叶用莴苣ARF6基因克隆及高温抽薹表达分析
2022-04-13张悦婷李云峰李振峰韩莹琰刘超杰郝敬虹
张悦婷,杜 扬,李云峰,李振峰,韩莹琰,刘超杰,郝敬虹
(农业应用新技术北京市重点实验室/北京农学院 植物科学技术学院,北京102206)
叶用莴苣,可供食用的重要多叶类蔬菜之一[1]。质脆微甜,口感独特,广受人们喜爱。其适宜生长的环境温度为15~20 ℃,高于30 ℃易于抽薹,致使食用价值降低,带来减产乃至绝收的严重危害;在周年生产中生菜的高温易抽薹问题非常严峻,目前急需解决。在众多影响抽薹的因子中,植物激素如GA、IAA对抽薹开花起着重要作用[2]。其中被最早发现的生长素,参与调控植物生长发育中的各个过程[3]。
生长素的感知和信号转导依赖于多种成分的协同作用,其中生长素应答基因受三种类型的转录调节因子控制,分别是生长素应答因子(ARFs)、Aux/IAAs以及Topless(TPLs)[4]。其中生长素/吲哚-3-乙酸(Aux/IAA)蛋白起着关键作用[5]。典型的ARF蛋白,其N端有一类似于B3 DNA的结合结构域,而在C端则有类似于Aux/IAA蛋白的结构域III区和 IV区;ARF蛋白可以通过其DNA结合域与生长素的应答基因启动子区域即生长素应答元件(AuxREs)结合,包括Aux/IAAs亦可;ARF蛋白作为转录激活因子或是转录抑制因子是由它们中间区域的氨基酸组成所决定[6]。有研究表明:在拟南芥中,两个相对应的生长素响应转录因子ARF6和ARF8协同调控雄蕊和雌蕊的成熟[7]。
就目前研究结果可知,ARF6蛋白是植物体内一种非常重要的转录因子,其参与多种激素的信号转导途径,对植物的生长发育有着重要的作用及意义。然而LsARF6基因与叶用莴苣抽薹性的相关关系尚不明确,此次试验通过克隆LsARF6基因,对其进行生物信息学分析,同时运用实时荧光定量PCR技术(Quantitative Real-time PCR)分析叶用莴苣LsARF6基因在不同温度处理下各个时期的相对表达量的情况,为进一步研究ARF6蛋白在叶用莴苣抽薹过程中的作用机制奠定良好的基础。
1 材料及方法
1.1 育苗材料及处理方法
材料是农业应用新技术北京市重点实验室编号保存的易抽薹叶用莴苣品种(GB-30)。200粒种子催芽,长出胚根,将芽播种于穴盘中。待幼苗长至4叶1心时定植于营养钵内,放入植物生长箱内生长。待缓苗2~3 d,对幼苗进行处理:相对湿度均为60%,光周期均为14 h/10 h。对照组:20 ℃(日)/13 ℃(夜);高温组:33 ℃(日)/25 ℃(夜);于处理的第0、2、4、8、16、24天取植株的茎及成熟叶片,置于-80 ℃保存备用,3次重复。
