insecticidal crystal proteins in the Bt-resistant strain ofP. xylostella
3 讨论
1986年,Halpin[15]首次发现美国夏威夷长期使用Bt制剂的田间小菜蛾已对Bt制剂产生了抗药性。迄今为止,全世界已有美国、巴西、中国、马来西亚、菲律宾及日本等地相继报道小菜蛾对Cry1Aa、Cry1Ab、Cry1Ac、Cry2Aa、Cry2Ab、Cry2B、Cry1Ca和 Cry1Da等Bt杀虫晶体蛋白存在不同程度的抗药性[16]。研究表明,小菜蛾不仅在田间表现出对Bt杀虫晶体蛋白的抗药性,而且在室内可通过药剂筛选表现出较高的抗性水平。Guo等[17]在室内分别用Cry1Ac杀虫晶体蛋白和Btk制剂继代汰选获得SZ-R及SH-R品系小菜蛾,其抗性倍数依次高达450倍和1900倍。本研究结果表明,在主效基因为Cry1Ac的Btk制剂的连续汰选压力下,小菜蛾对Cry1Ac的抗性上升速度最快,并且对Cry1Ab产生较高的交互抗性,对Cry1Ah和Cry1Ca的抗性发展较慢。因此,为延缓小菜蛾对Bt抗性的产生与发展,应设法减小选择压力,以保护小菜蛾对Bt的敏感性。
不同Bt杀虫晶体蛋白的组合混用亦是抗性治理的重要策略之一[18]。该策略既可应用于传统的Bt制剂加工及后期的施用,又可用于转基因植物的选育与应用。如Cao等[19]将Cry1Ac和Cry1C同时导入青花菜中,结果表明:转双价基因的青花菜比转单价的青花菜具有更好的杀虫效果,且前者对抗Cry1Ac和Cry1C杀虫晶体蛋白的小菜蛾均有毒杀作用。然而,小菜蛾对不同的Bt杀虫晶体蛋白可能存在交互抗性。为此,仅仅简单地将不同Bt杀虫晶体蛋白混用,则难以达到抗性治理的最佳效果,而是在混用前必须针对Bt杀虫晶体蛋白的交互抗性进行充分研究。
