苹果属PP2C A亚家族基因的克隆及生信分析
2022-04-12何钰贤姚允聪卢艳芬
许 璇,何钰贤,姚允聪,卢艳芬
(北京农学院 植物科学技术学院/农业应用新技术北京重点实验室,北京102206)
脱落酸(ABA)是一种重要的植物激素,已被证明参与植物响应环境胁迫及多种生长发育过程[1],ABA信号通路是植物特有的激素信号途径[2]。依赖于Mg2+/Mn2+的Ser/Thr蛋白磷酸酶2C(Group A-PP2C)是植物细胞内ABA信号网络的核心调控因子,其介导的ABA-PYR/PYL/RCAR-PP2Cs-SnRKs ABA信号感知与转导通路已成为ABA信号转导领域的重大突破之一。PP2C介导的蛋白质可逆磷酸化是植物体内调控细胞信号转导的重要方式[3],随着PYR1/PYLs/RCAR1 被鉴定为ABA受体[4],人们对ABA信号转导途径的分子机制有了更深入的了解。ABA通过调节PP2C 活性来调控细胞代谢,ABA受体结合PP2C并抑制PP2C的磷酸化活性,从而激活SnRK2s。SnRK2s进一步磷酸化并激活下游ABA依赖型转录因子AREB/ABF[5]。
PP2C对ABA信号的响应已成为一个研究热点,其中A亚家族PP2C更是ABA信号转导过程中的关键负调控因子[6]。在保卫细胞的细胞核中,A亚家族PP2C会抑制SnRK2激酶活性,进而抑制响应ABA信号的转录因子表达。而在胁迫条件下,A亚家族PP2C与ABA受体结合捕获ABA信号,激活SnRK2及下游基因表达[7]。在拟南芥中已鉴定出9种PP2C A亚家族基因[8],并发现ABA受体PYL5介导的A亚家族PP2C活性的抑制,可以增强植物的抗旱性[9]。在玉米[10]、鸢尾[11]等植物中也表现出相似的结果。
平邑甜茶是优秀的苹果砧木资源,为苹果属湖北海棠(MalushupehensisRehd.)的一个类型,因其无融合生殖的特殊生殖方式,使砧木具有很高的整齐度,具有适应性强、耐高温潮湿、嫁接亲和力强和单株产量高等诸多优良特性[12,被广泛应用于农业生产和基础科学研究。但是该砧木PP2C基因家族在ABA信号转导中的作用尚不明确。本研究以苹果砧木‘平邑甜茶’作为试验材料,寻找其中的重要PP2C基因。为今后探究‘平邑甜茶’中脱落酸信号转导的分子机制提供理论基础。
1 材料与方法
1.1 试验材料
苹果砧木品种‘平邑甜茶’为北京农学院组培中心保存的组培苗,培养于MS培养基(蔗糖:30 g/L、MS粉:4.43 g/L、6-BA:1 mg/L、IBA:0.2 mg/L、pH:5.85~5.90)中,培养温度为23 ℃±2 ℃,相对湿度保持在60%~70%,光照强度为1 800~2 000 lx,光照时间为16 h/d。
1.2 试验方法
1.2.1 ‘平邑甜茶’总RNA提取和cDNA反转录 ‘平邑甜茶’组培苗总RNA提取方法参照EASY spin Plus多糖多酚/复杂植物RNA快速提取试剂盒(北京艾德莱生物科技有限公司)。通过1%琼脂糖凝胶电泳检测所得总RNA质量,并用紫外分光光度计检测总RNA浓度。总RNA提取后,利用cDNA合成试剂盒(宝日医生物技术(北京)有限公司)反转录合成cDNA。
