环境因子对中华双扇蕨孢子萌发的影响
2022-04-14欧阳康婷庞金辉余小玲许展慧任海王发国马国华刘宗君胡明锋张亚坚谭广文
欧阳康婷, 庞金辉, 余小玲, 许展慧, 任海*, 王发国*,马国华, 刘宗君, 胡明锋, 张亚坚, 谭广文
环境因子对中华双扇蕨孢子萌发的影响
欧阳康婷1,2, 庞金辉1,2, 余小玲3, 许展慧1,2, 任海1,2*, 王发国1,2*,马国华1, 刘宗君4, 胡明锋4, 张亚坚4, 谭广文5
(1. 中国科学院华南植物园,广东省应用植物学重点实验室,广州 510650;2. 中国科学院大学,北京 100049;3. 仲恺农业工程学院,广州 510650;4. 广东南岭国家级自然保护区管理局,广东 韶关 512727;5. 广州普邦园林股份有限公司, 广州 510650)
为了解珍稀濒危植物中华双扇蕨()濒危原因,研究了温度和湿度对其孢子萌发的影响。结果表明,中华双扇蕨孢子萌发能力较强,萌发方式为型,配子体萌发为型。中华双扇蕨孢子繁殖不受温度影响,而湿度显著影响孢子繁殖过程,湿润环境中孢子正常萌发,并形成心形配子体,顺利长出幼孢子体;而干旱环境下孢子难以萌发,配子体形态不规则发育,褶皱增加。中华双扇蕨在不适环境下孢子繁殖存在障碍,可能是导致其野生种群及数量下降的原因之一,对这类珍稀濒危植物的就地和迁地保护需要注意营造湿润的生境。
中华双扇蕨;环境因子;组织培养;配子体;形态发育
许多蕨类植物由于自然和人为干扰而处于珍稀濒危状态,急需开展繁殖过程研究,为保护和利用提供支撑。蕨类植物是具有两个能独立生活的植物体,即孢子体和配子体[1]。孢子体和配子体的生活策略并不相同。在野外难以发现蕨类配子体,而孢子繁殖研究有助于揭示蕨类孢子-配子体-孢子体的发育过程。目前,我国报道了孢子繁殖研究的仅有34科80属157种蕨类植物,占全国蕨类植物总数的5%[2],需要对更多的、尤其是珍稀濒危蕨类的孢子繁殖进行研究。
双扇蕨科(Dipteridaceae)属于真蕨纲薄囊蕨亚纲,起源于2.5亿年前的中生代早期,有7属百余种在中生代遍布全球,物种非常丰富,但至今仅存1属8种,分布在热带-亚热带地区,为孑遗植物[1,3]。从一个地质历史上分布很广泛的类群成为现在的孑遗类群,这可能与双扇蕨科植物的繁殖特征和适应能力有一定联系。中华双扇蕨()是双扇蕨科双扇蕨属植物,小型或中型草本,陆生或石缝生[4]。中华双扇蕨具有重要的观赏、药用和科研价值, 被列为珍稀濒危物种[5],也是广东和湖南省重点保护的野生植物,主要零星分布在中国广东、广西、重庆、贵州、云南、湖南,中南半岛和缅甸北部[1]。
目前,关于中华双扇蕨的研究主要有分类修正[6]、系统进化史评估[7]、形态学[8]和化合物提取及应用[9],对双扇蕨()[10]和喜马拉雅双扇蕨()[11]的配子体发育过程和生长素与生根关系的研究已有报道。Ouyang等[12]在南岭国家级自然保护区的中华双扇蕨群落调查中发现幼苗数量少,种群结构衰退,孢子实生苗极其少见。然而,中华双扇蕨孢子体积很小,通过风可进行远距离散播[13],遇适宜的生境便会萌发,推测中华双扇蕨孢子繁殖和配子体适应能力可能是导致该类群数量少且物种单一的原因。而配子体形态对蕨类演化和发育以及生态适应具有重要的意义[14]。本研究通过采集成熟孢子,设置不同温度和湿度的培养环境,探讨孢子萌发率和配子体形态特征的变化,为中华双扇蕨的保护和利用提供科学参考。
