乙烯诱导水仙成花相关代谢物和基因的筛选与分析
2022-04-14何炎森李瑞美李和平
何炎森, 李瑞美, 李和平
乙烯诱导水仙成花相关代谢物和基因的筛选与分析
何炎森, 李瑞美, 李和平
(福建省农业科学院亚热带农业研究所,福建 漳州 363005)
为了解乙烯诱导水仙(var.)成花的生理和分子机制,利用代谢组和转录组测序技术,筛选乙烯诱导水仙成花的差异表达代谢物和基因。结果表明,乙烯处理的侧芽检测到12个差异表达代谢物(DEM),包括7个上调,5个下调,其中,(±)7-表茉莉酸、多巴胺、亚精胺可能与乙烯诱导水仙成花正相关,而吲哚及其衍生物呈负相关。转录组共获得1 021个差异表达基因(DEG),包括615个上调,406个下调,在DEG中鉴定筛选了45个与乙烯信号传导和开花相关的差异表达基因。乙烯诱导水仙成花启动可能先激活水仙鳞茎内源植物激素(尤其乙烯)信号通路的变化,与开花促进基因和的上调表达密切相关。9个基因的qRT-PCR结果验证了RNA-Seq的正确性。这些差异表达的代谢物和基因在水仙成花启动过程中可能具有重要作用。
水仙花;乙烯;成花诱导;转录组;代谢组;基因表达
水仙花(var.)为石蒜科(Amaryllidaceae)水仙属多年生草本球根花卉,是中国传统名花,极具观赏价值。水仙花雕刻品是以每粒鳞茎的花枝数来分等级,花枝数越多,等级越高,每支花的单价也越高[1]。为了提高经济效益, 市面上销售的水仙鳞茎球基本上全部经过乙烯利熏蒸催花处理。外源乙烯(ethylene, C2H4)对许多观赏植物和果树的开花有明显的促进作用[2],已被广泛应用于农业生产实践中。外源乙烯诱导水仙成花的研究主要在乙烯熏蒸处理时间、浓度、环境条件和效果等方面[3–4],相关机理研究较少。申艳红等用外源乙烯处理整个鳞茎球,提高了可溶性糖、蛋白质、吲哚乙酸、玉米素含量和过氧化物酶活性,筛选到31个成花相关基因[5]。外源乙烯诱导水仙成花的生理基础和分子机制的研究还亟待发展完善。
中国水仙必须经历‘芽仔’、‘钻仔’和‘种仔’阶段,栽培3 a后才成为商品球,自然条件下不能成花的水仙鳞茎母球内部最外侧芽可以被外源乙烯诱导成花。在生产实践上,花农判定水仙催花处理成败在于鳞茎母球内部的最外侧芽是否被诱导成花。本研究以中国水仙‘金盏银台’鳞茎母球内部最外侧芽为研究对象,利用高通量转录组和广泛靶向代谢组测序技术,从生理代谢和分子层面分析外源乙烯处理对代谢物及基因表达的影响,筛选乙烯诱导水仙成花的关键代谢物和基因,为乙烯诱导水仙成花机理的深入研究提供参考。
1 材料和方法
1.1 材料
中国水仙(var.)‘金盏银台’鳞茎球来源于福建省农业科学院亚热带农业研究所试验基地。同批次“种仔”阉割后种植在相同田块,从收获的3 a生鳞茎球中,选择健康、排花状、主鳞茎围径(21±0.5) cm的鳞茎球共200粒,随机分成2组各100粒,装于竹框内。一组放到花农仓库与3万多粒水仙花球同时进行乙烯利熏蒸处理[分别于7月23和26日各处理1次,便携式乙烯报警仪(PGD3-C-C2H4)测得处理过程仓库内乙烯气体质量浓度为1 250~2 500 mg/m3],然后自然室温贮藏;另一组无乙烯处理,自然室温贮藏为对照。在催花处理后(7月27日),对水仙鳞茎球进行解剖取样,鳞茎母球内部最外侧芽的样品包含芽点和部分鳞茎盘,取样部位如图1。每个处理3个重复,同时取样2份。有无乙烯处理的最外侧芽(lateral bud)分别标记为EL和L。所有样品液氮速冻后装入塑料袋内,存于–80 ℃冰箱备用。

图1 水仙鳞茎母球内部最外侧芽取样部位
1.2 石蜡切片观察
7月15和30日分别取最外侧芽,分别标记为L0715、L0730和EL0730。参考方舟[6]的方法制作石蜡切片, 对顶端生长点组织进行纵切面切片和电镜观察。
1.3 转录组、代谢组测序及数据分析