1.2.2 ‘平邑甜茶’PP2C A亚家族基因克隆 根据NCBI发布的拟南芥PP2C A亚家族基因的4个基因AtABI1(AT4G26080)、AtABI2(AT5G57050)、AtHAB1(AT1G72770)和AtHAB2(AT1G17550),利用蔷薇科基因组数据库GDR查找苹果同源基因,用Basic Local Alignment Search Tool下载完整CDS区序列,并通过Primer 5设计引物,序列见表1。以反转录得到的‘平邑甜茶’cDNA为模板,使用2×Phanta Max Master Mix(Dye Plus)高保真酶(南京诺唯赞生物科技股份有限公司)进行基因克隆,反应体系为50 μL,包含:cDNA模板2 μL;高保真酶Mix 25 μL;上、下游引物(10 μmol/L)各2 μL;ddH2O 19 μL,反应程序为:95 ℃ 3 min;95 ℃ 15 s,58 ℃ 15 s,72 ℃ 1 min,35个循环;72 ℃ 延伸5 min;4 ℃保温。利用FastPure Gel DNA Extraction Mini Kit胶回收/DNA纯化试剂盒(南京诺唯赞生物科技股份有限公司)进行PCR产物回收,连接于pEASY-Blunt Cloning Vector(北京全式金生物技术有限公司)后转化大肠杆菌DH5α感受态(上海唯地生物技术有限公司),挑菌,进行菌液PCR鉴定后,选取阳性克隆进行测序。

表1 引物序列Tab.1 Primer sequences
1.2.3 ‘平邑甜茶’PP2Cs A亚家族基因的生物信息学分析 通过DNAMAN进行多序列比对;MEME进行蛋白质基序分析;利用Cell-PLoc 2.0在线网站预测PP2C A的亚细胞定位;ExPASy ProtParam在线工具分析蛋白质理化性质;TMHMM与SignalP 4.0在线网站分析蛋白质是否存在跨膜结构和信号肽;NPS的SOPMA 工具用于蛋白质二级结构预测;SWISS-MODEL用于预测三级结构并通过NCBI Conserved Domain Search Service在线分析工具分析其蛋白质的保守功能结构域。通过MAGE 7.0软件构建系统进化树。
1.2.4 脱落酸处理下MhPP2Cs基因的表达分析 选择正常生长至3叶期的‘平邑甜茶’组培苗进行脱落酸处理。在正常的‘平邑甜茶’MS培养基中加入100 μM/L的脱落酸进行处理后取样,根据上文方法提取总RNA后反转录为cDNA。利用CFX96TM Real Time RCR System进行实时荧光定量PCR,引物序列见表1,反应体系为20 μL,包含:cDNA模板2 μL ;2×SYBR Green Pro Taq HS Premix 10 μL(湖南艾科瑞生物工程有限公司);上、下游引物(10 μmol/L)各1 μL;ddH2O 6 μL,反应程序为:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30s,40个循环。设置3次重复,使用2Δ-11Ct分析方法计算表达量差异。
2 结果与分析
2.1 ‘平邑甜茶’PP2C A亚家族基因克隆
以质量检测合格的‘平邑甜茶’叶片总RNA为模板反转录得到cDNA,克隆‘平邑甜茶’中的3个PP2C A亚家族基因的CDS序列(图1)。将这3个产物连接于pEASY-Blunt Cloning Vector后进行测序分析。
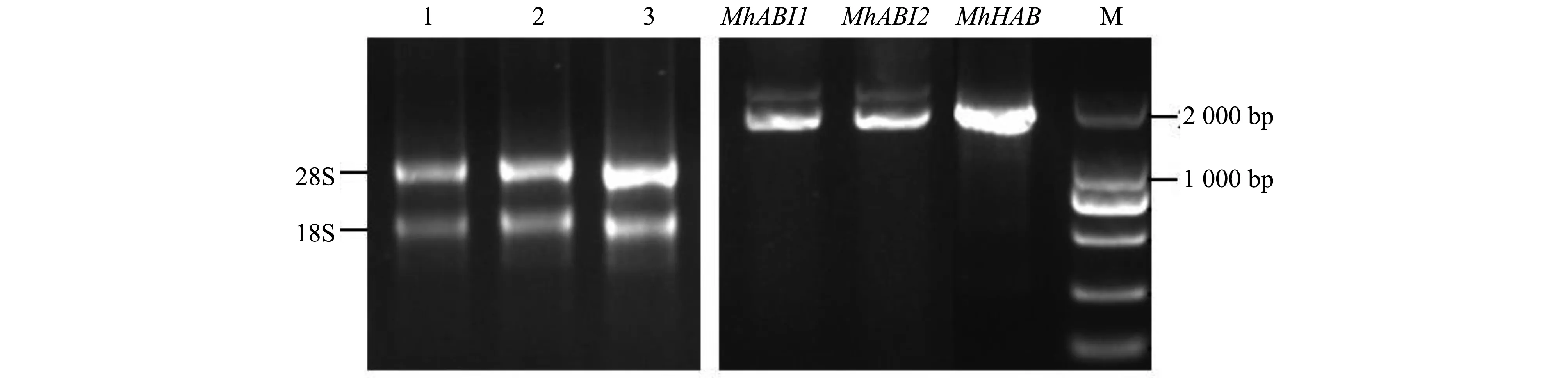
图1 ‘平邑甜茶’PP2C A亚家族基因扩增Fig.1 The cloning of group A-PP2C genes of ‘Malus hupehensis Rehd.’