1 材料和方法
1.1 孢子材料
2020年12月初,在南岭国家级自然保护区(24° 42′~24°47′ N, 112°51′~112°57′ E)的中华双扇蕨种群中剪取3株中华双扇蕨成熟植株的1/2叶片(有成熟孢子囊),放进硫酸纸折成的简易信封中带回实验室,竖直放置在自然干燥、无风、避光环境下,约7 d后孢子囊因自然干燥开裂,孢子弹射在硫酸纸上, 用1 mL离心管将孢子收集,放入-4 ℃冰箱保存备用。
1.2 孢子培育和观察
取出装有孢子的离心管,加入1 mL 5% NaClO溶液,不间断振荡消毒10 min后,10 000×离心2 min,无菌水洗涤离心重复3次,倒出废液后, 制成孢子悬浊液。用已消毒灭菌的镊子沾取孢子均匀接种在1/2MS培养基中,设置自然温度(10℃,孢子成熟期的均温)、25℃、不加水和加2 mL水共4组处理,置于人工培养箱(RXZ-1000B型,宁波江南仪器厂)中培养,培养条件为温度10℃/25℃, 湿度78%~80%,光照14 h/d,黑暗10 h/d,光照强度约90mol/(m2·s)。以出现假根为标准计算萌发率[15]。统计萌发率后将其转到湿润和干旱泥炭土培养盒中继续培养,将2等份已处理的泥炭土用无菌水充分润湿后,将培养基中片状体均匀地移入培养盒中并盖好,相同的水分控制,湿润培养盒3~5 d加水1次,培养条件相同。以上操作均在超净台完成。
接种15 d后开始观察,在孢子开始萌发及配子体发育时期,选取典型材料制成临时装片,放置在电镜(JSM-6360LV, JEOL, Japan)和光学显微镜下(Olympus-BH2, Japan),并用ECLIPSE-E600相机(Nikon, Japan)观察记录,保持同倍数镜头随机选择5个视野,拍照计数20个孢子,以平均值为最终结果[16]。
1.3 数据处理
将所有数据录入Microsoft Excel 2013,采用R 3.6.3进行单因素方差分析,以<0.01表示差异显著,使用Adobe Photoshop CC 2019处理图片。
2 结果和分析
2.1 孢子囊和孢子形态
中华双扇蕨孢子囊近圆形、细小,橙黄色,肉眼无法观察(图1: A),具有纵生环带,中部开裂弹射孢子。成熟孢子为透明色或浅黄绿色,极面观为椭圆形,两端稍平钝;赤道面观为豆形, 但近极面较平;孢子大小约为20.1m×40.0m; 孢子为单列缝,几乎长达孢子赤道线(图1: B, C)。还观察到极个别孢子为3裂缝(图1: D)。孢子外壁厚约1.0m (图1: E),没有明显的分层现象,附有一层薄薄的外膜,表面光滑,未见清晰的纹饰(图1: F)。

图1 中华双扇蕨孢子囊和孢子。A: 孢子囊; B, C: 单裂缝孢子; D: 三裂缝孢子; E: 孢子壁; F: 孢子纹饰; As: 环带; Mi: 单裂缝; Ti: 三裂缝; Sd: 孢子壁; Me: 薄膜。
2.2 孢子萌发