委托广东基迪奥生物技术有限公司进行转录组测序和液相色谱串联质谱分析代谢物。运用生物信息学技术和方法,对测序数据进行前期处理、序列比对、基因及基因组的注释、基因表达分析等。运用Analyst1.6.3及Mutiaquant软件分析代谢物质谱数据,利用广东基迪奥生物技术有限公司自建的代谢物数据库定性代谢物。以错误发现率(false discovery rate, FDR)<0.05且两样品(组)间表达量的比值(fold change, FC)以2为底的对数值的绝对值(|log2FC|)>1为标准筛选转录组的差异表达基因(differrentially expressed gene, DEG),以OPLS-DA中VIP≥1且|log2FC|>0.5为标准筛选代谢组的差异表达代谢物(differentially expressed metabolites, DEM)。
1.4 qRT-PCR分析
选择9个侧芽差异表达基因(表1),以基因(Unigene0067272)作为内参,采用qRT-PCR检测基因表达水平,3次生物学重复,这些差异基因的表达谱与RNA-Seq结果进行比较分析。
2 结果和分析
2.1 乙烯处理对水仙成花的影响
水仙花芽分化时间与品种特性有关,也受外界环境气候条件影响。漳州花农通常在7月15—30日(大暑节气前后1周)对水仙进行催花处理。对水仙鳞茎母球内部最外侧芽纵切面观察(图2),7月15日,最外侧芽(L0715)的顶端分生组织呈圆丘状,处于营养生长时期;7月30日,没有经过乙烯处理的最外侧芽(L0730)仍处于营养生长时期,而经过乙烯处理的最外侧芽(EL0730)的顶端分生组织已经形成凹凸面,进入花序原基形成期,这是营养生长向生殖生长转化的标志[7], 说明外源乙烯可诱导水仙鳞茎母球内部最外侧芽的成花启动。

表1 qRT-PCR分析的9个基因和引物
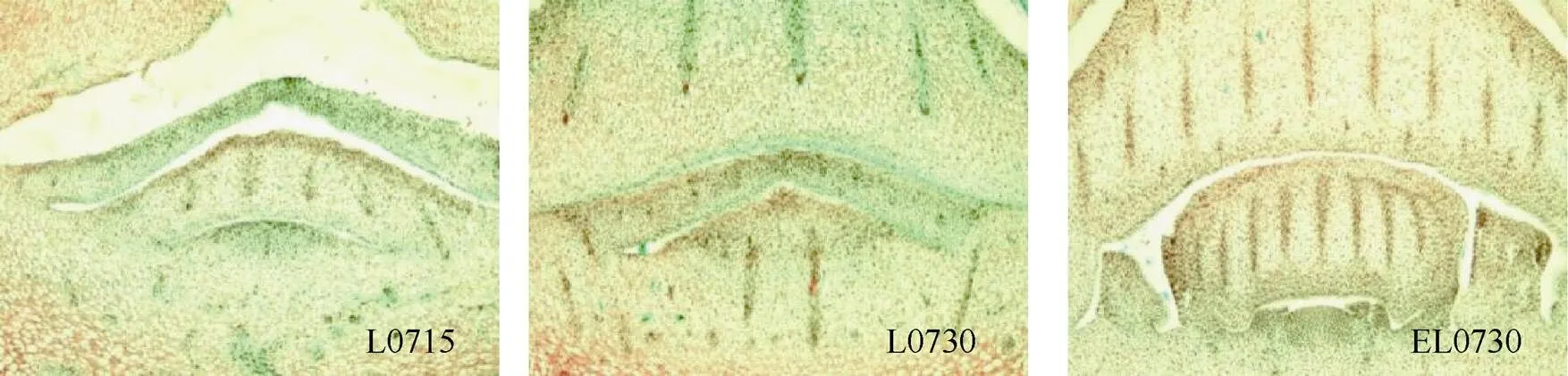
图2 水仙鳞茎母球内部最外侧芽纵切面。L0715: 7月15日无乙烯处理; L0730: 7月30日无乙烯处理,EL0730: 7月30日乙烯处理。
2.2 代谢组分析
根据公共数据库与基迪奥生物公司自建数据库对代谢物进行定性和鉴定,得到184个代谢物,包括99个已知和85个未知。已知代谢物主要包含16个羧酸和衍生物(carboxylic acid and derivative)、14个脂肪酰基(fatty acyl)、13个苯和取代衍生物(benzene and substituted derivative)、11个有机氧化合物(organooxygen compound)、7个小肽(mini pep- tide)、4个孕烯醇酮脂类(prenol lipid)、3个吲哚类和衍生品(indoles and derivative)、3个有机氮化合物(organonitrogen compound)、3个菲类和衍生品(phe- nanthrene and derivative)、3个嘧啶核苷酸(pyrimi- dine nucleotides)等。共有62个代谢物被注释到63代谢通路中,主要富集在氨基酸代谢、碳水化合物代谢、脂质代谢、其他次生代谢物生物合成和膜转运。
LvsEL组间共检测到12个DEM (表2),其中, (±)7-表茉莉酸、多巴胺、十八碳三烯酸、前列腺素、甘油磷酰胆碱、13-l-过氧氢亚油酸、亚精胺等7个代谢物上调表达;苯丙氨酸、吲哚及其衍生物等4个代谢物下调表达。有5个DEM注释到15个代谢通路, 主要富集在甜菜素生物合成、甘油磷脂新陈代谢、色氨酸的生物合成、亚油酸的新陈代谢、苯丙素的生物合成等通道。LvsEL组间,(±)7-表茉莉酸上调表达(FC 为21.93),乙烯处理的最外侧芽(EL)中的含量远大于无乙烯处理的最外侧芽(L);多巴胺上调表达(FC为1.49),4个吲哚及其衍生物表达下调(FC为–0.79~ 0.86);亚精胺(FC为0.65)上调表达。