1.2 总RNA提取和cDNA第一链的合成
使用总RNA快速提取试剂盒提取样品总RNA;再使用cDNA第一链合成试剂盒进行反转录,所制cDNA于-20 ℃储存,为后续LsARF6基因克隆做准备以及为实时荧光定量PCR做模板。
1.3 LsARF6基因克隆及实时荧光定量PCR
1.3.1LsARF6基因克隆 以转录组测序中的LsARF6序列为模板,使用Primer Premier 5软件设计克隆引物(LsARF6-F和LsARF6-R)和qRT-PCR引物(LsARF6-qF和LsARF6-qR),选用叶用莴苣18S作为内参基因(表1)。由生工生物工程股份有限公司提供引物。

表1 试验所用引物Tab.1 Primers used in the study
克隆LsARF6基因,经PCR反应所得产物于4 ℃保存。
上述PCR产物通过琼脂糖凝胶电泳,经紫外照射仪确定目的条带位置,使用DNA凝胶回收试剂盒回收目的条带,将回收获得的目的条带连接到pClone007 Versatile Simple Vector Kit克隆载体上,随后导入到大肠杆菌DH5α感受态细胞中培养,最后将菌液送至北京博迈德基因技术有限公司进行测序。
1.3.2 实时荧光定量PCR 以对照组和高温组各个时期的茎cDNA为模板,进行qRT-PCR。反应程序如下:95 ℃、30 s(预变性),95 ℃、10 s(变性),60 ℃、30 s(退火),39次循环。采用2-ΔΔCt相对定量分析方法进行计算LsARF6基因的相对表达量,试验数据的方差及显著性分析处理采用SPSS 20.0软件,表达量作图使用Excel 2019。
2 结果与分析
2.1 叶用莴苣ARF6基因克隆
PCR扩增得到一条长2 436 bp的特异性条带(图1),测序结果与转录组序列具有绝对一致性(100%)。
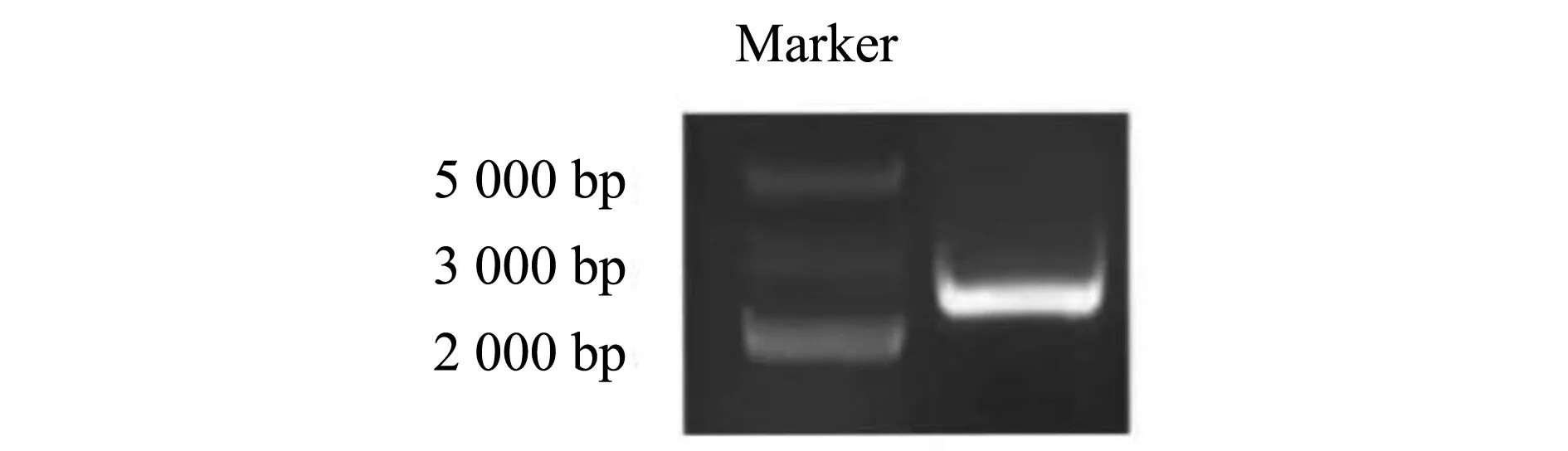
图1 扩增的LsARF6基因Fig.1 AmplifiedLsARF6 gene
运用NCBI ORF finder分析知晓,此阅读框编码了811个氨基酸。
2.2 叶用莴苣LSARF6氨基酸序列及结构域分析
