球孢白僵菌BB-T02生物学特性及对两种检疫性粉蚧的侵染活性
2022-04-22姚锦爱余德亿侯翔宇
黄 鹏,姚锦爱,余德亿,侯翔宇
(福建省农业科学院植物保护研究所/福建省作物有害生物监测与治理重点实验室/福建省作物有害生物绿色防控工程研究中心,福州 350013)
南洋臀纹粉蚧Planococcus lilacinus和石蒜绵粉蚧Phenacoccus solani是两种世界性花果害虫,南洋臀纹粉蚧可为害可可、番荔枝、红毛榴莲、番石榴、柑橘、美洲木棉、羊蹄甲、变叶木、桅子花和杜鹃等35个科100余种水果、林木和观赏植物[1,2],在2007年被我国列入进境植物检疫性有害生物[3];石蒜绵粉蚧可为害石蒜科、景天科、仙人掌科、爵床科、番杏科、大戟科、马齿苋科、菊科、豆科和茄科等40科101属131种观赏植物和经济作物[4,5],在2014年被国家质量监督检验检疫总局列为进口多肉植物重点关注的有害生物[6]。虽然这两种检疫性粉蚧目前在国内尚未有广泛分布的报道,但本研究前期调查发现它们分别已在福建省内的番荔枝果园和多肉植物种植基地发生为害严重,世代重叠明显,若虫和成虫常群集刺吸为害果实、叶片、幼芽和嫩茎等部分,引起落花、落叶、落果和长势衰弱,同时还分泌蜜露诱发煤烟病,直接或间接影响番荔枝和多肉植物的品质和经济价值,急需防控。
南洋臀纹粉蚧和石蒜绵粉蚧体表覆盖大量蜡质,不易防治,目前生产上采用的防控措施还偏重于使用化学农药,但长期使用化学农药,易造成“3R”、环境污染和生物多样性下降等负面问题,与我国种植业的绿色引领导向冲突不断,需寻找更加环保的措施协同控害。生物防治是一种安全、有效、持久的控害方法,其中挖掘、利用虫生真菌已是害虫生防的重要发展方向和研究热点之一[7,8]。目前,全世界已记载的虫生真菌约1000多种[9],但仅金龟子绿僵菌Metarhizium anisopliae中的F061和FM-03等菌株被挖掘用于防控这2种检疫性粉蚧[10,11],可供选择的菌株资源仍相对较少,因此有必要再寻找一些高效的虫生真菌菌株以减轻它们的为害。球孢白僵菌Beauveriabassiana是目前世界上研究和应用最多的虫生真菌之一,对人、畜、作物、害虫天敌及自然环境相对安全,已被广泛用于害虫生物防治[12,13];其中菌株BB-T02已被证实对蓟马的致死效果达87%以上[14]。本研究以南洋臀纹粉蚧和石蒜绵粉蚧的雌成虫(体型最大、体表蜡质最厚、取食和为害能力最强)为供试虫源,在菌株生物学特性的基础上,测定菌株BB-T02对两种检疫性粉蚧的致病力和胞外酶活性,明确该菌株对两种检疫性粉蚧的生防潜力,为利用虫生真菌防控检疫性粉蚧提供技术支持。
1 材料与方法
1.1 供试菌株与虫源