表2 差异代谢物
2.3 转录组分析
测序数据经质量控制和序列拼接组装,共获得116 715条Unigenes,平均序列长度为776 bp,GC含量41.89%,N50数量为20 219,N50长度1 219 bp,说明组装质量好。利用数据库对Unigenes进行注释,在Nr数据库注释到49 753个,在KEGG数据库注释到43 936个,在COG数据库注释到25 014个,在Swissprot数据库注释到26 661个,共注释到了49 949个Unigenes,占42.80%。
LvsEL差异表达基因在GO数据库中得到注释的共有1 021个Unigenes, 其中, 615个上调, 406个下调。对这些基因进行GO功能分类,可以分成3大类:细胞组分(cellular component)、分子功能(mole- cular function)和生物过程(biological process) (表3)。在细胞组分中,差异基因参与最多的是细胞(cell)、细胞区域(cell part)和细胞膜(membrane),分别有39、39和38个,占差异基因数的62.9%、62.9%和61.29%。在分子功能中,差异基因参与最多的是催化活动(catalytic activity)、蛋白结合(binding)和转运活性(transporter activity),分别有93、55和9个,分别占差异基因数的77.5%、45.83%和7.5%。生物过程方面,差异基因主要集中在代谢过程(metabolic process)、单生物过程(single-organism process)和细胞过程(cellular process),分别有90、75和71个, 分别占差异基因数的75.63%、63.03%和59.66%。
差异基因的KEGG通路富集分析表明,LvsEL有170个基因在KEGG数据库中得到注释,共注释到77个通路,其中,参与代谢大类(metabolic path-way)、次生代谢物生物合成(Biosynthesis of secon- dary metabolite)的差异基因最多,分别有73个(占42.94%)和43个(占25.29%)。Q<0.05的有6条通路(表4),分别为植物激素信号转导(plant hormone signal transduction)、植物MAPK信号通路(MAPK signaling pathway-plant)、植物-病原互作(plant- pathogen interaction)、类胡萝卜素生物合成(carote- noid biosynthesis)、亚油酸的新陈代谢(linoleic acid metabolism)、植物氮代谢(nitrogen metabolism)。其中,在植物激素信号转导通路中的差异基因有25个(占14.71%);在植物MAPK信号通路和植物-病原互作通路中各有20个(各占11.76%);在类胡萝卜素生物合成和亚油酸的新陈代谢通路中各有5个(各占2.94%);在植物氮代谢通路有6个(占3.53%)。此外,在碳代谢(carbon metabolism)和氨基酸合成(biosynthesis of amino acids)通路中各有8个(各占4.71%), 在氨基糖和核苷酸糖代谢通路(amino sugarand nucleotide sugar metabolism)中有7个(占4.12%)。可见,LvsEL差异基因显著性富集在植物激素信号转导通路、植物MAPK信号通路中。