孢子播种20~25 d体积开始稍微变大(图2: A),随后孢子内原生质逐渐收缩,同时产生若干个大小不一的叶绿体细胞(图2: B)。35 d后,孢子壁从裂缝处伸出1条假根,似圆状,无色透明,有一层薄膜(图2: C),随着假根伸长,可以清楚地观察到根部的绿色颗粒物质(图2: D)。初生假根一般垂直于近极面,首端不发生弯曲,极少数微微弯曲,向下生长(图2: E)。继续培养根部颗粒物逐渐消失,慢慢形成枝干形态(图2: F)。

图2 中华双扇蕨孢子萌发。A: 孢子变大; B: 初生假根(Pr); C: 假根内叶绿体; D: 初生假根基部膨大; E~F: 长假根。
2.3 片状体
继续发育至片状体阶段时,中华双扇蕨片状体形态变化多样,主要有片形片状体和立体片状体。中华双扇蕨的片状体有条形、圆形、方形、或者不规则形(图3: A~F),随着片状体细胞不断分化生长,其底部假根的数量也在增加。片状体的生长点两侧细胞分裂速率快慢,决定了后期原叶体的形状。随着片状体细胞不断分裂和发育,逐渐长出原叶体(配子体),但后期也会出现特殊形状的片状体(图3: F)。
2.4 原叶体
原叶体也称配子体,正常发育成熟的原叶体呈对称的心形(图4: A, B),原叶体边缘较平滑,假根在腹面两侧均有生长(图4: B),颜色也逐渐变成浅褐色(图4: B)。毛状体会在配子体时期出现,为乳突毛状体,散乱地分布在配子体上(图4: C)。性器官杂乱地出现在原叶体的腹部假根丛中,或者附着在边缘细胞的表面(图4: C, D)。中华双扇蕨的颈卵器,与大多数蕨类植物的颈卵器有着相同的特征,属于包埋型(图4: C)。颈卵器发生在配子体的表面细胞,其颈部由4列5层细胞构成。成熟的中华双扇蕨配子体腹面、中肋和边缘都分布着许多颈卵器, 待发育成熟后,顶端开裂;而精子器散生在原叶体边侧或背部,圆球形,与水龙骨科(Polypodiaceae)和紫萁科(Osmundaceae)植物精子器相似,均属于突出型(图4: D)。在有水环境下,精子器发育成熟释放大量的精子,精子进入颈卵器, 与卵结合进而发育成胚,最终形成幼孢子体(图4: E, F)。幼孢子体从成熟的原叶体腹面假根中长出,而原叶体逐渐变透明。一般幼孢子体为两侧对称的心形状,可以清晰看见孢子叶上有叶脉(图4: F)。
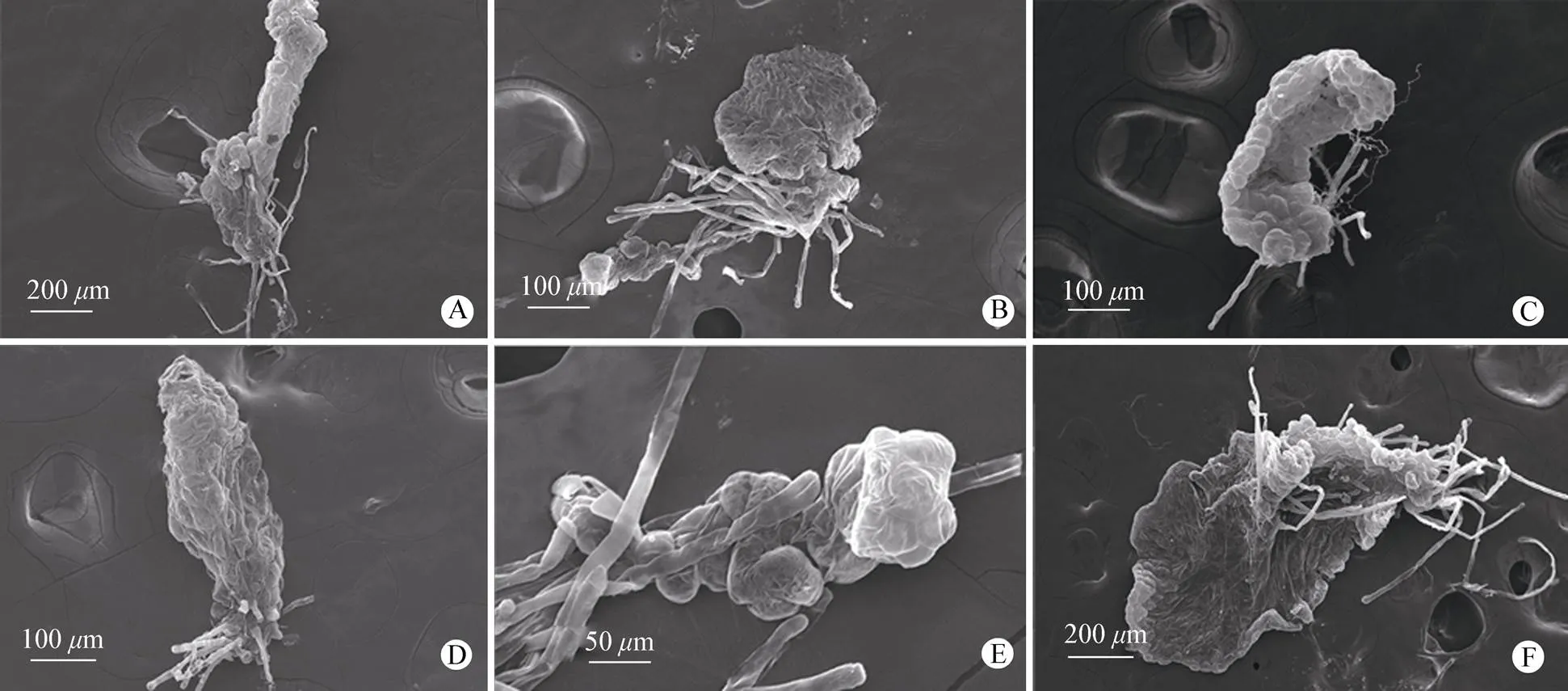
图3 中华双扇蕨的片状体。A: 条形; B: 圆形; C: 方形; D~E: 不规则形; F: 特殊形状片状体后期。

图4 中华双扇蕨原叶体。A, B: 心形原叶体; C: 颈卵器; D: 精子器; E: 幼孢子体; F: 心形幼孢子体; Mt: 乳突毛状体; Ap: 生长点; Rh: 假根; Ga: 配子体; Ve: 叶脉。
2.5 干旱培养条件下配子体形态特征
在干旱环境下培养,原叶体停止发育或呈不规则形状(图5: A~D)。背部和底部涌现大量假根(图5: A),假根由褐色逐渐加深,配子体的褶皱增加(图5: B~D),配子体的生长点附近保持绿色并缓慢生长,其余部分变为浅褐色。
2.6 孢子萌发率