供试球孢白僵菌BB-T02从受感染的马尾松毛虫Dendrolimus punctatus幼虫上分离获得,已保藏于中国微生物菌种管理委员会普通微生物中心(保藏号:CGMCC No.14130)[14],活化采用察氏培养基(NaNO33 g、KH2PO41 g、MgSO4·7H2O 0.5 g、KCl 0.5 g、FeSO40.01 g、蔗糖 30 g、琼脂粉 20 g和蒸馏水 1000 mL);取斜面保存的菌株BB-T02接种到培养基平板(d=9.0 cm,下同)上,置于温度(25±1)℃的HGZ-150型光照培养箱内活化培养10 d备用。供试南洋臀纹粉蚧和石蒜绵粉蚧分别采集于福建省漳州市诏安县金星乡的番荔枝果园和龙海市九湖镇的多肉种植基地,在室温 25~28 ℃、相对湿度 50%~60%、光周期12L:12D条件下分别用番荔枝果实和多肉植物“红心莲”饲养,取次代蜕皮3 d的雌成虫备用。
1.2 球孢白僵菌BB-T02生物学特性观察
1.2.1 温度对菌株BB-T02生长和产孢的影响 参考徐金柱等[15]的方法挑取菌株BB-T02的单孢接种于察氏培养基平板中央,然后分别置于24、28和32 ℃等3个温度的培养箱中黑暗培养,每个处理3次重复;培养10 d后,用十字交叉法测定菌落直径;再往每个处理培养皿中加入10 mL含0.05%吐温-80的无菌水,用灭菌玻片刮下分生孢子配制成孢悬液,经3层纱布过滤后,用纽鲍尔血球计数板在E200型生物显微镜(尼康仪器(上海)有限公司)下观察、测定每个处理的产孢量。
1.2.2 光照对菌株BB-T02生长和产孢的影响 挑取菌株BB-T02的单孢接种于察氏培养基平板中央,然后分别置于光周期16L:8D、12L:12D和8L:16D等3种光周期及温度(28±1)℃光照培养箱中培养,每个处理3次重复;培养10 d后,采用1.2.1的方法测定菌落直径和产孢量。
1.2.3 碳源对菌株BB-T02生长和产孢的影响 参考黄鹏等[16]的方法,在察氏培养基的基础上,分别以葡萄糖、蔗糖和麦芽糖等3种材料作为供试碳源,制成3种试验培养基,每个处理3次重复;挑取菌株BB-T02的单孢接种于察氏培养基平板中央,再置于温度(28±1)℃、光周期8L:16D的光照培养箱中培养10 d,采用1.2.1的方法测定菌落直径和产孢量。
1.2.4 氮源对菌株BB-T02生长和产孢的影响 在察氏培养基的基础上,以1.2.3筛选的材料作为碳源,再分别以硝酸钠、蛋白胨和牛肉膏等3种材料作为供试氮源,制成3种试验培养基,每个处理3次重复;挑取菌株BB-T02的单孢接种于察氏培养基平板中央,再置于温度(28±1)℃、光周期8L:16D的光照培养箱中培养10 d,采用1.2.1的方法测定菌落直径和产孢量。
1.3 球孢白僵菌BB-T02侵染活性测定