在没有基因组信息和缺乏参考序列的情况下,根据转录组数据注释结果,获得开花相关的Unigenes有99个(未列出)。从差异表达基因中鉴定和筛选了45个可能与乙烯信号传导和开花相关的差异表达基因(表5),包括植物激素信号传导相关基因25个,包括与乙烯相关的和,与赤霉素相关的,与脱落酸相关的、;与生长素相关的、、,与细胞分裂素相关的,与茉莉酸相关的;植物MAPK信号通路相关基因4个,包括、、和;激素生物合成相关基因7个,包括乙烯合成的,脱落酸合成的,油菜素类固醇合成的和,细胞分裂素合成的和,赤霉素合成的等;氨基酸、淀粉和糖代谢相关3个,包括、和;开花相关的差异基因6个,包括、、、、和。这些DEG有的可能仅仅参与乙烯的基本反应,有的直接或间接地参与了水仙的成花诱导和开花调控。

表3 LvsEL差异基因GO功能分类

表4 LvsEL差异基因KO富集的主要通路

表5 乙烯诱导水仙成花相关的差异基因
为验证转录组测序的准确性和可重复性,我们选择LvsEL中9个差异基因进行qRT-PCR,除Uni- gene0064815基因序列的GC含量稍高,导致熔解曲线左侧有小峰外,其余基因的扩增曲线和溶解曲线都正常,内参基因在不同样本中的ct值较稳定。验证的9个基因中,除Unigene0071640外,其余8个基因在qPCR比较组中的表达趋势与转录组中表达趋势一致(图3),qPCR验证率接近90%以上, 说明RNA-Seq结果是可靠的。
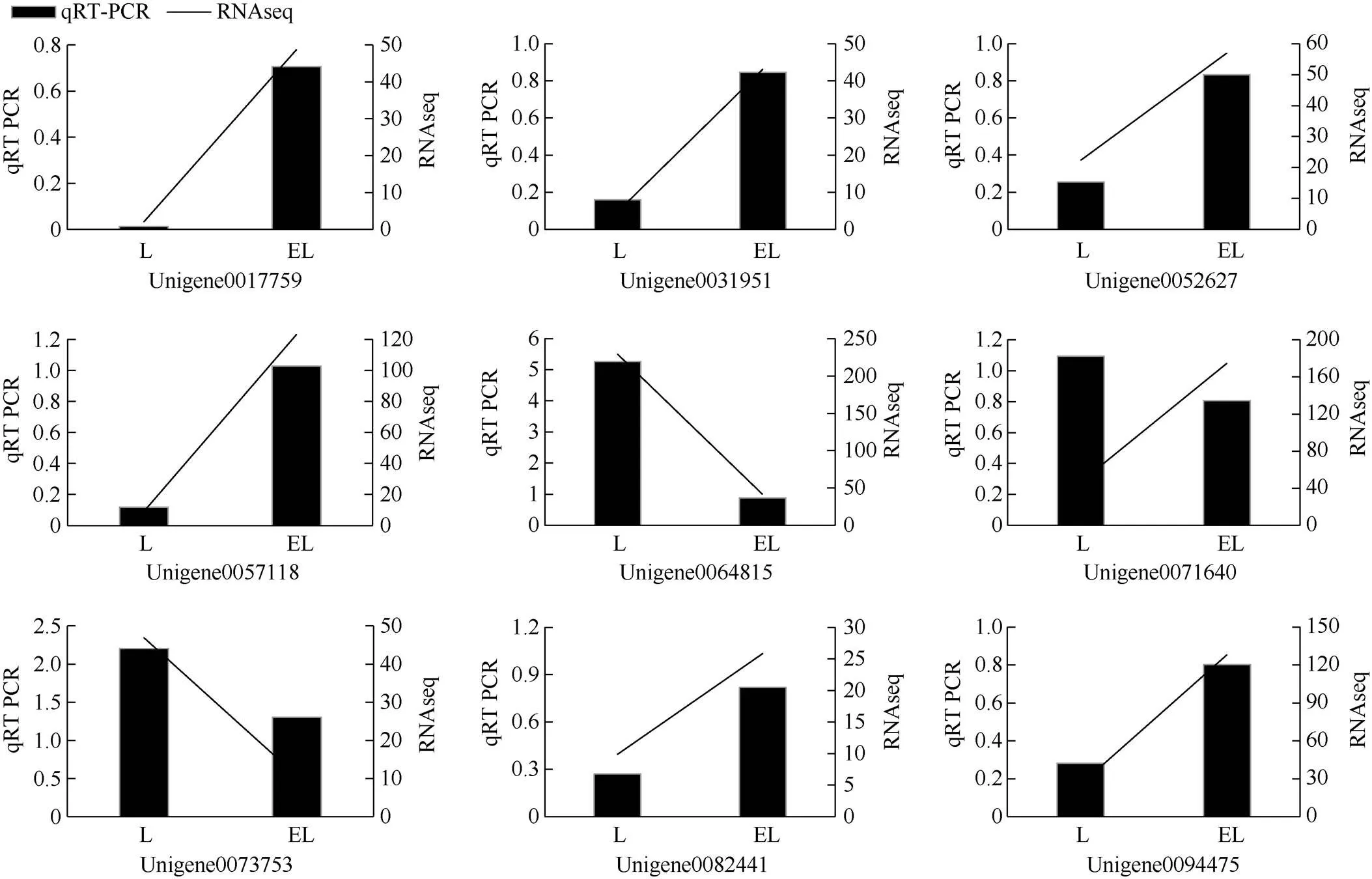
图3 qRT-PCR和RNA-Seq分析部分差异表达基因的相对表达量。L: 无C2H4处理; EL: C2H4处理。
3 结论和讨论
营养物质和内源激素含量在成花诱导和形成中起着重要作用[8]。代谢组分析表明,外源乙烯处理使12个代谢物出现差异表达,其中,7个代谢物上调表达,5个下调表达。外源乙烯处理显著促进了水仙外侧芽中(±)7-表茉莉酸的合成。已有研究报道,内源茉莉酸参与紫萍()的开花调控[9],茉莉酸类物质在水稻()颖花诱导和发育中起着重要作用[10]。外源乙烯处理也促进了多巴胺和亚精胺的合成。多巴胺是能诱导产生更多的花原基和显著促进花的发育[11],亚精胺可诱导烟草薄层外植体的营养芽成花[12]。同时,外源乙烯处理显著地促进十八碳三烯酸、前列腺素、甘油磷酰胆碱、亚油酸等营养活性物质的上调表达。这4种代谢物都是细胞膜的主要成分,还参与细胞膜对蛋白质的识别和信号传导。此外,外源乙烯处理显著地降低了吲哚及其衍生物的水平。外源乙烯处理降低内源激素吲哚-3-乙酸(IAA)的水平,有利于菠萝()的花芽分化[13],由吲哚类化合物组成的生长素(auxin)参与植物生长和发育诸多过程[14]。外源乙烯诱导水仙成花可能与这些内源激素和营养物质的差异表达有关。转录组分析还表明,外源乙烯处理,导致7个内源激素生物合成相关的酶基因(、、、、、和)以及3个营养物质合成酶基因(和)出现表达差异,可以进一步利用靶向代谢组技术进行准确定量和分析。
从石蜡切片观察到,外源乙烯处理可诱导水仙鳞茎母球内部自然不能成花的最外侧芽从营养生长向生殖生长转变,佐证了常规花农催花的科学性。LvsEL差异基因KEGG通路富集分析表明,外源乙烯处理,差异表达基因主要富集在植物激素信号转导通路,涉及乙烯、赤霉素、生长素、细胞分裂素、脱落酸、茉莉酸等共25个基因。乙烯、赤霉素、生长素、脱落酸、茉莉酸等植物激素参与开花调控[15],说明外源乙烯通过激活水仙鳞茎内部信号通路,从而调控水仙最外侧芽从营养生长向生殖生长转变。乙烯是植物的5大类内源激素之一, 外源乙烯处理引起植物内源乙烯含量的增加[16],内源乙烯参与拟南芥()、凤梨()从营养生长到生殖生长的过程[17–18]。本研究转录组分析表明,外源乙烯处理,乙烯生物合成关键酶基因上调表达,这与前人[19]对拟南芥、蝴蝶兰()、青花菜()的研究结果一致。