2.2 亚家族基因的理化性质分析
通过在线分析网站对这3个基因编码的蛋白质进行分析,MhABI、MhABI2和MhHAB分别是编码552、552、544个氨基酸的蛋白;其预测表观分子量为58.26(MhHAB)~59.44(MhABI2) kDa;等电点为4.52(MhABI2)~5.03(MhHAB),均为偏酸性蛋白;除MhABI1不稳定系数小于40,属于稳定蛋白外,其余均为不稳定蛋白;脂肪系数为86.87(MhABI2)~93.49(MhHAB);均属于亲水性蛋白;MhABI1仅定位于细胞核,MhABI2同时定位于细胞核及叶绿体,MhHAB定位于叶绿体(表2)。
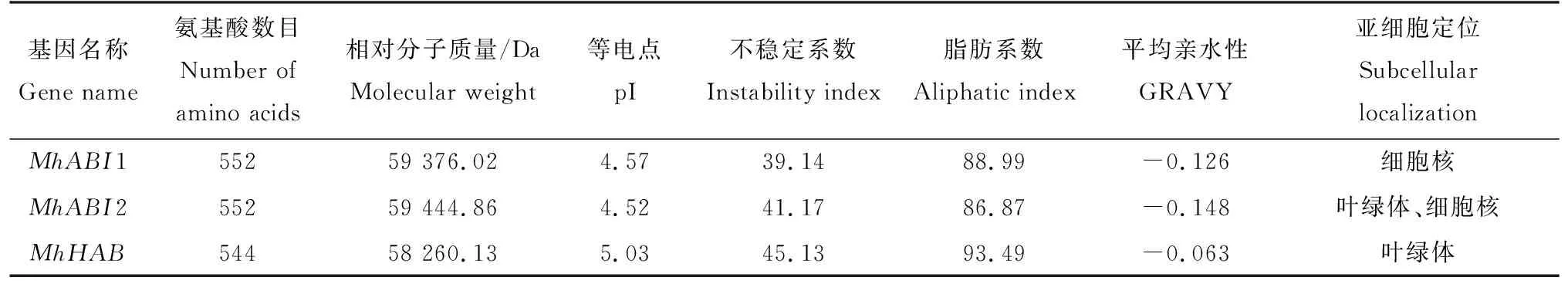
表2 ‘平邑甜茶’PP2C A亚家族蛋白生物信息学分析Tab.2 Analysis of physicochemical properties of Group A-PP2C proteins in ‘Malus hupehensis Rehd.’
利用CLUSTALW在线多序列分析工具对这3个基因编码蛋白的氨基酸序列进行同源性分析,发现其同源性为76.66%,且在第225至550个氨基酸间存在较高的相似性(图2)。进一步利用NCBI Conserved Domain Search Service在线分析工具分析这3个氨基酸的结构域,发现其高同源性区域集中在PP2C 超家族保守功能结构域处。

图2 ‘平邑甜茶’PP2C A亚家族各氨基酸序列同源性分析Fig.2 Homology analysis of the amino acid sequences of Group A-PP2Cs in‘Malus hupehensis Rehd.’