1.3.1 致病力测定 参考黄鹏等[10]的方法,在生物学特性观察的基础上,取单孢培养的菌株 BB-T02,先往培养皿中加入10 mL含0.05%吐温-80的无菌水,再用灭菌玻片刮下分生孢子,后经3层纱布过滤得孢子原液,充分混匀后,计算孢子浓度;用无菌水将孢子原液稀释成 1×108、1×107、1×106、1×105和 1×104孢子/mL 5个处理浓度备用,以0.05%吐温-80无菌水为对照,每个处理3次重复。针对南洋臀纹粉蚧,取直径12 cm、高10 cm的塑料保鲜盒,每个保鲜盒中放1个生长90 d的番荔枝果实;在每个保鲜盒的盖子上挖取直径6 cm的透气孔,并用胶水将1块直径8 cm 的防虫网(80目)粘在透气孔上方;用毛笔分别挑取健康的南洋臀纹粉蚧雌成虫30头,在各浓度处理菌液中浸泡10 s后挑到滤纸上,晾干2 min,再将南洋臀纹粉蚧挑到番荔枝果实上,盖上保鲜盖。针对石蒜绵粉蚧,取直径9 cm的培养皿,依次铺入相同直径的花泥和滤纸各1片,花泥厚0.5 cm吸水6 mL,滤纸上再放1片多肉植物“红心莲”的叶片,叶柄用湿棉花包裹并紧贴滤纸;接着用软毛笔分别挑取石蒜绵粉蚧雌成虫 30头,在各浓度处理菌液中浸泡10 s后挑到滤纸上,晾干2 min,再将石蒜绵粉蚧挑到叶片上,盖上培养皿。将保鲜盒和培养皿置于温度(28±1)℃、相对湿度(90±5)%、光周期8L:16D的MGC-350HP-2型人工气候箱(上海一恒科学仪器有限公司)内,每天定时观察并挑除死亡虫体,以虫体长出菌丝确认为有效侵染,观察至所有处理中试虫的死亡数停止发生变化为止。统计2种检疫性粉蚧的死亡率,计算菌株BB-T02致死中浓度LC50和不同浓度孢悬液下的致死中时LT50,并参考黄鹏等[14]的方法对死亡虫体上的致病菌再进行分离纯化和电子显微形态鉴定。
1.3.2 胞外酶活性测定 参考Lu等[17]的方法制作蛋白酶、几丁质酶和脂肪酶3种胞外酶液,并测定它们的活性。胞外酶液制作:取1×108孢子/mL的菌株BB-T02孢子液1 mL分别加入到含99 mL蛋白酶诱导培养基(明胶10 g,K2HPO4、NaCl和MgSO4·7H2O各0.3 g,溶于1 L浓度为0.02 mol/L、pH 7的磷酸盐缓冲液中)、几丁质酶诱导培养基(蛋白胨5 g,K2HPO4、KCl和MgSO47·H2O各0.5 g,ZnSO4·7H2O 1 g,胶状几丁质10 g,溶于1 L蒸馏水中)或脂肪酶诱导培养基(葡萄糖2 g,蛋白胨5 g,MgSO4·7H2O 0.1 g,K2HPO41 g,橄榄油20 mL,溶于980 mL蒸馏水中)的三角瓶中,置于25 ℃、180 r/min的QYC-2102C型全温培养摇床(上海新苗医疗器械制造有限公司)上培养1~10 d;每天分别取2 mL的3种培养液用12000 r/min的KDC-140HR型高速冷冻离心机(安徽中科中佳科学仪器有限公司)离心30 min,取上清液用于测定酶活性;每处理3个三角瓶,即重复3次。蛋白酶活性测定:以酪蛋白为底物,测定蛋白酶活性;以失活的蛋白酶溶液作为空白对照,利用UV-1800PC型紫外可见分光光度计(上海美谱达仪器有限公司)测定各组反应混合液在680 nm处的吸光度值;根据L-酪氨酸标准曲线,计算各组反应混合液的蛋白酶活性,以每分钟催化分解酪蛋白生成1 μmol酪氨酸的酶量为1个酶活单位。