由图6可见,在自然温度(10 ℃)和25 ℃下培养,除第26天自然温度的萌发率略低于25 ℃的外,两者的孢子萌发率没有显著差异。环境湿润程度对孢子的萌发率有显著影响(<0.01),与湿润环境相比,干旱条件下的孢子萌发率极低,仅在第50天才观察到极个别孢子有萌发迹象。
3 结论和讨论
3.1 孢子萌发过程
中华双扇蕨孢子繁殖周期约4个月左右,经历5个阶段,分别是孢子萌发及初生假根期、丝状体期、片状体期、原叶体期和幼孢子体期。首先单裂缝处伸出一条假根,透明伴有叶绿体,在发育过程中垂直于单裂缝方向伸长,偶见假根基部膨大或末端弯曲现象,这在蕨类孢子萌发中属常见现象[17]; 丝状体留存时间短,片状体一般为匙状,配子体多为雌雄同体,颈卵器为四面体,成熟精子器为圆球形。相比于不规则型的配子体来说,心型配子体更富有活力,较易观察到幼孢子体的产生,一般从配子体腹部长出,随着幼孢子体的生长,配子体逐渐凋零。

图5 干旱环境下中华双扇蕨的不规则型配子体(A~D)。Fo: 褶皱; Rh: 假根。

图6 温度和湿度对中华双扇蕨孢子萌发率的影响。**: P<0.01。
双扇蕨属萌发方式均为型[10–11],发育过程大致相似,但也存在些许差异。电镜观察表明,与双扇蕨孢子不同,中华双扇蕨孢子中有透明孢子和黄绿色孢子,数量相当,这在之前的研究中未见报道。黄绿色的孢子内部可能含有叶绿体,即绿色孢子,这种类型的孢子仅存在几个相互无关联的科和属中,或者部分孢子外壳为黄绿色。有人认为绿色孢子相较于非绿色孢子的寿命更短,萌发更快, 就像不耐干燥和贮藏的顽拗性种子[18–19]。Sandip-kuamr[11]报道喜马拉雅双扇蕨孢子在保存45 d后丧失萌发活力,而新鲜孢子萌发能力则很强,中华双扇蕨孢子的表现与之相同。最近的研究指出绿色孢子也具有干燥耐性,节节草()[20]、沼生木贼()[21]、欧紫萁()和荚果蕨()[22]的孢子在合适的储存方式下能延长孢子寿命。
在不利的环境下,与其同属蕨类不同的是,中华双扇蕨配子体会涌现乳突毛状体,而乳突毛状体被认为具有增强反光、抗旱、抵御昆虫啃食的能力[1],但现今未有毛状体作用机制的相关解释。另外,不利环境抑制了雌雄同体的配子体形成,其中,双扇蕨会产生许多精子器,并且有细长颈卵器,其颈部由7~9层细胞组成[10],而中华双扇蕨偏向产生更多的颈卵器,两者的颈卵器发育均与薄囊蕨类相同[1]。中华双扇蕨萌发趋于原始类型,以板块或团块状萌发,与紫萁科、里白科(Gleicheniaceae)相似[23–24]。然而,与系统发育树上最近的水龙骨科的萌发方式不同,水龙骨科发育方式有型和型[25]。这表明中华双扇蕨孢子萌发方式虽表现为原始类型,但黄绿色的孢子,乳突毛状体等现象的发生,说明中华双扇蕨处于较进化的位置,其配子体应具有一定的抗逆性。
3.2 湿度影响中华双扇蕨孢子萌发
中华双扇蕨孢子萌发类型属于型。在无菌培养中约20~25 d开始萌发,在不含任何营养物质的无菌水中也可萌发,说明中华双扇蕨孢子萌发要求不高,萌发能力强,不存在任何休眠期,因此,中华双扇蕨孢子的萌发特性不会导致其野外种群更新困难。而在不同温度、湿度和凋落物浓度培养条件下,温度对中华双扇蕨孢子萌发并没有影响,然而,湿度对孢子萌发有显著影响。首先,干旱环境下孢子不能顺利完成吸胀吸水过程。而中华双扇蕨恰巧在旱季(9月至次年1月份)产生孢子,降雨少,土壤极干,向下挖掘30 cm也不见潮湿的土壤,这可能是中华双扇蕨野生孢子实生苗少的原因之一。其次,中华双扇蕨的孢子遭遇长时间缺水可能引起其内部结构的变异、损坏,而无法开始萌发。