据报道,水仙过表达转基因烟草()与野生型相比缩短了营养生长期,开花较早[20]。同时, 外源乙烯处理,2个基因上调表达。在乙烯信号通路中起决定性作用,也承担了乙烯和其他信号交流互作的任务[21]。据报道,水仙基因具有正调控乙烯响应的功能[22],可能在乙烯诱导凤梨开花中起重要作用[23]。乙烯与生长素、光、赤霉素、茉莉酸、水杨酸、细胞分裂素、葡萄糖等途径存在着广泛的交叉反应,共同参与了调控植物的生长发育和应对生物、非生物胁迫等过程[24]。本研究由于检测方法的原因没能检测到乙烯成分, 但水仙最外侧芽能够成花最终是受到乙烯所调控, 因此推测,外源乙烯诱导水仙成花可能与内源乙烯信号传导关联密切。
模式植物开花有光周期、温敏、春化、自主、赤霉素、抑制和年龄等7个遗传通路参与调控[25]。本研究结果表明,外源乙烯处理影响和等6个开花相关基因的表达,其中涉及赤霉素途径的开花抑制基因下调表达,DELLA蛋白降解可以促进拟南芥开花[26];涉及抑制途径的开花抑制基因上调表达。据报道,在营养生长阶段,基因低水平表达会延迟开花的转变,但在花序分生组织(IM)中,基因高水平表达可以维持IM的特性[27]; 涉及光周期途径的上调表达,属于AP2/ ERF转录调控家族中的亚族, 可能调节光敏色素(PHY)的表达[28],参与花器官发育的起始和生长[29];涉及春化途径的春化独立蛋白基因上调表达,在花序分生组织中高表达,有利于花序茎的数量增加和簇生[30]。总体来说,外源乙烯处理能影响一些开花途径相关基因的表达,但在这个阶段并未表达,而开花整合子基因(Unigene0007988, FC为–0.06; Unigene 0013465, FC为–0.457)(Unigene0050424,FC为0.425)(20个Unigenes注释为基因,其中10个上调表达、10个下调表达)在处理间并没有显著差异。因此推测,乙烯诱导水仙成花启动可能并不通过已知开花调控途径中的任何一条。这与丛汉卿[31]对乙烯诱导凤梨开花的研究结果基本相同。
值得重点关注的是,本研究结果表明,外源乙烯处理导致1个开花促进基因和1个开花识别基因的大量上调表达。据报道,参与赤霉素信号途径中调控开花,组成性表达导致拟南芥幼年期缩短提前开花[32],基因可能通过调节顶端分生组织对开花识别基因的反应能力来控制开花时间[33]。拟南芥、水稻、烟草、和棉花(spp.)等多种植物的研究表明正调控开花时间[34],但目前其调控开花时间的信号途径并不清楚。而水仙的与拟南芥的为同源基因,是开花启动的标志基因, 处于成花调控网络的关键位置[35]。过量表达使植物呈现早熟表型和提前开花[36],推测乙烯诱导水仙成花启动可能与基因和基因的上调表达密切相关。
综上所述,本研究结果表明外源乙烯可以诱导3 a生水仙鳞茎母球内部最外侧芽成花启动,佐证了花农常规以最外侧芽是否被诱导成花来判断催花成败的科学性。利用代谢组和转录组技术,探讨了外源乙烯促进水仙成花的原因。代谢组分析表明有12个代谢物出现差异表达,其中,(±)7-表茉莉酸、多巴胺、亚精胺可能与乙烯诱导水仙成花正相关,而吲哚及其衍生物呈负相关。转录组鉴定和筛选了45个与乙烯信号传导和开花相关的差异表达基因,乙烯诱导水仙成花启动可能先激活水仙鳞茎内源植物激素(尤其乙烯)信号通路的变化,与开花促进基因和的上调表达密切相关。这些差异代谢物和差异基因在外源乙烯诱导水仙成花启动的具体作用仍需进一步研究和验证。
[1] YE J B. A study on mechanisms and application ofvar.Roem’s flower forcing technique [J]. Chin Hort Abstr, 2009, 25(7): 26–28. doi: 10.3969/j.issn.1672-0873.2009.07.012.