几丁质酶活性测定:以胶状几丁质为底物,测定几丁质酶活性;以失活的几丁质酶溶液作为空白对照,测定各组反应混合液在585 nm处的吸光度值,以每分钟催化分解几丁质生成1 μmol N-乙酰氨基葡萄糖的酶量为1个酶活单位;根据N-乙酰氨基葡萄糖标准曲线,计算各组反应混合液的几丁质酶活性。脂肪酶活性测定:以对硝基苯棕榈酸酯为底物,测定脂肪酶活性;失活的脂肪酶溶液作为空白对照,测定各组反应混合液在410 nm处的吸光度值;根据对硝基苯酚标准曲线,计算各组反应混合液的脂肪酶活性,以每分钟催化分解脂肪成1 μmol对硝基苯酚的酶量为1个酶活单位。
1.4 数据统计与分析
利用DPS 7.05中的Tukey检验法分别对球孢白僵菌BB-T02在不同温度、光照、碳源和氮源下的菌落直径和产孢量进行多重比较;利用机率值分析法计算菌株BB-T02对2种检疫性粉蚧的致死中浓度LC50和致死中时LT50,取实际累计死亡率达50%以上处理的计算数值作为有效结果。利用GraphPad Prism 7绘制菌株BB-T02胞外酶活性的变化趋势图及其他相关图片。
2 结果与分析
2.1 球孢白僵菌BB-T02生物学特性
2.1.1 温度对菌株BB-T02生长和产孢的影响 菌株BB-T02生长和产孢受温度影响较大。生长速度以28℃最快,培养10 d后菌落直径达4.52 cm,虽与24 ℃差异不显著,但显著大于32 ℃;产孢量仍以28 ℃最高,达2.67×108孢子/cm2,显著高于其他两个处理(图1A)。

图1 菌株BB-T02在不同温度(A)、光照(B)、炭源(C)和氮源(D)下培养10 d后的菌落直径和产孢量Fig. 1 Colony diameter and sporulation of strain BB-T02 in different temperatures (A), photoperiods (B), carbon sources (C) and nitrogen sources (D) after ten days cultivation
2.1.2 光照对菌株BB-T02生长和产孢的影响 菌株BB-T02生长和产孢受光照的影响明显不同。生长速度依次为光周期16L:8D、12L:12D和8L:16D,培养10 d后菌落直径分别在4.58~4.89 cm,但三者间的差异不显著;产孢量以光周期8L:16D最高,达3.19×108孢子/cm2,显著高于其他两个处理(图1B)。
2.1.3 碳源对菌株BB-T02生长和产孢的影响 菌株BB-T02在不同碳源下的生长速度和产孢量差异明显。生长速度以麦芽糖最快,培养10 d后菌落直径达5.27 cm,显著大于其他两个处理;产孢量仍以麦芽糖最高,达3.59×108孢子/cm2,虽与蔗糖差异不显著,但显著大于葡萄糖(图1C)。
2.1.4 氮源对菌株BB-T02生长和产孢的影响 菌株BB-T02在不同氮源下的生长速度和产孢量明显不同。生长速度以牛肉膏和蛋白胨最快,培养10 d后菌落直径分别达5.65 cm和5.60 cm,两者差异不显著,但均显著显著大于NaNO3;产孢量以蛋白胨最高,达4.22×108孢子/cm2,显著高于其他两个处理(图1D)。