通过TMHMM与SignalP4.0在线网站预测这3个蛋白质结构发现其氨基酸序列中均不含有跨膜区及信号肽序列。再通过NPS SOPMA 工具预测各蛋白质的二级结构,发现MhABI1结构中含有α-螺旋 30.43%、β-折叠 18.48% 和无规则卷曲 46.92%;蛋白质MhABI2结构中含有α-螺旋 30.07%、β-折叠 19.75%和无规则卷曲 46.01%;蛋白质MhHAB结构中含有α-螺旋 31.62%、β-折叠 14.89%、无规则卷曲 50.18%和β-转角 3.31%。进一步利用SWISS-MODEL预测其三级结构,对3个PP2C A亚家族各蛋白质进行同源建模,发现其预测结果与二级结构预测结果基本一致,且同源建模一致度大于30%(图3)。
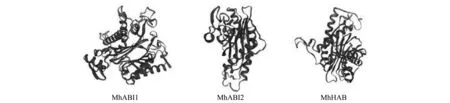
图3 ‘平邑甜茶’PP2C A亚家族蛋白三级结构Fig.3 The three-level structure prediction analysis of Group A-PP2C proteins in ‘Malus hupehensis Rehd.’
2.3 系统进化分析
由系统发育树(图4)发现MhABI1与MhABI2蛋白亲缘关系很近,包括MhHAB蛋白在内,三者与白梨的PP2C蛋白的一致性最高,其次与梅、扁桃和月季等植物中的PP2C蛋白也具有很高的一致性。同时分析保守功能域,MEME在线分析工具预测15个蛋白质的基序(motif),设置基序最大发现数为5,长度6~150个氨基酸进行分析。分析得到的5个motif,与已鉴定拟南芥PP2C A亚家族蛋白结果相似,分布于PP2C 超家族保守功能结构域处,其中motif 1~5含有41~142个氨基酸(图4)。
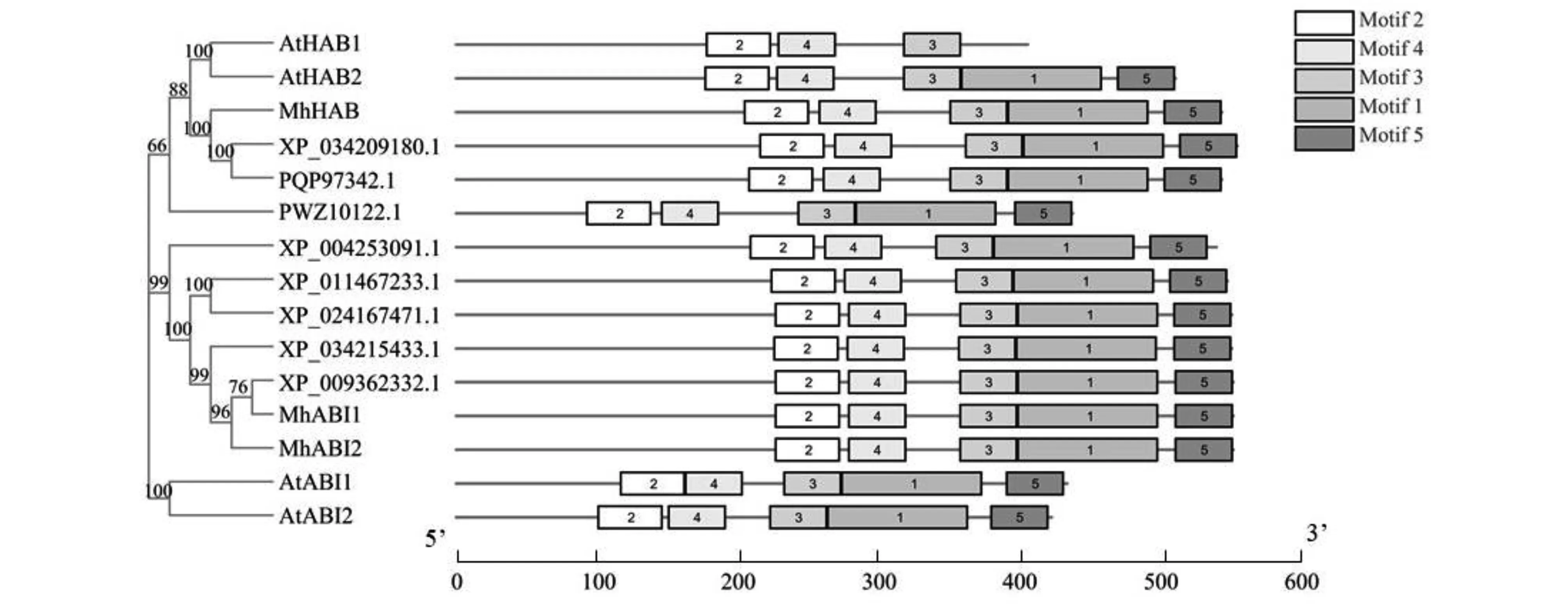
图4 ‘平邑甜茶’PP2C A亚家族蛋白的系统进化树分析Fig.4 Phylogenetic analysis of Group A-PP2C proteins in ‘Malus hupehensis Rehd.’