2.2.1 预测LsARF6氨基酸二级结构 运用NPS的SOPMA分析,该蛋白由无规则卷曲(含56.23%)、α螺旋(含25.15%)、延伸链(含15.04%)和β转角(3.58%)组成。
2.2.2 LsARF6氨基酸序列比对 对叶用莴苣、洋蓟及青蒿的LsARF6氨基酸进行序列比对(DNAMAN 9.0软件),结果表明ARF6蛋白与其他物种(洋蓟及青蒿)的蛋白具有较高的相似性。其中LSARF6蛋白具有FSQQPPAQELI、RDLHDNEWKFRHIFRGQPKRHLLTTGWSVFVSAKRL VAGDSVLFIWNEKN、EFVIPLAK、RSVKVGWDESTAGERQPRVSLWEIEPLTTFPMYPSQFP LRLKRPWPPGLP等保守结构(图2)。

图2 LsARF6氨基酸序列比对分析Fig.2 Alignment and analysis of amino acid of LsARF6
2.2.3 LsARF6蛋白的保守结构分析 利用NCBI CD-search分析,该蛋白具有三种结构域(B3、Auxin_resp、AUX_IAA)(图3),他们分别是ARF家族蛋白与DNA结合的区域,ARF家族的保守结构域,ARF家族蛋白二聚体化的结合位点。这三个结构域为大部分的ARF家族蛋白所共有,由此推断LsARF6蛋白可能具有ARF家族蛋白的功能。
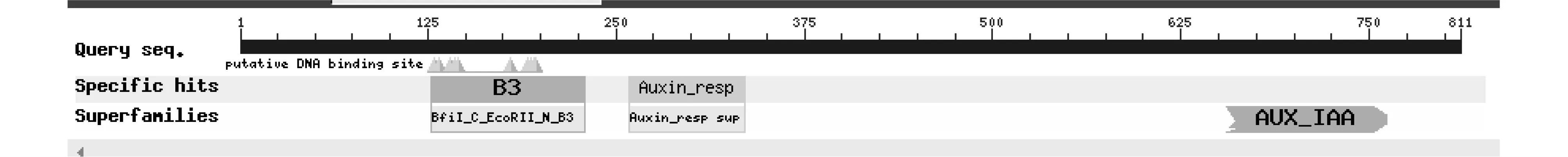
图3 LsARF6蛋白的保守结构域Fig.3 Conserved domain of LsARF6 protein
2.2.4 LsARF6氨基酸序列分析 利用ProtParam 软件得知,该蛋白分子量为89 004.40,理论等电点为5.91;其含较多的亮氨酸(含10.5%)、丝氨酸(含10.4%)、而甘氨酸(含8.3%);色氨酸(含1.8%)、酪氨酸(含1.7%)、半胱氨酸(含1.1%)含量较少,为不稳定蛋白(不稳定指数60.34);亲水性平均值为-0.357,预测为亲水性蛋白。
2.2.5 分析LsARF6蛋白信号肽 运用SignalP 5.0所得数据分析,可知其不属于分泌蛋白(不具信号肽,无跨膜结构)。
2.2.6 LsARF6蛋白质亚细胞定位 使用Psort软件进行预测,预测结果表明其亚细胞结构主要定位于细胞核。由此可知LsARF6蛋白可能在细胞核内与生长素原初响应基因的启动子上的(AuxREs)TGTCTC元件特异性结合,进而激活或抑制基因的表达,调控生长素的信号转导。