叶季波. 中国水仙鳞茎催花技术机理及应用研究[J]. 中国园艺文摘, 2009, 25(7): 26–28. doi: 10.3969/j.issn.1672-0873.2009.07.012.
[2] Chen X L, Lu M Y. Progress of studies on application of ethrel in plant flower-formation regulation [J]. Guangxi Agric Sci, 2005, 36(2): 110–112. doi: 10.3969/j.issn.2095-1191.2005.02.008.
陈香玲, 卢美英. 乙烯利在植物成花方面的应用及研究进展[J]. 广西农业科学, 2005, 36(2): 110–112. doi: 10.3969/j.issn.2095-1191. 2005.02.008.
[3] ZHANG J D. Study on chemical regulation ofblooms [J]. J Zhangzhou Teach Coll, 1995, 9(4): 112–112.
章骏德. 水仙化控的增花研究[J]. 漳州师范学院学报(哲学社会科学版), 1995, 9(4): 112–115.
[4] IMANISHI H. Effects of exposure of bulbs to smoke and ethylene on flowering ofcultivar ‘Grand Soleil d’Or’ [J]. Sci Hort, 1983, 21(2): 173–180.
[5] SHEN Y H, JIANG T, ZHAO W W, et al. Study on technology and mechanism of ethylene treatment promotes the formation of more flowers ofvar.[J]. J Agric Biotechnol, 2019, 27(6): 1003–1015. doi: 10.3969/j.issn.1674-7968.2019.06.006.
申艳红, 姜涛, 赵湾湾, 等. 乙烯处理水仙催多花技术和机理的研究[J]. 农业生物技术学报, 2019, 27(6): 1003–1015. doi: 10.3969/j. issn.1674-7968.2019.06.006.
[6] FANG Z. Morphological study on flower bud differentiation ofL. var.Roen [D]. Fuzhou: Fujian Agriculture and Forestry University, 2017: 11–13.
方舟. 水仙‘金玉’花芽分化研究[D]. 福州: 福建农林大学, 2017: 11–13.
[7] ZHANG X Q, GAO J, PENG Z H. Morphological observation on flower bud differentiation and influences of storage conditions on the quantity of flower buds ofvar.[J]. Bull Bot Res, 2012, 32(5): 549–553.
张晓晴, 高健, 彭镇华. 中国水仙花芽分化观察及储藏条件对花芽数的影响研究[J]. 植物研究, 2012, 32(5): 549–553.
[8] LI X L, YUAN Z Y, GAO D S. The research achievements on mecha- nism of floral formation in plants [J]. Acta Bot Boreali-Occid Sin, 2002, 22(1): 173–183. doi: 10.3321/j.issn:1000-4025.2002.01.031.
李宪利, 袁志友, 高东升. 高等植物成花分子机理研究现状及展望[J]. 西北植物学报, 2002, 22(1): 173–183. doi: 10.3321/j.issn:1000- 4025.2002.01.031.
[9] KRAJNČIČ B, NEMEC J. The effect of jasmonic acid on flowering in(L.) Schleiden [J]. J Plant Physiol, 1995, 146(5/6): 754–756. doi: 10.1016/S0176-1617(11)81945-0.
[10] HUANG Y M, ZENG X C. Induction effect of jasmonic acid (JA) related compounds on floret opening in rice [J]. Hubei Agric Sci, 2008, 47(10): 1125–1127. doi: 10.3969/j.issn.0439-8114.2008.10.008.
黄友明, 曾晓春. 茉莉酸相关化合物对水稻颖花开放的诱导效应[J]. 湖北农业科学, 2008, 47(10): 1125–1127. doi: 10.3969/j.issn.0439- 8114.2008.10.008.
[11] KHURANA J P, TAMOT B K, MAHESHWARI N, et al. Role of cate- cholamines in promotion of flowering in a short-day duckweed,6746 [J]. Plant Physiol, 1987, 85(1): 10–12.
[12] KAUR-SAWHNEY R, TIBURCIO A F, GALSTON A W. Spermidine and flower-bud differentiation in thin-layer explants of tobacco [J]. Planta, 1988, 173(2): 282–284. doi: 10.1007/BF00403022.
[13] LIU S H, ZANG X P, ZHANG X M, et al. Changes of endogenous hormone levels during the inflorescence differentiation in pineapple (Smooth Cayenne. cv) [J]. Chin J Trop Crops, 2010, 31(9): 1487–1492. doi: 10.3969/j.issn.1000-2561.2010.09.010.