2.4 脱落酸处理下MhPP2Cs基因表达分析
实时荧光定量PCR结果显示,在脱落酸处理下,3个MhPP2Cs基因的表达量均显著升高,证明MhABI1、MhABI2、MhHAB在ABA信号转导过程中发挥重要作用(图5)。
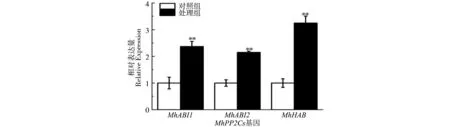
图5 ABA处理下PP2C A亚家族基因表达分析Fig.5 Expression analysis of MhPP2Cs under ABA treatment
3 讨 论
蛋白磷酸酶2C是重要的蛋白磷酸酶之一,在细胞周期、代谢、蛋白泛素化降解等过程中发挥作用[13]。在植物中,PP2Cs是ABA的共受体,参与ABA信号转导机制,对植物生长发育、胁迫响应都有着重要意义[14]。苹果属植物因其生长特性,易受干旱、冷害等非生物胁迫的影响,造成果实产量降低、品质下降,且由于成年植株高大,不易管理,进一步影响了其经济效益,对苹果进行矮化砧木嫁接,已成为目前主流的栽培方式。‘平邑甜茶’为作为目前广泛应用的苹果砧木,具有整齐度好、实生苗变异小的优良特性[15]。
本研究以苹果砧木‘平邑甜茶’为试材,通过与拟南芥的PP2C A亚家族基因进行同源性分析,从组培苗植株叶片cDNA中分离得到3个PP2C A亚家族基因。这3种蛋白均含有PP2C 超家族保守功能结构域,证明其均属于蛋白磷酸酶PP2C,推测其可能参与ABA信号转导并进一步调节植株响应干旱胁迫。但通过理化性质分析可以看出,3种PP2C A亚家族蛋白之间存在一定差异,表明其发挥的生物学功能应该不尽相同。通过分析发现3种基因编码的蛋白质均不含跨膜区及信号肽序列,而亚细胞定位结果表明MhABI1定位于细胞核、MhABI2定位于细胞核和细胞核,而MhHAB定位于叶绿体,推测它们可能通过调节细胞核基因表达或者叶绿体功能来发挥作用。同时,脱落酸处理后的基因表达量分析证实了MhABI1、MhABI2、MhHAB这三个基因在ABA信号转导途径中发挥作用。
目前,已有大量报道在拟南芥、烟草和水稻等模式植物中证明蛋白磷酸酶PP2C是ABA信号通路中的重要组成部分[16]。ABA信号转导遵循“抑制解除”的传导机制,当没有ABA存在时,PP2C会结合下游激酶OST1(SnRK2.6)并抑制其活性,阻止响应ABA信号的相关基因的表达;当ABA存在时,ABA会与其受体PYR/ PYL/ RCAR (PYLs) 结合,形成复合物,该复合物进一步与共受体PP2C结合[17],这种互作会暴露PP2C的泛素化位点,使其被降解[18],达成“抑制解除”目的,激活下游基因的表达,帮助植物应对干旱和冷害等非生物胁迫。尽管对PP2C的研究在模式植物中已取得重大进展,但类似研究在苹果属植物中仍然较少。本研究克隆并分析苹果属植物PP2C A亚家族基因,并对其编码的蛋白质进行理化性质分析、结构预测及进化树构建,通过脱落酸处理下的PP2C A亚家族基因表达量,证实在ABA信号转导中发挥重要作用,可能参与苹果砧木‘平邑甜茶’响应非生物胁迫过程,阐述了苹果属植物PP2C A亚家族成员的分子特性,有助于深入研究苹果砧木ABA信号转导途径的分子作用机制,为进一步培育抗性砧木提供理论参考。