2.2 球孢白僵菌BB-T02侵染活性
2.2.1 致病力 菌株BB-T02能有效侵染南洋臀纹粉蚧和石蒜绵粉蚧(图2A,B),接菌10 d后的LC50分别为5.00×105孢子/mL和2.17×105孢子/mL(表1);两种粉蚧虽然在1×104和1×105孢子/mL处理的死亡率均未达 50%,机率值分析法计算出的 LT50均相对偏大,超出试虫停止死亡的时间;但在其他 3个处理的死亡率均有明显提高,LT50也均随菌液浓度的升高而缩短,特别是1×108孢子/mL处理浓度的死亡率分别达85.39%和88.76%(图2C),LT50分别为5.78 d和5.19 d(表2);对死亡虫体上的菌株再进行分离纯化和电子显微形态鉴定,发现再分离菌株的形态特征与供试菌株[14]一致,确定致病菌株为BB-T02(图2D)。
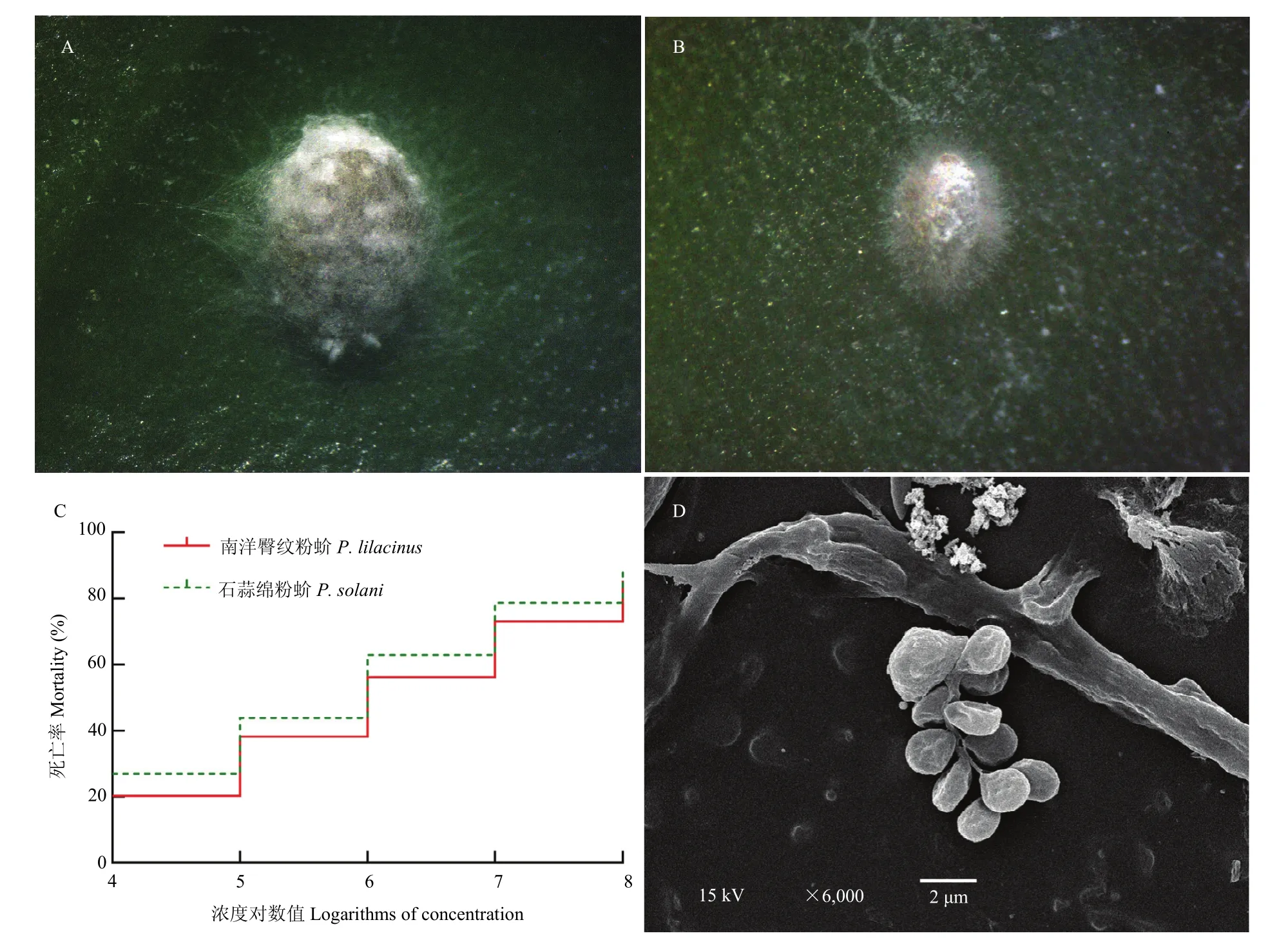
图2 菌株BB-T02对南洋臀纹粉蚧(A)和石蒜绵粉蚧(B)的侵染及两种粉蚧受侵染10 d后的死亡率(C)和再分离菌株形态特征(D)Fig. 2 P. lilacinus(A) andP. solani(B) infected by strain BB-T02, the mortality of two mealybugs (C) and morphological characteristics of reisolated strain (D) after ten days infection

表1 侵染10 d后菌株BB-T02对南洋臀纹粉蚧和石蒜绵粉蚧的LC50Table 1 LC50of strain BB-T02 againstP. lilacinusandP. solaniafter ten days infection

表2 侵染10 d后菌株BB-T02对南洋臀纹粉蚧和石蒜绵粉蚧的LT50Table 4 LT50of strain BB-T02 againstP. lilacinusandP. solaniafter ten days infection
2.2.2 胞外酶活性 菌株BB-T02的3种胞外酶中,蛋白酶的上升和下降幅度均较大,在第5 d到峰值22.68 U/mL,但在第10 d时仍维持在14.60 U/mL;几丁质酶和脂肪酶活性的变化趋势相似,上升和下降幅度则相对较为平稳,均在第6 d分别达到峰值13.19 U/mL和9.77 U/mL,在第10 d时分别维持在8.76 U/mL和5.19 U/mL(图3)。

图3 菌株BB-T02的胞外酶活性动态Fig. 3 Activities dynamic of extracellular enzymes of strain BB-T02
3 讨论