刘胜辉, 臧小平, 张秀梅, 等. 乙烯利诱导菠萝[(L.) Merril]花芽分化过程与内源激素的关系[J]. 热带作物学报, 2010, 31(9): 1487–1492. doi: 10.3969/j.issn.1000-2561.2010.09.010.
[14] ZHANG J. Review on Auxin signal transduction pathway and biolo- gical functions [J]. Life Sci Res, 2009, 13(3): 272–277. doi: 10.16605/ j.cnki.1007-7847.2009.03.011.
张娟. 生长素信号转导途径及参与的生物学功能研究进展[J]. 生命科学研究, 2009, 13(3): 272–277. doi: 10.16605/j.cnki.1007-7847. 2009.03.011.
[15] ZHOU L P, PAN C, WANG M X, et al. Progress on the mechanism of hormones regulating plant flower formation [J]. Hereditas, 2020, 42(8): 739–751. doi: 10.16288/j.yczz.20-014.
邹礼平, 潘铖, 王梦馨, 等. 激素调控植物成花机理研究进展[J]. 遗传, 2020, 42(8): 739–751. doi: 10.16288/j.yczz.20-014.
[16] KE D S, WANG A G, LUO G H. The effect of activated oxygen during the production of endogenous ethylene induced by exogenous ethylene [J]. Acta Phytophysiol Sin, 1997, 23(1): 67–72.
柯德森, 王爱国, 罗广华. 活性氧在外源乙烯诱导内源乙烯产生过程中的作用[J]. 植物生理学报, 1997, 23(1): 67–72.
[17] OGAWARA T, HIGASHI K, KAMADA H, et al. Ethylene advances the transition from vegetative growth to flowering in[J]. J Plant Physiol, 2003, 160(11): 1335–1340. doi: 10.1078/ 0176-1617-01129.
[18] DUKOVSKI D, BERNATZKY R, HAN S S. Flowering induction ofby ethylene [J]. Sci Hort, 2006, 110(1): 104–108. doi: 10. 1016/j.scienta.2006.05.004.
[19] POGSON B J, DOWNS C G, DAVIES K M. Differential expression of two 1-aminocyclopropane-1-carboxylic acid oxidase genes in broccoli after harvest [J]. Plant Physiol, 1995, 108(2): 651–657. doi: 10.1104/pp. 108.2.651.
[20] SUN S S, WEN X P, YANG F Y, et al. Cloning of ACC oxidase gene fromvar. ‘Yunxiang’ and its transformation [J]. Acta Hort Sin, 2017, 44(7): 1388–1396. doi: 10.16420/j.issn.0513-353x.2016- 0884.
孙申申, 温秀萍, 杨菲颖, 等. ‘云香’水仙ACC氧化酶基因克隆及遗传转化[J]. 园艺学报, 2017, 44(7): 1388–1396. doi: 10.16420/j. issn.0513-353x.2016-0884.
[21] DOLGIKH V A, PUKHOVAYA E M, ZEMLYANSKAYA E V. Shaping ethylene response: The role of EIN3/EIL1 transcription factors [J]. Front Plant Sci, 2019, 10: 1030. doi: 10.3389/fpls.2019.01030.
[22] XU J. Functional study of NFT1 and NEIN3 genes related to dormancy regulation invar.[D]. Shanghai: East China Normal University, 2013: 63–65.
许静. 中国水仙休眠调控相关基因和的功能研究[D]. 上海: 华东师范大学, 2013: 63–65.
[23] LI Z Y, ZHANG X Q, ZHANG K, et al. Cloning and expressing analysisofin[J]. Mol Plant Breed, 2015, 13(1): 139–144.
李志英, 张学全, 张鲲, 等. 蜻蜓凤梨中的克隆及其表达特性分析[J]. 分子植物育种, 2015, 13(1): 139–144.
[24] AN F Y, GUO H W. Molecular mechanism of ethylene signal trans- duction [J]. Chin Bull Bot, 2006, 23(5): 531–542. doi: 10.3969/j.issn. 1674-3466.2006.05.009.
安丰英, 郭红卫. 乙烯信号转导的分子机制[J]. 植物学通报, 2006, 23(5): 531–542. doi: 10.3969/j.issn.1674-3466.2006.05.009.
[25] SONG Y, DOU L D, ZHANG H J. Molecular and genetic mechanisms of control of floral induction in higher plants [J]. Plant Physiol J, 2014, 50(10): 1459–1468. doi: 10.13592/j.cnki.ppj.2014.0270.
宋杨, 窦连登, 张红军. 高等植物成花诱导调控的分子和遗传机制[J]. 植物生理学报, 2014, 50(10): 1459–1468. doi: 10.13592/j.cnki.ppj. 2014.0270.
[26] BAO S J, HUA C M, SHEN L S, et al. New insights into gibberellin signaling in regulating flowering in[J]. J Integr Plant Biol, 2020, 62(1): 118–131. doi: 10.1111/jipb.12892.