2.2.7 LsARF6保守序列分析 利用MEME软件对LsARF6的保守基因序列进行分析,发现该蛋白有位于B3、Auxin_resp、AUX_IAA保守结构域内的3个高度保守的基因序列(图4)。

图4 LsARF6的保守基因序列组成分析Fig.4 Sequence composition analysis of conserved genes LsARF6
2.2.8 LsARF6蛋白质三级结构的预测 进一步利用SWISS-MODEL推测LsARF6蛋白质三级结构,其匹配上多个ARF蛋白的模型,且含有ARF家族特有的保守结构,这一结果与NCBI CD-search分析的结果相吻合(图5)。
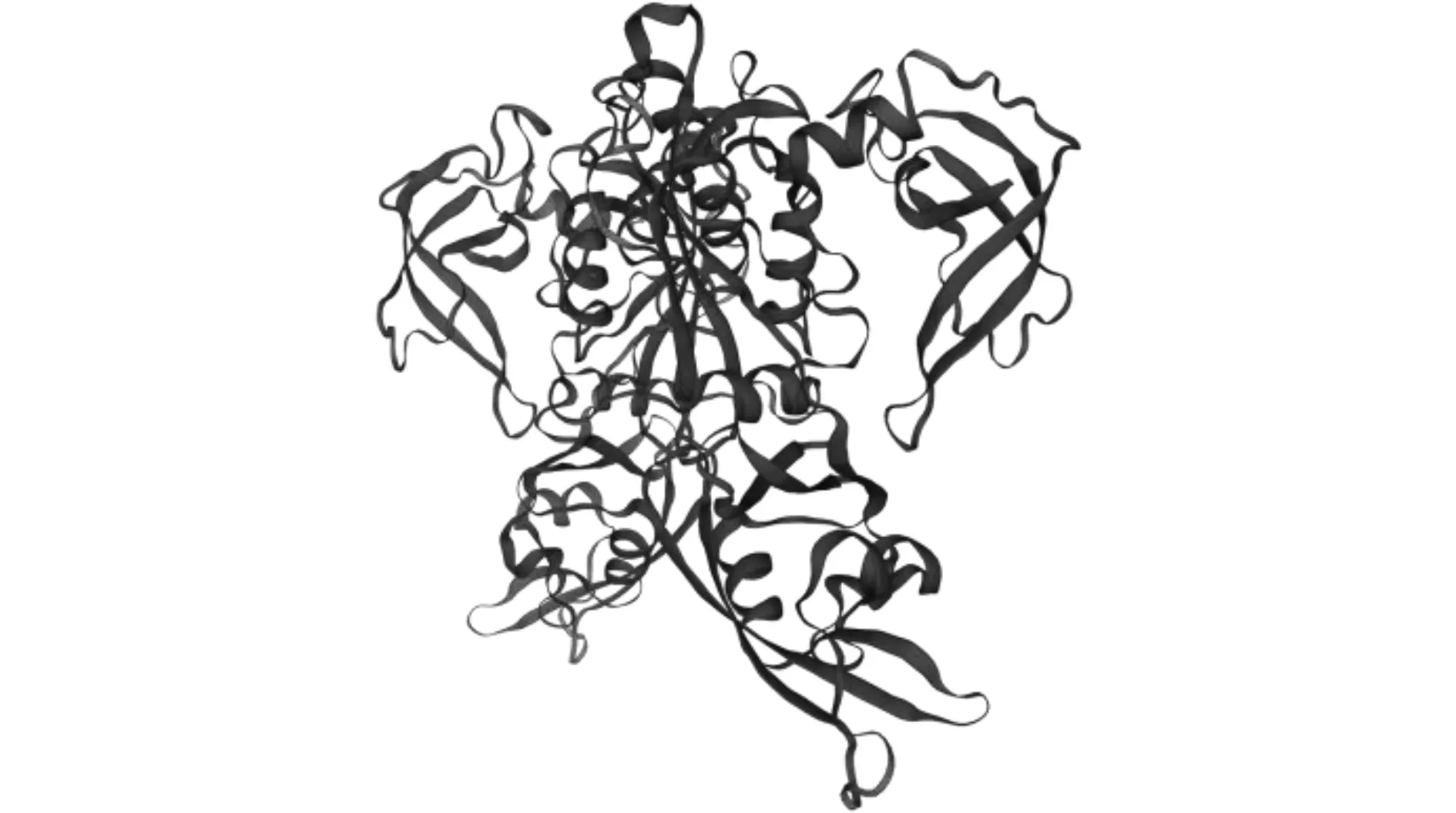
图5 LsARF6蛋白质三级结构预测Fig.5 The tertiary structure prediction of LsARF6
2.3 叶用莴苣ARF6蛋白的系统进化分析
将得到的LsARF6的氨基酸序列,在NCBI蛋白数据库中进行比对,发现它与仙人掌、月季、向日葵等10种植物中ARF6蛋白具有较高的同源性。利用软件MEGA 7.0构建系统进化树。由图6可以看出,这11条氨基酸序列明显地分为三类,刺苞菜蓟和向日葵等为第一类,两者都是菊科作物,亲缘关系最近;油菜等为第二类;芜菁为第三类。

图6 LsARF6蛋白的系统进化树Fig.6 Phylogenetic analysis of LsARF6
2.4 叶用莴苣ARF6基因高温处理后表达分析
由图7可以看出,LsARF6基因在高温组中的表达量均呈上升趋势,并且其数值均高于常温组。其中第8、16及24天的表达量差异明显,均为极显著差异。
综合上述分析,得出结论:高温促进易抽薹品种GB-30中LsARF6基因的表达,由此推测LsARF6基因可能在叶用莴苣抽薹过程中发挥功能。
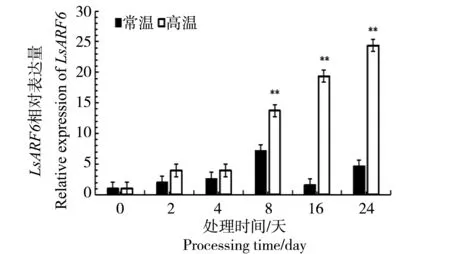
*代表显著,P<0.05;**代表极显著,P<0.01* stands for significant, P<0.05; ** stands for extremely significant, P<0.01图7 叶用莴苣GB-30在20/13 ℃、33/25 ℃下ARF6基因的相对表达量Fig.7 Relative expression analysis of ARF6 under 20/13 ℃,33/25 ℃ of lettuce GB-30