虫生真菌是农作物害虫的一类重要致病菌[8];球孢白僵菌作为虫生真菌的主要种类之一,已被广泛用于害虫生物防治[12,13]。菌株的生长速度及产孢量等生物学特性是衡量虫生真菌优良与否的重要指标,主要受温度、光照、碳源和氮源等条件影响,因此明确虫生真菌生长和产孢最适的条件,获得菌株最快生长和最高产孢量,对虫生真菌的生产和应用意义重大[18,19]。球孢白僵菌不同菌株间的生物学特性存在差异,如钱晶晶等[20]发现球孢白僵菌 Bb01生长和产孢最适的温度为26 ℃;齐永霞等[21]发现球孢白僵菌Bb06生长和产孢适宜的碳源为白砂糖、氮源为蛋白胨;刘晓晓等[22]发现球孢白僵菌B-2生长和产孢最适的温度也是26 ℃,但碳源为葡萄糖、氮源为酵母浸出液;而本研究发现球孢白僵菌 BB-T02生长和产孢最适的温度为28 ℃、光周期为8L:16D、碳源为麦芽糖、氮源为蛋白胨,这些差异可能是由于地理和寄主来源不同所致。
致病力是衡量虫生真菌生防潜力和应用前景的重要指标[14,16];本研究发现球孢白僵菌BB-T02对南洋臀纹粉蚧和石蒜绵粉蚧雌成虫的侵染致死效果均随菌液孢子浓度升高而上升,累计致死率、LC50和 LT50最终分别可达85.39%和88.76%、5.00×105孢子/mL和2.17×105孢子/mL及5.78 d和5.19 d,与黄鹏等[10,11]利用金龟子绿僵菌F061和FM-03侵染这2种检疫性粉蚧的效果类似。而虫生真菌在侵染昆虫的过程中,蛋白酶、几丁质和脂肪酶等胞外酶对昆虫体壁(主要成分包括蛋白质、几丁质和脂肪)的降解具有极其重要的作用,其活性值大小被认为是决定菌株毒力的主要因素之一[23-25];本研究发现球孢白僵菌BB-T02的蛋白酶活性变化幅度较大,到达峰值的时间较短,推测这应该是菌株BB-T02能够有效突破2种检疫性粉蚧体壁中实现侵染致死的主要原因;而几丁质酶和脂肪酶活性虽然到达峰值的时间稍长,但变化幅度相对较平稳,推测这可能是菌株BB-T02能够持续突破和瓦解两种检疫性粉蚧体壁的另一个重要原因。可见,球孢白僵菌BB-T02对南洋臀纹粉蚧和石蒜绵粉蚧中体型最大、体表蜡质最厚、取食和为害能力最强的雌成虫具有强的侵染活性,可以预见该菌株对两种粉蚧的若虫和雄成虫也会具有高致病力,可用于这两种粉蚧的生物防治;研究结果丰富了两种检疫性粉蚧的生防微生物资源,也推动了球孢白僵菌等虫生真菌在粉蚧生防中的应用。
虫生真菌是生物农药的重要组成部分,只有在室内和田间对目标害虫均表现出良好侵染效果的菌株,才具有生防应用价值。因此,本研究后续将进一步细化研究球孢白僵菌BB-T02对南洋臀纹粉蚧和石蒜绵粉蚧其他虫态的侵染效果及对它们生长、生殖等生物学特性的影响,验证该菌株对两种检疫性粉蚧的田间防效,探讨该菌株的室内发酵方式及继代培养、保存方法,以开发出相应的生防产品,并延缓菌种退化和生防效价降低的问题,为利用虫生真菌防控这两种检疫性粉蚧及其他害虫提供技术支持。
致谢:本研究采集的南洋臀纹粉蚧和石蒜绵粉蚧承蒙北京林业大学武三安教授帮助鉴定,在此深表感谢!