蕨类孢子的萌发机制遵循与种子相同的原理[26]。大多数蕨类孢子吸水后体积变大,内部亲水物质吸引水分子,代谢活动增强,启动一系列生理反应。然而,有研究表明仅10%的蕨类物种具有较好的耐干旱性[27]。蕨类孢子在遭遇干旱时,孢子细胞体积减小[28],细胞原生质或脂质体从流体结构变成固体结构[29],Walters[30]认为细胞在干燥环境中容易处于不稳定状态,极易超过细胞体积的最小阈值。相关研究也表明,固体结构中因分子的聚集阻碍了分子迁移速率,细胞基质可能丧失生存活力[31–33]。
3.3 干旱环境下中华双扇蕨配子体褶皱增加
中华双扇蕨配子体在适宜环境下可以顺利发育出性器官并完成受精,干旱条件下的中华双扇蕨配子体的体型变大,形状不规则化,假根增生,性器官数量比例分配不均。据此,我们猜测中华双扇蕨的配子体具有一定耐旱性,其复杂的形态可短暂地抵抗干旱。不同于种子植物通过增厚叶片,改变叶片角度等方式来减少水分蒸发,抗旱能力较强的蕨类配子体通常具有复杂的形态[34]。Watkins[35]研究发现蕨类配子体复杂多变的褶皱可能在干燥生境中对物种的生存起着重要作用。水龙骨科、骨碎补科(Davalliaceae)、条蕨科(Oleandraceae)等蕨类种群多达60%~70%为附生或干旱陆生物种,大多数配子体形态复杂多变,褶皱重叠弯曲[35],从而形成微小空间的高湿空气,延长并保持了湿润和光合作用的时间[36]。Pittermann等[37]报道短叶风铃草()复杂形态配子体的干燥速度远慢于具有简单形态的陆生植物糠秕藻()。因此, 我们推测中华双扇蕨配子体在遭遇干旱或缺乏自由水的情况下,通过丰富其配子体形态变化和生理活动调节来获得存活和完成受精的机会,还需进一步的实验观察证据。
中华双扇蕨的濒危原因包括生境胁迫、孢子萌发受阻和配子体发育畸形3个方面。历史时期中华双扇蕨由于古气候的重大变化和地质活动使得该蕨生境面积狭窄,呈零星分布;而在恶劣(干旱)的环境下,孢子难以萌发或配子体形态畸变导致其生存与扩繁的机会渺茫,种群数量不易增加,种群更新困难。孢子繁殖对于蕨类种群扩张和更新是极其重要的途径。因此,对中华双扇蕨实施就地保护过程中,可以在孢子萌发季节通过喷灌改善其萌发条件。在迁地保护时,营造湿润的生境很重要。当然,还可结合人工分株繁殖和人工孢子繁殖方法帮助育苗用于园林绿化。而针对濒危蕨类物种,应对其生境特点、孢子特性、配子体发育规律和繁殖生物学等进行系统研究来制定科学高效的保护措施。
致谢 感谢黄耀、周婉敏同学在野外采样工作中的帮助;感谢中国科学院华南植物园公共实验室胡晓颖老师在电镜观察实验中的悉心教学;感谢哈尔滨师范大学刘保东教授提供的帮助。
[1] LU S G, ZHANG G F, SU W H, et al. Pteridology [M]. Beijing: Higher Education Press, 2007: 12.