3 讨 论
在ARF家族成员的研究中,Guilfoyle等[8]对拟南芥的ARF家族进行了鉴定,共鉴定出23个AtARFs,其中 AtARF6在氨基酸序列的中间区富含谷氨酰胺(Q),丝氨酸(S)和亮氨酸(L)残基,形成了一个激活结构域(AD),能够激活下游生长素响应基因的表达。这极有可能是SlARF6基因发挥抽薹作用的一种机制。相关研究表明:在茎及花器官中SlARF6表达水平最高,在果实中也有少量表达,此结果可以证明SlARF6基因参与果实的形成和发育过程,在一定程度上可以表明ARF家族基因在番茄发育过程中具有基因功能的多样性[9]。而王飞燕等研究发现,叶片内ARF6几乎不表达,在果实中有较高的表达量;试验结果显示番茄中的SLARF6、SLARF8-1经过2,4-D处理后表达量下调,其中经GA处理后SlARF6表达量也下调;这说明内源生长素信号在植物体内的传导过程对花及幼果的生长发育极为重要,而外源激素(如赤霉素、生长素)在一定程度上能够影响某些生长素相关基因的表达,进而对果实的发育进行诱导调节[10]。这与此次试验结果有一定相似性,与高温上调SlARF6基因表达促进抽薹是相一致的。Nagpal等研究发现:整朵花生长素水平在花的成熟过程中没有发生变化,可推测生长素可能仅在特定的环境条件下或者是在花的局部器官或组织中调控花的成熟;ARF6和ARF8单突变体和倍半突变体(一个突变为纯合,另一个突变为杂合)延缓了雄蕊发育,降低了植株的繁殖能力,由此可知ARF6和ARF8基因可能影响了花的成熟时间[11]。这同时也佐证了为什么SlARF6基因是在高温条件下促进叶用莴苣抽薹。随着各物种全基因组的研究开展,ARF家族的成员在拟南芥、烟草、甘草和小麦等植物中陆续得到鉴定[12-14]。这为研究ARF转录因子家族在植物生长发育过程中的功能和作用机制,以及如何参与生长素和其他植物激素的信号转导途径等研究奠定了良好的基础。
此次试验通过对易抽薹品种中LsARF6基因在不同温度下表达量的分析发现,高温组和对照组在处理过后的第2天出现差异,第8天开始及之后处理天数差异极显著,LsARF6基因的表达水平受到高温的促进而显著上调,说明了LsARF6基因可能在叶用莴苣抽薹过程中发挥着重要的作用。后续可开展LsARF6调节叶用莴苣高温抽薹的作用机制研究。