[27] WANG L N, LIU Q L. Phylogenetic tree and function analysis of inflorescence meristem identity gene[J]. China Biotechnol, 2008, 28(1): 106–112. doi: 10.3969/j.issn.1671-8135.2008.01.019.
王丽娜, 刘青林. 花序分生组织特性基因的系统发育及其功能分析[J]. 中国生物工程杂志, 2008, 28(1): 106–112. doi: 10.3969/j. issn.1671-8135.2008.01.019.
[28] KARLBERG A, BAKO L, BHALERAO R P. Short day-mediated cessation of growth requires the downregulation of AINTEGUMEN- TALIKE1 transcription factor in hybrid aspen [J]. PLoS Genet, 2011, 7(11): e1002361. doi: 10.1371/journal.pgen.1002361.
[29] MUDUNKOTHGE J S, KRIZEK B A. Threegenes act in combination to regulate shoot apical meristem function [J]. Plant J, 2012, 71(1): 108–121. doi: 10.1111/j.1365-313X.2012.04975.x.
[30] TAKAGI N, UEGUCHI C. Enhancement of meristem formation by, a mis-sense allele of thegene encoding a WD40 repeat protein in[J]. Genes Cells, 2012, 17(12): 982–993. doi: 10.1111/gtc.12014.
[31] CONG H Q. Expression characterization of flowering related genes after ethylene treatment inand preliminary study on molecular mechanism of flower induction [D]. Haikou: Hainan University, 2012: 87–92.
丛汉卿. 乙烯诱导蜻蜓凤梨开花相关基因表达分析及其催花分子机制的初步研究[D]. 海口: 海南大学, 2012: 87–92.
[32] KANIA T, RUSSENBERGER D, PENG S, et al. Fpf1 promotes flowering in[J]. Plant Cell, 1997, 9(8): 1327–1338. doi: 10.1105/tpc.9.8.1327.
[33] MELZER S, KAMPMANN G, CHANDLER J, et al.modulates the competence to flowering in[J]. Plant J, 1999, 18(4): 395–405. doi: 10.1046/j.1365-313X.1999.00461.x.
[34] WANG X Y, FAN S L, SONG M Z, et al. Upland cotton geneconfers promotion of flowering time and shade-avoidance responses in[J]. PLoS ONE, 2014, 9(3): e91869. doi: 10. 1371/journal.pone.0091869.
[35] MANDEL M A, GUSTAFSON-BROWN C, SAVIDGE B, et al. Molecular characterization of thefloral homeotic gene[J]. Nature, 1992, 360(6401): 273–277. doi: 10.1038/360 273a0.
[36] LU S J, WEI H, WANG Y, et al. Overexpression of a transcription factor OsMADS15 modifies plant architecture and flowering time in rice (L.) [J]. Plant Mol Biol Rep, 2012, 30(6): 1461–1469. doi: 10.1007/s11105-012-0468-9.
Screening of Metabolites and Genes Related to Floral Formation of Chinese Narcissus Induced by Ethylene
HE Yansen, LI Ruimei, LI Heping
(Institute of Subtropical Agriculture, Fujian Academy of Agricultural Sciences, Zhangzhou 363005, Fujian, China)
In order to understand the mechanism of the narcissus (var.) floral induction, the differentially expressed metabolites and genes were screened from the outermost buds treated with exogenous ethylene by using metabolome and transcriptome sequencing techniques. The results showed that 12 differentially expressed metabolites (DEMs) were detected, including 7 up-regulated and 5 down-regulated DEMs. Among them, (±) 7-epigenJasmonic acid, dopamine and spermidine might be positively correlated with narcissus floral induction, while indole and its derivatives were negatively correlated. A total of 1 021 differentially expressed genes (DEGs) were identified in the transcriptome, including 615 up-regulated and 406 down-regulated DEGs. Forty-five differentially expressed genes related to ethylene signal transduction and flowering were identified in the DEGs. The changes of endogenous plant hormone (especially ethylene) signaling pathway in narcissus bulbs were activated firstly by exogenous ethylene, and the floral induction of narcissusexogenous ethylene was closely related to the up-regulated expression ofand. Nine genes correlated with flowering were verified by qRT-PCR analysis and the expression profiles were consistent with the RNA-Seq results. Therefore, these DEMs and DEGs might have vital function on the narcissus floral induction, which might play an important role in the floral formation of narcissus induced exogenous ethylene.
; Ethylene; Floral induction; Transcriptome; Metabolite; Gene expression
10.11926/jtsb.4441
2021-05-08
2021-07-05
福建省漳州市科技重大专项(ZZ2019ZD03); 福建省公益类科研院所专项(2020R1030002)资助
This work was supported by the Major Project for Science and Technology in Zhangzhou City, Fujian (Grant No. ZZ2019ZD03); and the Special Project for Public Welfare Research Institutes in Fujian (Grant No. 2020R1030002).
何炎森(1968~ ),男,博士,副研究员,主要从事观赏植物栽培和遗传育种研究。E-mail: 1907750591@qq.com
. E-mail: 1907750591@qq.com