陆树刚, 张光飞, 苏文华, 等. 蕨类植物学[M]. 北京: 高等教育出版社, 2007: 12.
[2] YAN Y H, LIU H M, WANG F G. Fern garden in China’s main Botanical Gardens [J]. Landscape Archit, 2014(3): 16–21. doi: 10.3969/ j.issn.1000-0283.2014.03.003.
严岳鸿, 刘红梅, 王发国. 中国主要植物园中的蕨园[J]. 园林, 2014(3): 16–21. doi: 10.3969/j.issn.1000-0283.2014.03.003.
[3] SKOG J E. Biogeography of Mesozoic leptosporangiate ferns related to extant ferns [J]. Brittonia, 2001, 53: 236–269. doi: 10.1007/BF02 812701.
[4] WANG R J. The Key Protected Plants of Guangdong [M]. Guangzhou: Guangdong Science and Technology Press, 2019: 81.
王瑞江. 广东重点保护野生植物[M]. 广州: 广东科技出版社, 2019: 81.
[5] TAN H N, YANG Y, DONG S Y, et al. Threatened species list of China’s higher plants [J]. Biodiv Sci, 2017, 25(7): 696–744. doi: 10. 17520/biods.2017144.
覃海宁, 杨永, 董仕勇, 等. 中国高等植物受威胁物种名录[J]. 生物多样性, 2017, 25(7): 696–744. doi: 10.17520/biods.2017144.
[6] LU S G, TIAN Z J. A taxonomic revision of the Dipteridaceae from Yunnan, China [J]. Plant Sci J, 2011, 29(4): 432–434. doi: 10.3724/SP. J.1142.2011.40432.
陆树刚, 田治蛟. 云南双扇蕨科植物的分类订正研究[J]. 植物科学学报, 2011, 29(4): 432–434. doi: 10.3724/SP.J.1142.2011.40432.
[7] CHOO T Y S, ESCAPA I H. Assessing the evolutionary history of the fern family Dipteridaceae (Gleicheniales) by incorporating both extant and extinct members in a combined phylogenetic study [J]. Amer J Bot, 2018, 105(8): 1315–1328. doi: 10.1002/ajb2.1121.
[8] CHANDRA S, KAUR S. Vasculature of the rhizome in the fern genus dipteris[J]. Amer Fern J, 1993, 83(2): 71–72. doi: 10.2307/1547571.
[9] WANG K. Chemical constituents of 6 species, such as,, et al. [D]. Kunming: Kunming Institute of Botany, Chinese Academy of Sciences, 2010.
王扣. 苏铁蕨、纤枝金丝桃等六种植物的化学成分[D]. 昆明: 中国科学院昆明植物研究所, 2010.
[10] STOKEY A G. The gametophyte of[J]. Int J Plant Sci, 1945, 106(4): 402–411. doi: 10.1086/335312.
[11] BEHERA S K, RAWAT V K, SINGH A P, et al. Studies on the spore germination, developmental pattern and sexuality of gametophytes in(R. Br. ex Hook. et Grev.) T. Moore [J]. Ind Fern J, 2011, 28(1/2): 172–178.
[12] OUYANG K T, REN H, XU Z H, et al. Habitat characteristics and population structure of, a relict plant in China [J]. Appl Ecol Environ Res, 2021, 19(3): 1939–1951. doi: 10.15666/aeer/ 1903_19391951.
[13] NOBLIN X, ROJAS N O, WESTBROOK J, et al. The fern sporran- gium: A unique catapult [J]. Science, 2012, 335(6074): 1322. doi: 10. 1126/science.1215985.
[14] GUO Y D, CAO J G, DAI X L, et al. Gametophyte development and apogamy of the fern[J]. Bull Bot Res, 2014, 34 (3): 322–327. doi: 10.7525/j.iissn.1673–5102.2014.03.006.
郭严冬, 曹建国, 戴锡玲, 等. 舟山碎米蕨配子体发育及无配子生殖[J]. 植物研究, 2014, 34(3): 322–327. doi: 10.7525/j.iissn.1673– 5102.2014.03.006.
[15] NAYAR B K, KAUR S. Gametophytes of homosporous ferns [J]. Bot Rev, 1971, 37(3): 295. doi: 10.1007/BF02859157.
[16] SHEN J, LI X G, DAI X L, et al. The effect of temperature and spore sterilization on the gametophyte development ofHoltt [J]. J Shanghai Norm Univ (Nat Sci), 2009, 38(3): 293–296. doi: 10.3969/j.issn.1000-5137.2009.03.012.
沈建, 李新国, 戴锡玲, 等. 温度及孢子消毒对黄腺羽蕨配子体发育的影响 [J]. 上海师范大学学报(自然科学版), 2009, 38(3): 293– 296. doi: 10.3969/j.issn.1000-5137.2009.03.012.
[17] WEI X P, ZHANG X C. Distributional patterns of the monolete and trilete ferns in China [J]. Biodiv Sci, 2016, 24(10): 1129–1134. doi: 10. 17520/biods.2016219.
魏雪苹, 张宪春. 蕨类植物不同孢子裂缝类型在中国的分布格局 [J]. 生物多样性, 2016, 24(10): 1129–1134. doi: 10.17520/biods.2016219.
[18] LLOYD R M, KLEKOWSKI J R E J. Spore germination and viability in pteridophyta: Evolutionary significance of chlorophyllous spores [J]. Biotropica, 1970, 2(2): 129–137. doi: 10.2307/2989770.
[19] DYER A F. The Experimental Biology of Ferns [M]. London: Aca- demic Press, 1979: 253–305.
[20] BALLESTEROS D, ESTRELLES E, WALTERS C, et al. Effect of storage temperature on green spore longevity for the fernsand[J]. Cryo Lett, 2011, 32(2): 89–98.
[21] MAGRINI S, SCOPPOLA A. First results from conservation studies of chlorophyllous spores of the Royal fern (, Osmun- daceae) [J]. Cryobiology, 2012, 64(1): 65–69. doi: 10.1016/j.cryobiol. 2011.10.001.
[22] LÓPEZ-POZO M, BALLESTEROS D, LAZA J M, et al. Desiccation tolerance in chlorophyllous fern spores: Are ecophysiological features related to environmental conditions? [J]. Front Plant Sci, 2019, 10: 1130. doi: 10.3389/fpls.2019.01130.
[23] ZHANG Y H, SHI Q, AI J G, et al. Tissue culture and rapid propa- gation of(Thunb.) Ching [J]. Plant Physiol Commun, 2010, 46(5): 473–474. doi: 10.13592/j.cnki.ppj.2010.05.007.
张延恒, 施琼, 哀建国, 等. 里白的组织培养与快速繁殖 [J]. 植物生理学通讯, 2010, 46(5): 473–474. doi: 10.13592/j.cnki.ppj.2010.05.007.
[24] DAI X L, ZHOU J, CAO J G. The analysis of the primitive features offrom gametophyte [J]. Bull Biol J, 2019, 54(11): 3– 4. doi: 10.3969/j.issn.0006-3193.2019.11.002.
戴锡玲, 周洁, 曹建国. 从配子体世代再谈紫萁的原始特征[J]. 生物学通报, 2019, 54(11): 3–4. doi: 10.3969/j.issn.0006-3193.2019.11.002.
[25] JIANG N. Phylogeny of the Polypodiaceae in China [D]. Shanghai: Shanghai Normal University, 2011: 48–49. doi: 10.7666/d.y1868256.
姜楠. 中国水龙骨科植物的系统演化研究 [D]. 上海: 上海师范大学, 2011: 48–49. doi: 10.7666/d.y1868256.
[26] BANKS J A. Gametophyte development in ferns [J]. Annu Rev Plant Physiol Plant Mol Biol, 1999, 50: 163–186. doi: 10.1146/annurev.ar plant.50.1.163.
[27] LÓPEZ-POZO M, FERNÁNDEZ-MARÍN B, GARCÍA-PLAZAOLA J I, et al. Desiccation tolerance in ferns: From the unicellular spore to the multi-tissular sporophyte [M]// FERNÁNDEZ H. Current Advances in Fern Research. Cham: Springer, 2018: 401–426. doi: 10.1007/978- 3-319-75103-0_19.
[28] SHAMBLIN S L, TANG X L, CHANG L Q, et al. Characterization of the time scales of molecular motion in pharmaceutically important glasses [J]. J Phys Chem B, 1999, 103(20): 4113–4121. doi: 10.1021/jp 983964+.
[29] BALLESTEROS D, HILL L M, WALTERS C. Variation of desiccation tolerance and longevity in fern spores [J]. J Plant Physiol, 2017, 211: 53–62. doi: 10.1016/j.jplph.2017.01.003.
[30] WALTERS C. Orthodoxy, recalcitrance and in-between: Describing variation in seed storage characteristics using threshold responses to water loss [J]. Planta, 2015, 242(2): 397–406. doi: 10.1007/s00425- 015-2312-6.
[31] BUITINK J, LEPRINCE O, HEMMINGA M A, et al. Molecular mobility in the cytoplasm: An approach to describe and predict lifespan of dry germplasm [J]. Proc Natl Acad Sci USA, 2000, 97(5): 2385– 2390. doi: 10.1073/pnas.040554797.
[32] ROUSSENOVA M, MURITH M, ALAM A, et al. Plasticization, antiplasticization, and molecular packing in amorphous carbohydrate- glycerol matrices [J]. Biomacromolecules, 2010, 11(12): 3237–3247. doi: 10.1021/bm1005068.
[33] FUNDO J F, QUINTAS M A C, SILVA C L M. Molecular dynamics and structure in physical properties and stability of food systems [J]. Food Eng Rev, 2015, 7(4): 384–392. doi: 10.1007/s12393-015-9109-z.
[34] WATKINS JR J E, CARDELÚS C L. Ferns in an angiosperm world: Cretaceous radiation into the epiphytic niche and diversification on the forest floor [J]. Int J Plant Sci, 2012, 173(6): 695–710. doi: 10.1086/ 665974.
[35] WATKINS JR J E, MACK M C, SINCLAIR T R, et al. Ecological and evolutionary consequences of desiccation tolerance in tropical fern gametophytes [J]. New Phytol, 2007, 176(3): 708–717. doi: 10.1111/j. 1469-8137.2007.02194.x.
[36] DASSLER C L, FARRAR D R. Significance of gametophyte form in long-distance colonization by tropical, epiphytic ferns [J]. Brittonia, 2001, 53(2): 352–369. doi: 10.1007/BF02812705.
[37] PITTERMANN J, BRODERSEN C, WATKINS JR J E. The physio- logical resilience of fern sporophytes and gametophytes: Advances in water relations offer new insights into an old lineage [J]. Front Plant Sci, 2013, 4: 285. doi: 10.3389/fpls.2013.00285.
Effects of Environmental Factors on Spore Germination of
OUYANG Kangting1,2, PANG Jinhui2, YU Xiaoling3, XU Zhanhui1,2, REN Hai1,2*, WANG Faguo1,2*, MA Guohua1, LIU Zongjun4, HU Mingfeng4, ZHANG Yajian4, TAN Guangwen5
(1. Guangdong Provincial Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences,Guangzhou 510650, China; 2. University of Chinese Academy of Sciences,Beijing 100049, China; 3. Zhongkai University of Agriculture and Engineering,Guangzhou 510650, China; 4. Guangdong Nanling National Nature Reserve Administration,Shaoguan 512727, Guangdong, China; 5. Pubang Landscape Architecture Co., Ltd,Guangzhou 510600, China)
To understand the endangered reasons of, a rare and endangered plant, the effects of temperature and humidity on spore germination was studied. The results showed that the spore germination ability was strong, with-type germination mode and-type gametophyte germination. The spore propagation ofwas not affected by temperature, while significantly influenced by humidity. The spores germinated normally in humid condition, forming heart-type gametophytes, and producing young sporophytes successfully. However, spores could not germinate under drought condition, gametophytes were inclined to irregularly development and folds increased. Therefore, the obstacles in spore reproduction ofunder unsuitable environment might be one of the reasons leading to the decline of wild population, so thatandconservation of these rare and endangered plants should pay attention to creating moist habitats.
; Environmental factor; Tissue culture; Gametophyte; Morphological development
10.11926/jtsb.4422
2021-04-08
2021-07-05
广东省科技计划项目(2019B121202006); 广东省野生动植物保护管理项目资助
This work was supported by the Project for Science and Technology in Guangdong (Grant No. 2019B121202006), andthe Project for Wildlife Protection and Management in Guangdong.
欧阳康婷,女,硕士研究生,主要从事园林种质资源开发与利用研究。E-mail: 865970586@qq.com
. E-mail: renhai@scbg.ac.cn, wangfg@scbg.ac.cn
